
种子成熟度差异对老山芹种子层积效果的影响
2022-06-02 07:53李富恒张宏发张永芳张晓雯
东北农业大学学报 2022年4期
李富恒,张宏发,张永芳,于 萍,张晓雯
(1.东北农业大学生命科学学院,哈尔滨 150030;2.山西大同大学生命科学学院,山西 大同 037009;3.北京阅微基因技术股份有限公司,北京 100044)
老山芹是东北林区主要山野菜之一,为伞形科独活属多年生宿根草本植物[1],包括短毛独活(Heracleum moellendorffiiHance)和兴安独活(Hera⁃cleum dissectumLedeb)两个种。其富含维生素C、核黄素和氨基酸等营养物质[2],具有祛风除湿、退热解毒、清洁血液、降糖降压等功效,被称为山野菜中的“绿色黄金”[3]。
老山芹种子具有较强休眠特性,采收后种胚尚未发育完全,需经历后熟过程,秋季播种后到第二年春天才发芽。生产上存在种子发芽率低、出苗不整齐、育苗技术不成熟、育苗周期过长等问题,成为老山芹规模化人工栽培限制因素。在需求量不断增加、野生资源日益枯竭情况下,开展老山芹育苗技术研究具有重要理论及实践意义,满足保护林区生态环境、发展林下经济、实现老山芹大面积人工栽培需求。
选择高成熟度种子是完善育苗技术前提和保障,有利于提高种子处理效果和发芽率。以往老山芹研究主要集中于植株及种子生长发育特性[4]、遮阴处理对幼苗生长及生理特性影响[5]、层积条件下种子后熟过程[6]、变温层积效果比较[7]、提取物活性成分及作用[8]、层积过程中种胚主要发育期间差异蛋白变化[9]等方面。本研究前期对老山芹种子成熟度差异研究结果表明,种子采收时个体间胚发育状态不一致,主要处于心形胚和鱼雷形胚时期;种子质量与种子形态和胚发育各项指标密切相关,质量越大,成熟度越高[10]。但有关老山芹不同成熟度种子对层积效果影响相关研究尚未见报道。
本研究以夏季采收短毛独活种子为材料,种子按成熟度分成高、中、低3 组,分别层积处理,测定层积过程中种子及种胚形态指标,观测并记录层积过程中种胚所处发育时期,统计种子发芽数量,比较不同成熟度种子后熟过程动态变化及种子发芽率差异。为深入研究老山芹种子休眠及萌发机制、提高种子处理效果和发芽率、保障种苗供应、发展老山芹产业提供理论依据和技术支撑。
1 材料与方法
1.1 材料
材料为短毛独活种子,于2020年8月采集于黑龙江省森林植物园(哈尔滨市)试验田。
1.2 方法
1.2.1 种子按成熟度分组
利用直径为7.5、6.0 及5.0 mm 筛子将种子分成3 组,分别作为高成熟度(平均质量12.5 mg)、中成熟度(平均质量9.2 mg)和低成熟度(平均质量6.1 mg)种子。
1.2.2 种子及基质处理
种子用75%乙醇消毒30 s,蒸馏水冲洗干净后浸种6 h,捞出备用。取干净细砂置于110 ℃烘箱中高温灭菌3 h,晾至室温后加水搅拌均匀,使其含水量达70%左右。
1.2.3 层积处理
将处理后种子与细砂按1∶3 搅拌均匀,放入长×宽×高分别为19 cm×13 cm×11 cm塑料盒中,在盖子上打孔以利于空气交换,于4 ℃低温条件下层积处理,每隔10 d清洗1次。
1.2.4 取样
分别在层积0、20、40、60、80、100、120 d时取样,测定相关指标。
1.2.5 种子形态指标测定
利用游标卡尺测定种子最大长度及宽度(mm)、胚乳最大长度及宽度(mm),分别代表种长及种宽、胚乳长及胚乳宽,重复测量10次,取平均值。
1.2.6 种胚形态指标观测
利用解剖刀切开种子并剥离种胚制成临时装片,在显微镜下运用直线式测微尺分别测定胚长、胚宽,重复测量10次,取平均值,计算胚率。
