
幼兔和成年兔感染兔出血热病毒后血清中IL-8、IL-10、TNF-α、TGF-β和MCP-1检测分析
2022-07-06 02:09杨晓莉杨东亮毕冬琳张潇文刘方程李琼毅柏家林
西北民族大学学报(自然科学版) 2022年2期
李 菊,杨晓莉,杨东亮,毕冬琳,张潇文,刘方程,李琼毅,柏家林
(1.西北民族大学 生物医学研究中心生物工程与技术国家民委重点实验室,甘肃 兰州 730030;2.西北民族大学 生命科学与工程学院,甘肃 兰州 730030)
兔出血症(Rabbit hemorrhagic disease,RHD)俗称“兔瘟”,是兔类的一种急性、暴发性传染病,该病的病原体是兔出血症病毒(Rabbit hemorrhagic disease virus,RHDV)[1].RHDV通过口腔、鼻腔或结膜途径传播,感染兔通常死于暴发性肝衰竭(Fulminant hepatic failure,FHF),血清、肝脏和脾脏等组织中细胞因子增加[2].RHDV属于杯状病毒科(Calicivirus)兔病毒属(Lagovirus),为单股正链无囊膜RNA病毒,核酸稳定性高[3].目前,已鉴定出4个RHDV基因型[4],其中RHDV(GI.1)、RHDV2(GI.2新变异型)是较为常见的毒株[5].RHDV较RHDV2宿主相对单一,但致死性更高[6].成年兔感染RHDV后死亡率接近100%,而免疫系统不成熟的幼兔对RHDV却表现出普遍抗性[7].
研究表明,细胞因子(Cytokine,CK)在调节炎症反应和抗病毒免疫中发挥着重要作用,其异常水平可能导致炎症和免疫反应失调,会放大RHDV引起的FHF[8-9].以往研究报道,感染RHDV的成年兔血清中TGF-β、IL-10水平升高[10],随后相继发现感染成年兔血清中IL-10、TNF-β、GM-CSF[8]和IL-6、IL-8、TNF-α、IFN-γ[9]水平升高.感染幼兔血清中TNF-α、IFNα、IL-6、IL-1和IL-8水平升高,而IL-10和IFN-γ水平无变化[2].免疫幼兔感染RHDV后血清中的IL-6和IL-10水平升高[11],且感染幼兔在3天内死亡.这些数据表明,细胞因子参与了急性RHDV感染过程.
在中国,RHDV仍然是家兔死亡的首要病因.由于缺乏RHDV体外培养细胞系,RHDV对幼兔与成年兔致病差异的作用机制至今尚不清晰.因此,探索RHDV感染后幼兔和成年兔血清中细胞因子的水平差异,有助于解释该病毒感染在成年兔中的致命机制,提示血清中细胞因子在病毒性FHF中的潜在作用.本研究对感染RHDV的幼兔和成年兔血清中白细胞介素8(Interleukin 10,IL-8)、白细胞介素10(Interleukin 10,IL-10)、肿瘤坏死因子α(Tumor necrosis factor α,TNF-α)、转化生长因子β(Transforming growth factor-β,TGF-β)和单核细胞趋化蛋白1(Monocyte chemoattractant protein-1,MCP-1)水平进行检测,分析幼兔和成年兔在不同感染时间(0、12、24、36、48、60 h)血清中上述细胞因子水平的变化,以期发现RHDV感染中具有潜在作用的细胞因子,并推测其在RHDV对幼兔和成年兔致病差异中的作用机制,为阐明RHD发病机制、开发抗RHDV疫苗提供基础资料.
1 材料和方法
1.1 病毒分离
本研究病料样品采集自青海西宁已被诊断为RHDV感染死亡家兔(蔡葵蒸教授团队).在生物安全柜中,解剖携带RHDV病兔尸体,取其肝脏于-80 ℃冻融后加入等体积的PBS于4 ℃充分研磨.在-80 ℃反复冻融3次后,300 g离心10 min取上清,0.22 um滤膜过滤并加入一定量的双抗,保存于-80 ℃备用.