测量标准如下:
胚长(mm):直线测微尺测量种胚长度最大值;
胚宽(mm):直线测微尺测量种胚宽度最大值;
胚率计算公式:胚率(%)=胚长(mm)/胚乳长(mm)×100%;
测微尺校正:将目微尺放于目镜内,台微尺放在显微镜载物台上。移动台微尺至两尺平行,起点线重合,确定另一两尺刻度重合处,记录起点线到重合线间各尺刻度(格数)。按下式计算该放大倍数下目微尺每格所代表实际长度:
目镜测微尺每格所代表实际长度=(台微尺格数/目微尺格数)×10 μm。
1.2.7 种胚发育时期观测
利用解剖刀切开种子并剥离种胚制成临时装片,在显微镜下观测胚发育时期差异并作显微摄影。各处理每次解剖30 粒种子,重复测量3 次,取平均值。
种胚发育时期包括心形胚前、中及后期,鱼雷形胚前、中及后期,子叶期胚前、中及后期,共分为9个发育时期。
1.2.8 种子发芽数量统计和计算
在层积期间定期观察记录各处理已发芽种子数量,计算发芽率。分析各处理层积期间种子发芽率动态变化过程,比较各处理间差异。种子发芽率计算公式如下:

1.2.9 数据处理与分析
运用SPSS Statistics 19 和Excel 2019 软件分析处理数据并绘制图表。
2 结果与分析
2.1 层积过程中不同成熟度种子种胚发育进程动态变化与比较
2.1.1 种胚发育进程
老山芹种子存在形态后熟现象,即采收时种胚未发育完全,在层积过程中种胚继续发育,经心形胚-鱼雷形胚-子叶期胚阶段,逐渐完成胚后熟过程。仅完成形态后熟后种子才可能发芽。不同成熟度种子层积前种胚所处发育时期有差异,层积过程中在种胚形态、发育进程等方面也有区别。
层积前高、中、低三类成熟度种子种胚大小不同,所处发育时期也有区别。高、中两类成熟度种子胚所处发育时期多数在心形胚后期(见图1 A1 和B1),低成熟度种子胚所处发育时期多数在心形胚中期(见图1 C1)。
层积至20 d 时,高、中两类成熟度种子胚发育时期多为鱼雷形胚前期(见图1 A2 和B2),低成熟度种子胚发育时期多为心形胚后期(见图1 C2)。此时低成熟度种子胚形态与高、中两类成熟度种子未层积时种胚相似。高成熟度种子胚伸长,子叶原基基部向内凹陷。
层积至40 d 时,高、中两类成熟度种子仍保持较一致的胚发育进程,种胚发育时期主要集中在鱼雷形胚中期(见图1 A3 和B3)。低成熟度种子种胚发育时期主要集中在鱼雷形胚前期(见图1 C3)。与层积20 d相比,高成熟度种子子叶原基基部向内凹陷更明显,子叶原基向内弯曲程度更大,两原基间夹角也变大;中成熟度种子子叶原基基部向内凹陷,子叶原基尖端向内弯曲;低成熟度种子子叶原基尖端稍向内测弯曲,无子叶原基基部向内凹陷。
层积至60 d 时,高、中两类成熟度种子胚发育时期主要集中在鱼雷形胚后期(见图1 A4 和B4),低成熟度种子胚发育时期多在鱼雷形胚中期(见图1 C4)。此时高成熟度种子子叶原基及胚轴进一步变宽变长,初具子叶轮廓;中成熟度种子胚出现与高成熟度种子类似现象,但变化幅度较小;低成熟度种子胚形态发生明显变化,出现较为清晰的子叶原基基部向内凹陷。
层积至80 d 时,高、中两类成熟度种子胚发育时期已进入子叶期胚前期阶段(见图1 A5 和B5),低成熟度种子胚发育时期还处在鱼雷形胚后期阶段(见图1 C5)。与层积60 d 相比,高成熟度种子子叶原基已初步发育成子叶,子叶主脉已能观察到,胚轴变细伸长,不再是椭圆形;中成熟度种子刚刚形成子叶轮廓,未观察到主脉;低成熟度种子子叶原基与胚轴进一步伸长。
层积至100 d时,多数高、中两类成熟度种子胚发育至子叶期胚中期阶段(见图1 A6 和B6),低成熟度种子种胚发育时期仍集中在鱼雷形胚后期阶段(见图1 C6)。此时高成熟度种子子叶进一步变长、变宽,两子叶间夹角变大,子叶主脉清晰可见,网状脉序基本形成;中成熟度种子两片子叶上出现主脉;部分低成熟度种子胚发育时期仍处于鱼雷形胚后期阶段,但胚轴、子叶原基伸长生长较明显,形态与高、中两类成熟度种子层积60 d时接近。
层积至120 d 时,多数高成熟度种子胚发育至子叶期胚后期阶段(见图1 A7),完成形态后熟;中成熟度种子胚发育仍集中在子叶期胚中期阶段(见图1 B7);低成熟度种子胚发育时期集中在子叶期胚前期阶段(见图1 C7)。与层积100 d 时相比,高成熟度种子胚进一步生长发育,子叶上网状脉序已清晰可见,子叶及胚轴变宽、变长,两子叶间夹角进一步变大;中成熟度种子胚进一步生长发育,胚轴变长,子叶间夹角变大,子叶主脉更清晰;低成熟度种子胚发育进入子叶期胚前期阶段,大致轮廓与中成熟度种子层积80 d 时种胚状态类似,子叶主脉尚不清晰。
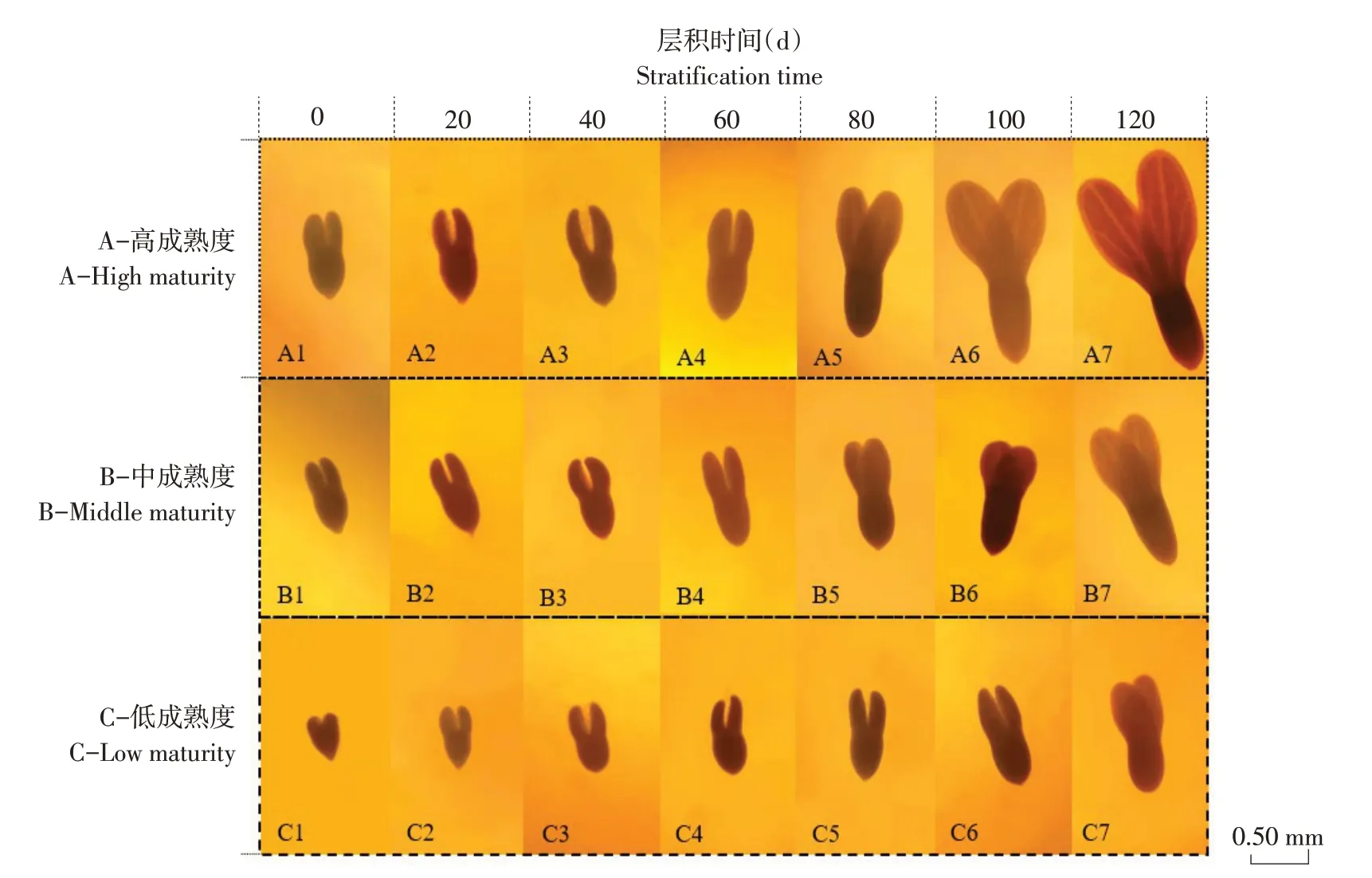
图1 层积过程中不同成熟度种子种胚发育进程Fig.1 Development process of seed embryo in different maturity during stratification
由以上分析可见,不同成熟度种子在层积过程中胚发育进程区别明显,成熟度越高,胚发育进程越快。选择高成熟度种子可缩短层积时间,有利于种子早发芽及发芽整齐,提高育苗质量。
2.1.2 种胚发育进程量化分析
老山芹种子成熟度不一致,虽将种子分成高、中和低成熟度三类,但每类种子内部依然存在这种差异。因此,有必要量化分析不同成熟度种子在层积过程中胚发育进程,定量化比较这三类种子胚发育进程差异。
由表1可看出,层积前,高、中和低三类不同成熟度种子,可观测到种胚发育时期及其所占比例有区别。其中高和中两类成熟度种子,部分种子已发育至鱼雷形胚后期,分别占6.67%和3.33%,而低成熟度种子仅有4.44%发育至鱼雷形胚中期;高和中两类成熟度种子,心形胚后期种胚占比最高,分别占供试种子总数31.11%和42.22%,低成熟度种子心形胚中期种胚占比最大,为35.56%。说明低成熟度种子种胚发育程度低于高和中两类成熟度种子;种子成熟度越高,其种胚发育程度亦高,二者呈正相关关系。
层积至20 d 时,高成熟度种子可观测到由心形胚中期至子叶期胚中期7个种胚发育时期,其中占比最高是鱼雷形胚前期种子,占供试种子总数28.89%,极显著高于除心形胚后期外其他各发育时期种子所占比例;占比最低是子叶期胚中期种子,占供试种子总数3.33%,极显著低于除心形胚中期和子叶期胚前期外其他各发育时期种子所占比例。中成熟度种子也可观测到心形胚前期至子叶期胚前期7个种胚发育时期,其中占比最高是鱼雷形胚前期,占供试种子总数35.56%,除与心形胚后期种子占比差异不显著外,极显著高于其他各发育时期种子所占比例;占比最低是子叶期胚前期种子,占供试种子总数2.22%,除与鱼雷形胚中期及后期种子所占比例差异不显著外,极显著低于其他各发育时期种子所占比例。低成熟度种子可观测到心形胚前期至鱼雷形胚后期6个种胚发育时期,比高和中成熟度处理少一个发育时期,其中占比最高是心形胚后期,占供试种子总数27.78%,极显著高于除心形胚中期及鱼雷形胚前期外其他各发育时期种子占比;占比最低是鱼雷形胚后期种子,占供试种子总数3.33%,除与鱼雷形胚中期种子占比差异不显著外,极显著低于其他各发育时期种子占比。