1.2 病毒鉴定
按照RNA提取试剂盒说明书提取1.1上清中病毒RNA,并利用试剂盒反转录成cDNA,保存于-80 ℃备用.参照NCBI中RHDV VP60基因序列(GenBank号:DQ205345.1),利用Primer 5.0设计VP60引物为F:5’-GAAAATTCATCCGCATCGGT-3’,R:5’-CTCAAAGTCATCACTTGGTCTTGTT-3’,由上海生工公司合成.以cDNA为模板,PCR进行鉴定.鉴定正确的PCR产物纯化后与pMD-18T载体连接,送上海生工公司测序.从Genbank中下载5株RHDV2毒株、6株经典RHDV毒株以及兔杯状病毒(Rabbit calicivirus,RCV)共12株病毒的VP60基因序列.采用MEGA4.1软件进行同源性分析,并构建遗传系统发育树.
1.3 实验动物
无RHDV抗体的SPF新西兰兔(甘肃省水思圆智能科技工程有限公司):2周龄幼兔6只,随机分组,每3只一组,编号标记(实验组幼兔1、2、3,对照组幼兔1、2、3);3月龄成年兔6只,随机选择并分组,每3只一组,编号标记(实验组成年兔1、2、3;对照组成年兔1、2、3).
1.4 试剂和仪器
兔白细胞介素8(IL-8)、兔白细胞介素10(IL-10)、兔肿瘤坏死因子α(TNF-α)、兔转化生长因子β(TGF-β)和兔单核细胞趋化因子1(MCP-1)ELISA试剂盒(Ruixin Biotech),生理盐水,75%医用酒精(山西可利尔生物科技有限公司),脱毛膏(广州拜高健康产业有限公司).LX-200离心机(海门其林贝尔),902-ULTS超低温冰箱(Thermo Fisher Scientifi),Centrifuge 5424R小型高速冷冻离心机(Eppendorf),酶标仪MK3(Thermo)等.
1.5 动物攻毒和样品采集
对实验组成年兔和幼兔进行RHDV攻毒试验,实验组幼兔和成年兔肌肉注射检测为RHDV核酸阳性的病兔肝脏悬液1.5 mL,对照组幼兔和成年兔同时注射等量生理盐水,注射后同等条件下分笼饲喂.在感染后0 h、12 h、24 h、36 h、48 h、60 h通过耳缘静脉采集血样2 mL,3 000 r/min离心10 min,将血清和红细胞分离,收集血清至-20 ℃箱储存备用,直至兔死亡.在感染后0 h、6 h、12 h、18 h、24 h、30 h、36 h、42 h、48 h、60 h记录兔的存活情况,出现死亡病例后采血也随之终止,60 h后将未死亡兔进行人工处死.
1.6 ELISA检测血清中细胞因子IL-8、IL-10、TNF-α、TGF-β和MCP-1表达
设置标准品孔和样品孔,所有样品以3个重复进行分析.将50 uL不同浓度标准品加入相应的标准孔,样品孔加入50 uL稀释好的待测样本.随后标准孔和样本孔中每孔加入辣根过氧化物酶(HRP)标记的检测抗体100 uL,37 ℃作用1 h,弃掉孔内液体并用 PBST 洗板5次,加入100 uL TMB显色,37 ℃避光孵育15 min,最后加入终止液终止反应.15 min内在450 nm波长处测定各孔OD值.在表格中以标准品浓度作横坐标,对应OD值作纵坐标,绘制出标准品线性回归曲线,按曲线方程计算各样本浓度值.
1.7 统计学分析

2 结果与分析
2.1 RT-PCR检测
病兔肝脏组织提取RNA进行RT-PCR检测,结果如图1所示.琼脂糖凝胶电泳得到一条474 bp左右的特异性条带,与目标基因大小一致,证实分离的病毒为RHDV阳性.

M:DNA标准DL5000;1:VP60基因扩增产物
2.2 同源性分析
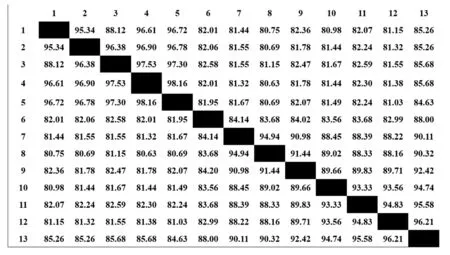
通过MEGA4.1软件分析结果显示(图2),扩增的VP60部分序列与Genbank报道的7株经典RHDV(基因型为GI.1)的VP60核苷酸序列一致性高达90.11%~96.21%,与报道的5株RHDV2型(基因型为GI.2)VP60核苷酸序列一致性为84.63%~85.68%.由此可见,本研究分离的RHDV属于经典毒株,基因型为GI.1.
1~5.兔血性病毒2型毒株;6.兔杯状病毒毒株;7~12.经典兔血性病毒毒株;13.本实验室分离的兔出血性病毒毒株
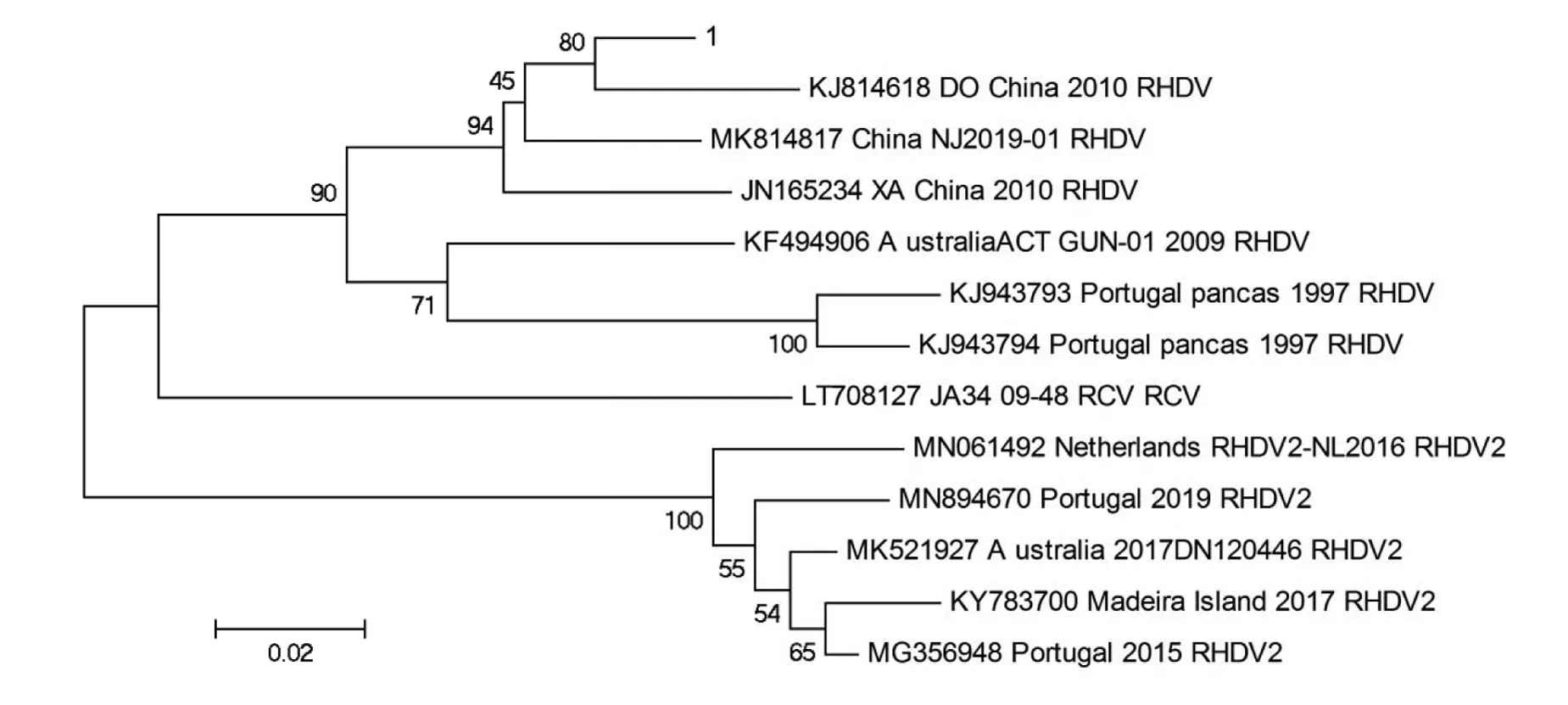
2.3 VP60基因遗传进化分析
根据进化树结果可以看出(图3),本实验室分离的毒株与经典毒株RHDV(基因型为GI.1)的亲缘关系最为接近,且与2010年中国分离出的毒株(GenBank号:KJ814618DO/China/2010)最为接近,并且他们属于同一个大的分支,与RHDV2(基因型为GI.2)的亲缘关系较远.
1.本实验室保存毒株VP60的部分核苷酸序列
2.4 RHDV感染幼兔与健康幼兔,感染成年兔与健康成年兔存活情况
RHDV感染后,幼兔与成年兔存活情况如图2-4.幼兔感染RHDV后,实验组与对照组幼兔均未出现死亡现象.成年兔感染RHDV后在36 h开始死亡,48 h时全部死亡,对照组成年兔无死亡现象.