层积至40 d 时,高成熟度种子可观测到由心形胚中期至子叶期胚后期8个种胚发育时期,比层积20 d时多一个发育时期,2.22%种子已进入子叶期胚后期,但占比最低,除与心形胚及子叶期胚中期种子占比差异不显著外,极显著低于其他各发育时期种子占比;占比最高是鱼雷形胚中期,占供试种子总数23.33%,极显著高于除心形胚后期外其他各发育时期种子占比。中成熟度种子可观测到心形胚前期至子叶期胚中期8 个种胚发育时期,其中占比最高的处于鱼雷形胚中期,占供试种子总数26.67%,除与鱼雷形胚前期种子占比差异不显著外,极显著高于其他各发育时期种子占比;占比最低是心形胚前期种子,仅占供试种子总数2.22%,除与心形胚及子叶期胚中期种子占比差异不显著外,极显著低于其他各发育时期种子占比。低成熟度种子此时也可观测到心形胚前期至子叶期胚中期8个种胚发育时期,其中占比最高是鱼雷形胚前期,占供试种子总数26.67%,极显著高于其他各发育时期种子占比;占比最低是心形胚前期种子,仅占供试种子总数3.33%,极显著低于除子叶期胚中期之外其他各发育时期种子占比。
层积至60 d 时,高成熟度种子可观测到由心形胚后期至子叶期胚后期7个种胚发育时期,其中占比最高是鱼雷形胚后期,占供试种子总数24.44%,极显著高于其他各发育时期种子占比;占比最低是鱼雷形胚前期,占供试种子总数7.78%,极显著低于除子叶期胚前期之外其他各发育时期种子占比。中、低两类成熟度种子均可观测到心形胚中期至子叶期胚后期8 个种胚发育时期,其中中成熟度种子占比最高也是鱼雷形胚后期,占供试种子总数21.11%,极显著高于其他各发育时期种子占比;占比最低是心形胚中期,仅占供试种子总数3.33%,极显著低于其他各发育时期种子占比。低成熟度种子占比最高是鱼雷形胚中期,占供试种子总数22.22%,极显著高于除鱼雷形胚前期外其他各发育时期种子占比;占比最低是心形胚中期及子叶期胚后期种子,均占供试种子总数4.44%,极显著低于除心形胚后期外其他各发育时期种子占比。
层积至80 d 时,高成熟度种子可观测到种胚发育时期与层积60 d 时一致,仍为心形胚后期至子叶期胚后期7个种胚发育时期,但占比最高的发育时期由鱼雷形胚后期变为子叶期胚前期,占供试种子总数32.22%,极显著高于其他各发育时期种子占比;占比最低的发育时期由鱼雷形胚前期变为心形胚后期,仅占供试种子总数2.22%,极显著低于其他各发育时期种子占比。中、低两类成熟度种子也均可观测到与高成熟度一致的心形胚后期至子叶期胚后期7个种胚发育时期,但占比不同,其中占比最高的发育时期分别为子叶期胚前期和鱼雷形胚后期,分别占供试种子总数24.44%和26.67%,均极显著高于其他各发育时期种子占比;占比最低的与高成熟度种子一致,均为心形胚后期,分别占供试种子总数6.67%和4.44%,中成熟度种子除与鱼雷形胚后期、低成熟度种子除与子叶期胚后期种子占比差异不显著外,均极显著低于其他各发育时期种子占比。
层积至100 d时,高成熟度种子可观测到由鱼雷形胚前期至子叶期胚后期6个种胚发育时期,其中占比最高是子叶期胚中期,占供试种子总数38.89%,极显著高于其他各发育时期种子占比;占比最低是鱼雷形胚前期,仅占供试种子总数4.44%,极显著低于其他各发育时期种子占比。中、低成熟度种子可观测到仍为心形胚后期至子叶期胚后期7个种胚发育时期,其中中成熟度种子占比最高的发育时期为子叶期胚中期,占供试种子总数22.22%,极显著高于其他各发育时期种子占比;而低成熟度种子占比最高的发育时期仍为鱼雷形胚后期,占供试种子总数23.33%,极显著高于除鱼雷形胚中期外其他各发育时期种子占比,此时发育至子叶期胚种子占比显著上升;占比最低仍为心形胚后期,分别占供试种子总数3.33%和2.22%,均极显著低于其他各发育时期种子占比。

?