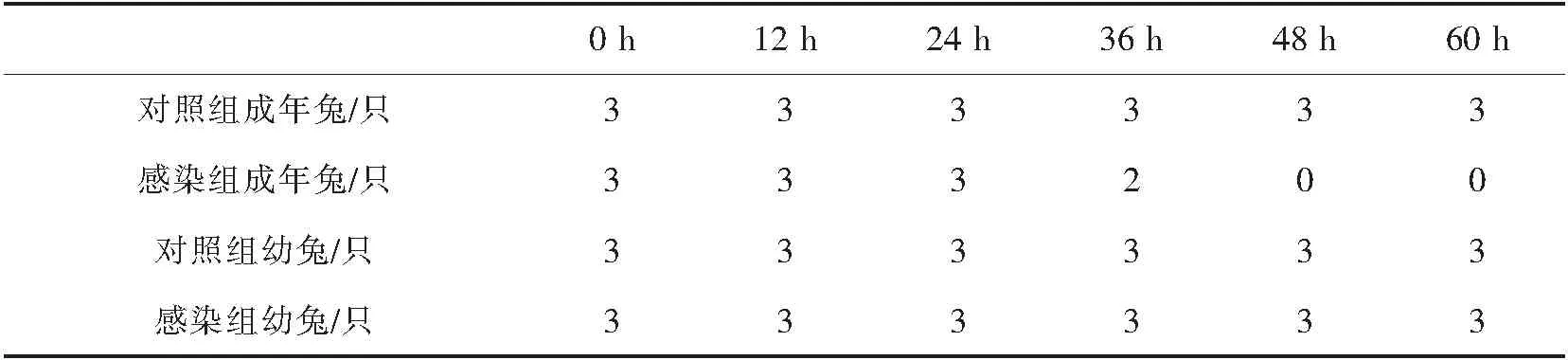
表1 幼兔和成年兔感染RHDV后的存活情况
2.5 RHDV感染成年兔血清中细胞因子水平分析
成年兔在感染RHDV后36 h开始死亡,因此检测实验组与对照组成年兔在0~36 h血清中所选细胞因子水平.结果显示,实验组成年兔血清中IL-8、IL-10和TGF-β水平在感染后明显增加,36 h均达到峰值(P<0.001)(图4A、B、C).不同的是,IL-8、IL-10在感染的0 h开始增加,TGF-β在12 h开始增加.MCP-1和TNF-α水平在感染的24 h达到峰值(P<0.001),36 h时仍明显高于对照组(P<0.01)(图4D、E);对照组成年兔血清中IL-8、IL-10、TNF-α、TGF-β和MCP-1水平均无明显变化(图2-5).结果表明,成年兔感染RHDV后血清中IL-8、IL-10、TNF-α、TGF-β和MCP-1的水平显著升高.
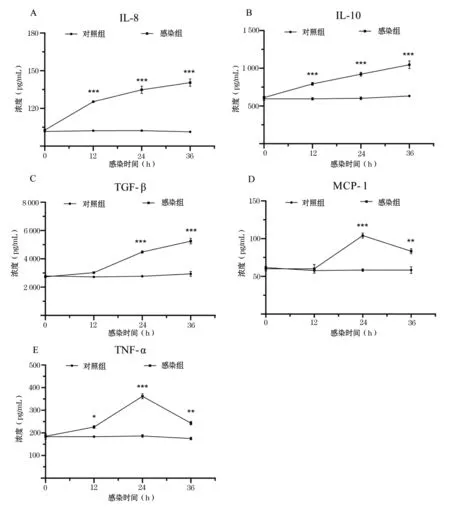
图4 成年兔感染RHDV后血清中细胞因子水平变化
2.6 RHDV感染幼兔血清中细胞因子水平分析
就幼年组而言,由于整个试验过程中未出现死亡现象,因此检测了0~ 60 h实验组与对照组幼兔血清中5种细胞因子的水平变化.结果显示,实验组幼兔血清中IL-8水平在24 h显著增加(P<0.01),36 h达到峰值(P<0.001),48 h、60 h仍显著高于对照组(P<0.05)(图4A).TNF-α和TGF-β水平变化无明显规律(图5B、C).IL-10、MCP-1水平未出现明显差异(P>0.05)(图5D、E).结果表明,幼兔感染RHDV后引起血清中IL-8水平升高.