层积至120 d时,高成熟度种子可观测到种胚发育时期仍为鱼雷形胚前期至子叶期胚后期6个种胚发育时期,其中占比最高的时期由子叶期胚中期变为子叶期胚后期,占供试种子总数45.56%,极显著高于其他各发育时期种子占比;占比最低的仍是鱼雷形胚前期种子,仅占供试种子总数2.22%,极显著低于除鱼雷形胚中期外其他各发育时期种子占比。中和低两类成熟度种子也均可观测到鱼雷形胚前期至子叶期胚后期6个种胚发育时期。其中中成熟度种子占比最高的发育时期仍与上一次取样时一致,处于子叶期胚中期,占供试种子总数30.00%,较上一时期显著上升,除与子叶期胚后期种子占比差异不显著外,均极显著高于其他各发育时期种子占比;占比最低是鱼雷形胚前期,占供试种子总数6.67%,极显著低于其他各发育时期种子占比。低成熟度种子占比最高是子叶期胚前期种子,占供试种子总数24.44%,极显著高于其他各发育时期种子占比;占比最低是鱼雷形胚前期种子,占供试种子总数5.56%,极显著低于其他各发育时期种子占比。
由以上分析可知,不同成熟度种子在不同取样期均可观察到多个种胚发育时期,但总体上高成熟度种子种胚发育更快,最先进入子叶期胚后期阶段。
2.1.3 种胚后熟过程快慢比较
老山芹种子成熟度不一致,各取样时期均包括多个胚发育进程,后续比较各处理种胚后熟过程存在困难。为此,仅以高、中和低三类成熟度种子占比最大的发育时期为研究对象,用占比最大的胚发育时期代表此时种胚发育状况,比较各处理种胚后熟过程快慢。
由图2可见,层积前高、中和低三类成熟度种子占比最大的胚发育时期均在心形胚时期,其中高和中两类成熟度种子占比最大的胚发育时期为心形胚后期,而低成熟度种子是心形胚中期。
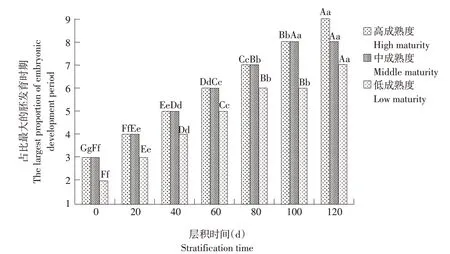
图2 层积过程中不同成熟度种子占比最大的种胚发育时期动态变化Fig.2 Dynamic change of the largest proportion of seeds with different maturity during seed embryo development during stratification process
层积至20 d 时,高和中两类成熟度种子占比最大的胚发育时期已进入鱼雷形胚阶段(鱼雷形胚前期),而低成熟度种子还停留在心形胚阶段(心形胚后期)。
层积至80 d 时,高和中两类成熟度种子占比最大的胚发育时期已进入子叶期胚阶段(子叶期胚前期),而低成熟度种子还停留在鱼雷形胚阶段(鱼雷形胚后期)。
层积至120 d时,高成熟度种子占比最大的胚发育时期已达到子叶期胚后期阶段,即多数种子已完成形态后熟,陆续进入发芽阶段;中成熟度种子占比最大的胚发育时期达到子叶期胚中期阶段;低成熟度种子占比最大的胚发育时期达到子叶期胚前期阶段。三类成熟度种子均进入了子叶期胚阶段。
计算层积过程中各处理占比最大种胚发育速率(每经历一个胚发育时期所需层积天数,d·D-1),结果见表2。结果表明,在层积80 d范围内,各成熟度处理占比最大胚发育速率相同,均为20 d·D-1;在层积100 d 时间范围内,高和中两类成熟度处理占比最大的胚发育速率保持不变,为20 d·D-1,而低成熟度种子占比最大的胚发育速率变慢,为25 d·D-1;在整个层积过程中,高成熟度处理占比最大的胚发育速率为20 d·D-1,而中和低成熟度种子为24 d·D-1。

表2 层积过程中不同成熟度种子占比最大的种胚发育速率动态变化Table 2 Dynamic change of embryo development rate was the largest proportion of seeds with different maturity during stratification
2.2 层积过程中不同成熟度种子形态指标动态变化
2.2.1 种子形态指标动态变化
由图3可知,随层积时间增加,不同成熟度种子种长、种宽、胚乳长及胚乳宽等形态指标均呈现前期增加后期变缓趋势。在层积40 d 以前,各形态指标增长幅度较大,40 d以后无变化。原因是层积前期种子吸水膨胀,各形态指标数值变大,种子充分吸水膨胀后变化停止。

图3 层积过中不同成熟度种子形态指标动态变化Fig.3 Dynamic change of seed morphology indexes of seeds with different maturity during stratification
三类成熟度种子种长、种宽、胚乳长及胚乳宽差别较大,这与各处理种子采收时种胚发育程度不同有关。成熟度越高,种胚发育程度也高,体积和质量大,各形态指标高于成熟度低处理。
2.2.2 胚形态指标动态变化
由图4可知,胚长与胚率间为正相关关系,即胚长越长,胚率越大;层积过程中不同成熟度种子胚长及胚率增长幅度及达到最大增幅时间均有差异。

图4 层积过程中不同成熟度种子胚长及胚率动态变化Fig.4 Dynamic change of long embryo rate of seed embryos with different maturity during stratification
层积前,高、中和低三类成熟度种子胚长及胚率差别明显,胚长分别为0.82、0.61和0.46 mm,胚率分别为17.47%、14.13%和12.74%。高成熟度种子胚长及胚率为中成熟度种子1.34 和1.24 倍,是低成熟度种子1.78 和1.37 倍。中成熟度种子胚长及胚率是低成熟度种子1.33 和1.11 倍。高成熟种子胚长和胚率显著高于中、低两类成熟度种子。
层积至20 d 时,高、中、低三类成熟度种子胚长分别为1.19、0.80 和0.55 mm,胚率分别为25.11%、18.22%和15.10%,较层积前胚长增幅分别为45.12%、31.15%和19.57%,胚率增幅分别为43.73%、28.95%和18.52%。高成熟度种子胚长和胚率增幅极显著高于中、低成熟度种子,中成熟度种子极显著高于低成熟度种子。
层积至40 d 时,高、中、低三类成熟度种子胚长分别为1.86、1.15 和0.68 mm,胚率分别为36.74%、24.61%和17.64%,相比上一取样时期胚长增幅分别为56.30%、43.75%和23.64%,胚率增幅分别为46.32%、35.07%和16.82%。胚长和胚率较上一层积时期增幅同样表现为高成熟种子极显著高于其他两类成熟度种子。此时高成熟度种子胚长及胚率增幅均达到最大值,而其他两类成熟度种子尚未达到最大值。
层积至60 d 时,高、中、低三类成熟度种子胚长分别为2.74、1.75 和0.98 mm,胚率分别为53.26%、36.92%和25.06%,相比上一取样时期胚长增幅分别为47.31%、52.17%和44.12%,胚率增幅分别为44.96%、50.02%和42.06%。中成熟度种子胚长和胚率增幅极显著高于高和低两类成熟度种子,而高和低两类成熟度种子之间差异不显著。此时中成熟度种子胚长及胚率增幅均已达层积期间最大值,比高成熟度种子增幅达到最大值时间晚20 d左右。
层积至80 d 时,高、中、低三类成熟度种子胚长分别为3.36、2.41 和1.43 mm,胚率分别为64.74%、50.48%和36.37%,相比上一取样时期胚长增幅分别为22.63%、37.71%和45.92%,胚率增幅分别为21.55%、36.73%和45.13%。此时低成熟度种子胚长和胚率增幅达到最大值,极显著高于高、中两类成熟度种子,比中成熟度种子增幅达到最大值时间晚20 d,比高成熟度种子晚40 d左右。
层积至100 d时,高、中、低三类成熟度种子胚长分别为3.53、2.66和1.69 mm,胚率分别为67.41%、55.27%和42.75%,相比上一取样时期胚长增幅分别为5.06%、10.37%和18.18%,胚率增幅分别为4.12%、9.49%和17.54%。此时低成熟度种子胚长和胚率增幅仍极显著高于其他两类成熟度种子。
层积至120 d 时,高、中、低三类成熟度种子胚长分别为3.68、2.88 和1.82 mm,胚率分别为69.83%、59.60%和45.94%,相比上一取样时期胚长增幅分别为4.25%、8.27%和7.69%,胚率增幅分别为3.59%、7.83%和7.46%。此时,中、低两类成熟度种子胚长和胚率增幅均显著高于高成熟度种子,而二者之间差异不显著。
2.3 层积过程中不同成熟度种子发芽率动态变化
由图5可看出,三类成熟度种子层积至50 d时陆续出现发芽现象,但不同成熟度种子在层积过程中种子发芽率均存在较大差别。

图5 层积过程不同成熟度种子萌发率动态变化Fig.5 Dynamic change of seed germination rate at different maturity during stratification