图5 幼兔感染RHDV后血清中细胞因子水平变化
2.7 RHDV感染幼兔与成年兔血清中细胞因子水平分析
检测幼兔和成年兔感染RHDV后0~36 h血清中所选细胞因子表达量.结果显示,成年兔血清中IL-10、TGF-β的表达量在感染后显著增加(P<0.001),幼兔无明显变化(P>0.05)(图6A、B).IL-8水平在幼兔和成年兔血清中均明显增加,且成年兔中IL-8水平显著高于幼兔(P<0.05)(图6C).成年兔血清中MCP-1水平在0~36 h内均明显高于幼兔(P<0.001)(图6D).特别在幼年组血清中TNF-α水平在0~12 h内远高于成年兔(P<0.001),在12~24 h时TNF-α的含量急速增加,并在24 h高于幼兔(P<0.001),24~36 h内又以相似的速率降低,甚至在36 h低于幼兔(P<0.01)(图6E).结果表明,幼兔和成年兔感染RHDV后血清中IL-8、IL-10、TNF-α、TGF-β和MCP-1的水平存在差异.
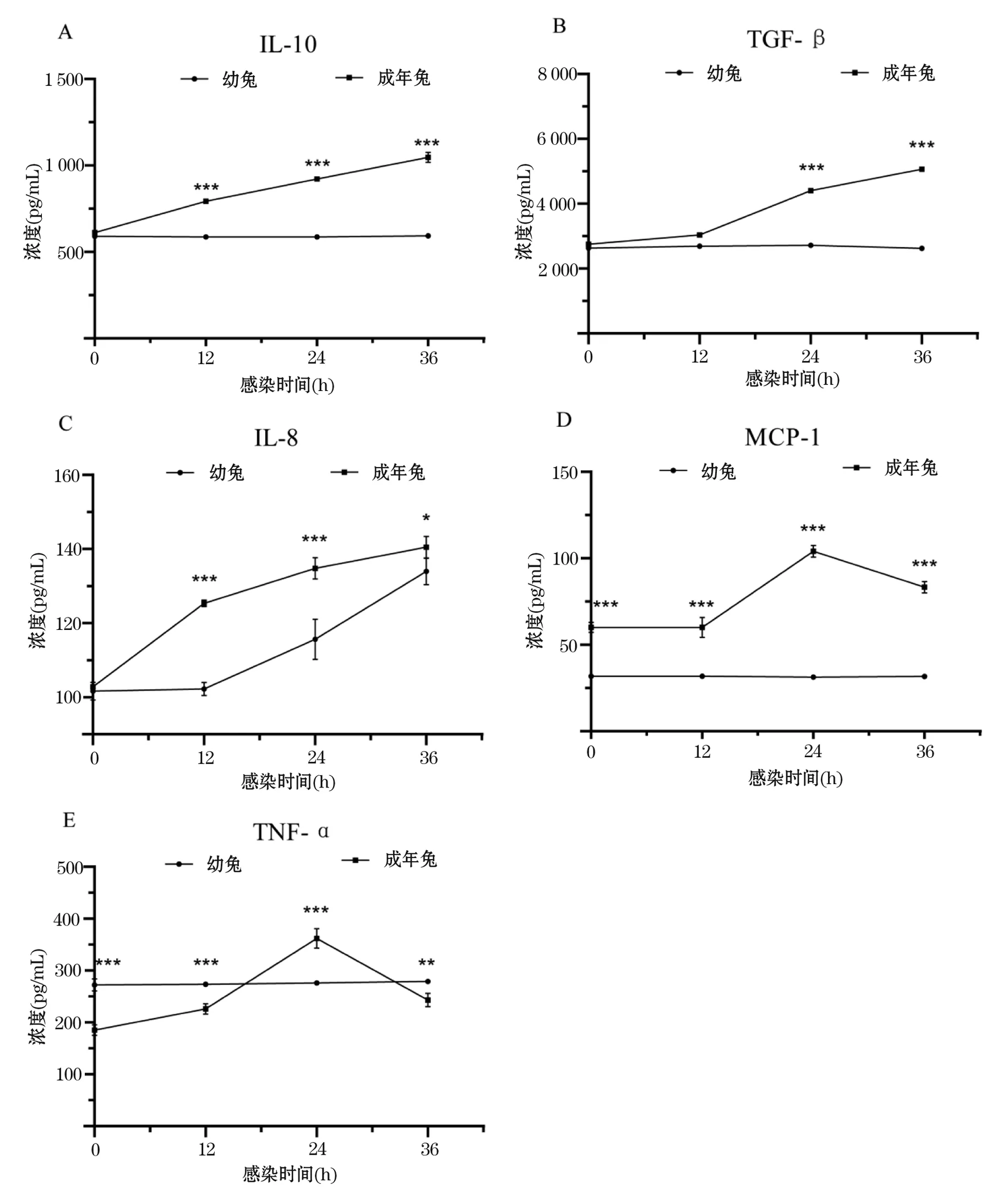
图6 幼兔与成年兔感染RHDV后血清中细胞因子水平变化
3 讨论
RHD的高发病率和高死亡率对我国的兔养殖业和野生兔类存在巨大威胁.细胞因子作为抗病毒免疫调节的重要因子,与病毒性疾病密切相关[8-9].目前,细胞因子在RHDV对幼兔和成年兔致病机制中的作用尚不明确,仅少许研究关注了细胞因子在RHDV感染的幼兔或成年兔中的作用[2,8-9],目前还没有研究指出RHDV感染后二者中水平差异的细胞因子,也并未阐明这些细胞因子在RHDV对幼兔和成年兔致病机制中的作用.