层积至50 d 时,高、中和低三类成熟度种子发芽率分别为16.96%、8.33%和3.89%。高成熟度种子发芽率约为中和低成熟度种子发芽率2.04 和4.36倍。高成熟度种子发芽率极显著高于中和低两类成熟度种子,中成熟种子发芽率极显著高于低成熟度种子,说明种子成熟度对种子发芽初期影响较大。
层积至60 d 时,高成熟度种子发芽率为23.65%,较上一层积时期增幅为39.45%。中成熟度种子发芽率为14.32%,较上一层积时期增幅为71.91%,增幅达到最大值。低成熟度种子发芽率为5.74%,较上一层积时期增幅为47.56%。高成熟度种子发芽率极显著高于中和低成熟度种子,而发芽率增幅则表现为中成熟度种子极显著高于高、低两类成熟度种子。
层积至70 d 时,高成熟度种子发芽率为30.48%,发芽率增幅降低至28.88%。中成熟度种子发芽率为18.33%,增幅降低为28.00%。低成熟度种子发芽率为9.98%,增幅达到最大值,为73.87%。此时低成熟度种子发芽率增幅极显著高于高和中两类成熟度种子,而高和中两类成熟度种子差异不显著。
层积至80 d 时,高成熟度种子发芽率为35.42%,增幅为16.21%。中成熟度种子发芽率为23.13%,增幅为26.19%。低成熟度种子发芽率为14.41%,增幅为44.39%。此时低成熟度种子发芽率增幅仍极显著高于高和中两类成熟度种子,而中成熟度种子显著高于高成熟度种子。
层积至90 d 时,高成熟度种子发芽率为38.08%,增幅降低为7.51%。中成熟度种子发芽率为26.40%,增幅为14.14%。低成熟度种子发芽率为17.35%,增幅为20.40%。低成熟度种子发芽率增幅极显著高于高成熟度种子,显著高于中成熟度种子。
层积至100 d 时,高成熟度种子发芽率为40.50%,增幅降至6.36%。中成熟度种子发芽率为29.95%,增幅为13.45%。低成熟度种子发芽率为20.58%,增幅为18.62%。低成熟度种子发芽率增幅极显著高于高成熟度种子,显著高于中成熟度种子。
层积至110 d 时,高成熟度种子发芽率为42.42%,增幅降低为4.74%。中成熟度种子发芽率为33.23%,增幅持续降低,为10.95%。低成熟度种子发芽率为24.24%,增幅变化小,为17.82%。低成熟度种子发芽率增幅仍极显著高于高成熟度种子,显著高于中成熟度种子。
层积至120 d 时,高成熟度种子发芽率为43.70%,增幅降至3.02%。中成熟度种子发芽率为34.90%,增幅为5.03%。低成熟度种子发芽率为28.41%,增幅为17.20%,此时低成熟度种子增幅极显著高于高和中两类成熟度种子,而高和中两类成熟度种子增幅差异不显著。
层积期间高、中和低三类成熟度种子发芽率为28%~44%,表现为高成熟度种子>中成熟度种子>低成熟度种子。说明选择高成熟度种子可提高发芽率、育苗效率和质量。各处理发芽率低与老山芹是野生山野菜植物有关,这是其固有生物学特性。
3 讨 论
由于人工选择和长期驯化结果,栽培植物(如各种农作物)基因型、成熟期和种子成熟度相对一致,种子休眠性弱或休眠期短,因此种子成熟度差异对生产影响较小,栽培时可不考虑这一因素。野生植物未经人工选择和驯化过程,以物种为单位种植,种内基因型多样,成熟期不一致,种子成熟度差异较大。为适应和抵御不良环境条件,野生植物种子多具有休眠特性,播种后需较长时间才发芽,存在出芽率低、出苗不齐和育苗周期长等问题,给野生植物大规模人工栽培带来困难。选择成熟度相对一致高成熟度种子可缩短种子后熟过程,有利于提高种子处理效果和发芽率,使出苗整齐一致,便于管理。
植物后熟作用主要体现在种子后熟及果实后熟两方面。后熟作用是指成熟种子或果实离开母体后,仍需经一系列生理生化变化后才完成生理成熟并具备发芽能力的生理过程[11]。种子后熟作用可分为形态后熟、生理后熟和形态生理后熟三种类型,其中老山芹种子属于形态生理后熟类型。研究发现,成熟度较高(发育期120~130 d)时,可抑制“红阳”猕猴桃果实生理后熟代谢过程、延长果实货架期,保障果实品质[12]。本研究中,高成熟度老山芹种子胚发育速率高于中、低成熟度种子,后熟过程显著加快,表明种子成熟度差异可通过影响老山芹种胚发育速率进而调控后熟过程完成。