IL-8也称为C-X-C基序趋化因子配体8(C-X-C motif chemokine ligand 8,CXCL8),是一种促炎趋化因子.临床研究表明,IL-8不仅促进中性粒细胞和T淋巴细胞向组织浸润,还能激活核因子κB(Nuclear factor kappa-B,NF-κB)通路,加剧炎症反应,与慢性病毒性肝炎有关[12].
IL-10也称为细胞因子合成抑制因子(Cytokine synthesis inhibitory factor,CSIF),可抑制多种细胞因子的合成而起到抗炎作用.此外,IL-10通过抑制T细胞、自然杀伤细胞(NK)和活化的巨噬细胞功能减轻或阻止炎症反应产生,但同时抑制了天然免疫应答[13].
MCP-1又称C-C基序趋化因子配体2(C-C Motif Chemokine Ligand 2,CCL2),可调节单核细胞、记忆T淋巴细胞和NK细胞的迁移和浸润,具有将单核细胞招募到活跃炎症灶中的趋化活性,参与了抗病毒免疫反应[14].
TNF-α作为先天免疫和细胞免疫反应的中介,其水平升高能够导致过度的抗病毒免疫反应,这在低致病性流感病毒(x31,H3N2)感染的死亡小鼠[15]中可以观察到.已知在病毒感染前期TNF-α高水平有利于招募额外的靶单核细胞,因此可以猜测在RHDV感染前期TNF-α水平上升可能会导致过度免疫应答,造成机体的免疫损伤[15].
TGF-β是体内最强的免疫功能抑制细胞因子,其早期的高水平抑制了NK细胞功能和早期病毒控制,加重了新冠病毒(Coronavirus disease 2019 ,COVID-19)感染者疾病进程[16].由此我们猜想TGF-β对RHDV感染兔(5E)可能具有相似的作用,即TGF-β的高水平可能抑制了机体对RHDV的控制与清除,是引起成年兔死亡的潜在因素.
细胞因子作为炎症反应和免疫反应的主要蛋白,在短期刺激下可诱发适宜的炎症反应和免疫反应,而在病毒感染刺激下诱导产生的高水平细胞因子长期作用时可引发过度的免疫应答,造成机体多器官功能障碍和感染动物的死亡.许多研究显示,细胞因子的异常水平与严重病毒性疾病有关,包括急性肝炎、流感和新冠肺炎等[9,12,15-16].本研究表明,IL-8、IL-10、TNF-α、TGF-β和MCP-1在RHD发病机制中具有重要作用,幼兔与成年兔对RHDV的不同抗性可能与其水平差异有关.本研究所获得的结果可能会证明细胞因子在病毒性FHF中的潜在作用.
猜你喜欢
中国现代医生(2022年21期)2022-08-22
罕少疾病杂志(2022年6期)2022-06-07
科学大观园(2022年2期)2022-01-23
文萃报·周二版(2021年47期)2021-12-14
当代体育(2021年7期)2021-09-10
环球时报(2021-07-12)2021-07-12
现代养生·下半月(2018年7期)2018-09-12
师道·教研(2017年4期)2017-04-22
祝您健康(1990年6期)1990-12-30