种子萌发是指种子吸胀后一系列有序的形态发生和生理过程[13],种子萌发受多种因素协同调控,其中种子成熟度差异对其影响较为关键。种子成熟度差异原因较多,植株不同部位种子其成熟度也不同。对川泽泻(Alisma plantago- aquaticaLinn.)研究表明,主花茎第三轮及中上部位种子成熟度高于其他部位,且种子发芽率随种子成熟度增加呈双峰状变化,即开花后第20 天达到最大值(37.5%),而在第35 天达到次级高峰(31.3%)[14]。品种不同其种子成熟度也存在差异。研究发现,北柴胡(Bupleurum chinenseDC.)绿熟及黄熟种子发芽率显著高于褐熟及黑熟种子[15]。成熟度差异与种子活力有密切关系。益智(Alpinia oxyphyllaMiq.)果实开花后110 d时种子活力最大,此时种子成熟度达到最高、种子发芽率达到最大值,为最佳采收时期[16]。成熟度差异与种子硬实性有关。硬实苦参(Sophora flavescensAit.)种子其成熟度相对较高,继而种子发芽率及发芽势也表现出显著优越性[17]。成熟度差异与种子储存时间有关。研究不同成熟期穿心莲[Andrographis paniculata(Burm. f.) Nees]种子休眠和发芽规律发现,储存9 个月高成熟度(灰棕色种皮)种子达到50%发芽率可比低成熟度(灰紫色种皮)种子提早两天[18]。成熟度差异与种子采收时期有关。于大花杓兰(Cypripedium macran⁃thumSw.)授粉后60 和75 d 收集种子为早熟种子,此时种子成熟度中等,若播种在培养基上其发芽率及原球茎形成率均显著高于未成熟(45 d,成熟度低)和成熟(90 d,成熟度高)种子[19]。研究枇杷[Eriobotrya japonica(Thunb.) Lindl.]不同种种子发芽率及苗高发现,完熟期采收的品种“大房”种子其发芽率显著高于转色期种子,而品种“软条白沙”种子发芽后苗高(43.3%)显著高于转色期果实[20]。种子成熟度差异还与坐果时间有关。紫薇(Lagerstro⁃emia indicaL.)坐果后56 d 其种子(中成熟度)发芽率最高可达到93.86%,其次是坐果后63 d种子(高成熟度)发芽率为86.39%,坐果后49 d 种子(低成熟度)发芽率最低为80.71%[21]。本研究中,以种子质量作为种子成熟度分类标准,将老山芹种子分为高成熟度(平均质量12.5 mg)、中成熟度(平均质量9.2 mg)和低成熟度(平均质量6.1 mg)3组,通过对层积过程中种胚形态发育指标测定、对比发现,成熟度差异与种子质量密切相关。
研究发现,高成熟度种子其发芽率也较高。漆树[Toxicodendron vernicifluum(Stokes) F. A. Barkl.]地面收集及完全成熟的种子在发芽率、发芽势及种子活力指数等方面均显著优越于半成熟度和未成熟的种子[22]。同样,高成熟度(红果)构树[Brous⁃sonetia papyifera(Linn.)]果实种子发芽率最高,而低成熟度(微红果)果实种子发芽率较低,未成熟(青果)种子未出现发芽现象[23]。本研究中,随层积时间增加,不同成熟度种子发芽率不断升高,其中高成熟度种子发芽率(42.42%)显著高于其他两个成熟度种子,结果表明老山芹种子成熟度越高其发芽率越高。
本研究结果表明播种和层积处理时选择高成熟度种子有利于提高层积效果和出芽率,这为提高野生植物育苗技术提供新思路和新措施。今后仍需对不同成熟度老山芹种子生理后熟过程作进一步探究,包括测定糖类、可溶性蛋白等贮藏物质含量及调控物质代谢关键酶活性等,以深入研究种子成熟度差异对老山芹种子后熟过程的影响机制。
4 结 论
a.层积过程中不同成熟度老山芹种子胚发育进程有区别,成熟度越高,胚发育进程越快。高成熟度种子每经历一个胚发育时期所需层积天数为20 d,而中和低成熟度种子为24 d。
b. 高、中和低三类成熟度种子种长、种宽、胚乳长及胚乳宽等形态指标在层积40 d 前增加,40 d后基本无变化。胚长、胚率及发芽率均表现为高成熟度>中成熟度>低成熟度。
c.本试验设计范围内,高成熟度种子发芽率可达42.42%,高于中和低成熟度种子。
猜你喜欢
农业工程学报(2022年13期)2022-10-09
小资CHIC!ELEGANCE(2020年28期)2020-09-17
阅读(科学探秘)(2018年3期)2018-05-14
学苑创造·B版(2018年12期)2018-03-04
湖北农业科学(2017年6期)2017-04-26
农家顾问(2016年10期)2016-11-21
科技知识动漫(2016年5期)2016-06-04
江苏农业科学(2015年11期)2016-01-27
山西果树(2015年3期)2015-05-29
长江蔬菜·学术版(2014年7期)2015-01-21
