
刺槐Kuntiz型胰蛋白酶抑制剂基因(RpKTI)的克隆及表达分析
2022-07-09 04:00张晓静董启迪王霞张晓娜周建
山东农业科学 2022年6期
张晓静,董启迪,王霞,张晓娜,周建,3
(1.河南科技学院园艺园林学院,河南 新乡 453003;2.长垣市林业局,河南 长垣 453400;3.河南省园艺植物资源利用与种质创新工程研究中心,河南 新乡 453003)
胰蛋白酶抑制剂(trypsin inhibitor,TI),即抗胰蛋白酶因子(antitrypsin),属于蛋白酶抑制剂四大分类中的丝氨酸蛋白酶抑制剂[1]。TI在动植物及微生物中均有发现,最早提取于牛胰腺[2],后来又在大豆中分离出Kunitz型胰蛋白酶抑制剂(Kunitz trypsin inhibitor,KTI)[3]。对植物TI的研究主要集中于豆科,但在茄科、禾本科以及十字花科中也有发现[4,5]。
目前,对KTI的研究较为深入[6-8],分子量在21 kDa左右,属于丝氨酸蛋白酶抑制剂,在植物抗性研究中的作用极其重要,主要涉及对生物逆境与非生物逆境的响应,如盐胁迫、抗虫能力等[9]。高越峰等[10]得到的转基因(大豆Kunitz型胰蛋白酶抑制剂基因,SKTI)烟草植株具有明显的抗棉铃虫能力。谢可方等[11]发现大豆Kunitz型胰蛋白酶抑制剂基因(SBTi-A2)对棉铃虫幼虫生长有一定的抑制作用;大豆根系的胰蛋白酶抑制剂(STI)能阻碍大豆胞囊线虫的发育[12]。Srinivasan等[13]克隆了烟草胰蛋白酶抑制剂基因(NtPI),发现转基因烟草相比野生型烟草对NaCl的耐受性更强;在pH值为4~8之间可正常生长;对斜纹夜蛾和棉铃虫有一定的抗性,表明烟草NtPI基因具有多重抗胁迫性。向缅[14]将克隆得到的决明Kunitz型胰蛋白酶抑制剂基因2(COTI2)转入拟南芥并进行盐胁迫及干旱胁迫处理,发现转基因拟南芥抗盐及抗旱能力均优于野生型。
TI的作用广泛,通过转基因技术可以极大地提高转基因植株对逆境胁迫的抗性响应。但目前在刺槐中的研究并不多,尤其是有关非生物逆境胁迫响应的相关研究报道较少。
刺槐为豆科刺槐属落叶乔木,是重要的生态保护木本植物,有很强的抗旱、耐瘠薄、耐盐碱、耐铅镉能力,对土壤改良有重要意义[15,16]。由于其根蘖及枝条萌发能力强,因此多与其他树种组成混交林用于保土固沙,成为防护造林、生态修复必不可少的物种之一。我国土壤质量差,污染问题越来越严峻,尤其是重金属污染,严重危害人类健康[17]。因此,对重金属污染的修复极其重要。本试验在铅镉胁迫下提取并克隆得到刺槐Kunitz型胰蛋白酶抑制剂基因(RpKTI),并对其进行生物信息学分析及表达分析,为后期RpKTI基因的功能验证、应用及其种质资源创新等方面的研究奠定理论基础。
1 材料与方法
1.1 试验材料
试验所用植物材料为刺槐(Robinia pseudoacacia L.);大肠杆菌(Escherichia coli))感受态细胞DH5α;载体为pMD18-T(Takara)。
1.2 试验方法
1.2.1 总RNA的提取 采用改良CTAB法[18]提取刺槐叶片的总RNA,利用NanoDrop 2000C分光光度计检测浓度,并采用PrimeScriptTMⅡ1st Strand cDNA Synthesis Kit试剂盒将其反转录成cDNA。
1.2.2 刺槐KTI基因的克隆 刺槐KTI基因的克隆采用SMARTer RACE 5′/3′Kit(Takara,Code No.634858)试剂盒进行。核心目的片段由本课题组前期对刺槐的转录组测序数据分析获得。根据核心片段分别设计内外引物(表1)进行3′端和5′端的扩增,扩增体系如表2。按照试剂盒说明书合成RACE Ready cDNA作为第一轮扩增模板,扩增引物为外引物,第二轮扩增体系的模板为稀释后的第一轮扩增体系产物,扩增引物为内引物。扩增程序均为95℃1 min;95℃30 s,55℃30 s,68℃30 s,35个循环;68℃3 min。将获得的PCR产物采用1%琼脂糖凝胶电泳进行检测,目的条带切胶回收纯化后连接到载体pMD18-T,并转化至大肠杆菌感受态细胞DH5α,挑取单菌落摇菌后进行PCR检测,选择阳性克隆菌落送至上海生工生物工程股份有限公司测序。将3′端和5′端扩增序列片段通过DNAMAN 6.0进行拼接,设计CDS区扩增引物,并克隆全长,扩增体系如表3,扩增程序同上。
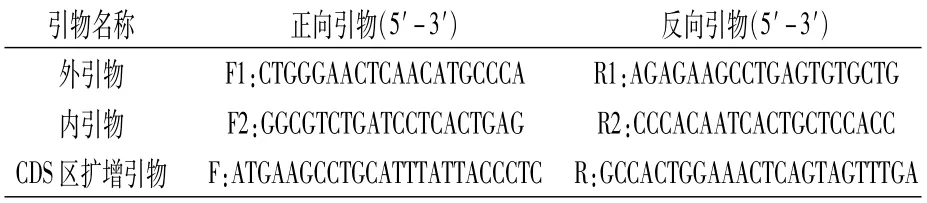
表1 刺槐KTI基因克隆的引物序列

表2 3′/5′-RACE PCR扩增体系

表3 刺槐KTI基因全长PCR扩增体系
1.2.3 刺槐KTI基因的生物信息学分析 利用ORF finder分析RpKTI基因的开放阅读框;Prot-Param及ProtScale在线程序对其基本理化性质及亲/疏水性进行预测;通过TMHMM 2.0(Tiedmixture Hidden Markov Models)分析RpKTI基因编码蛋白的跨膜结构域;SignalP-4.1分析其信号肽位点;在线工具NetPhos 3.1及NetNGlyc 1.0分析RpKTI基因编码蛋白的磷酸化及糖基化位点;SOPMA在线分析预测该蛋白的二级结构;建模软件SWISS-MODEL对RpKTI基因编码蛋白的氨基酸序列同源建模得到蛋白质的三维结构模型;通过NCBI Conserved Domain Search分析RpKTI基因的保守结构域;利用DNAMAN 6.0及MEGAX软件分析刺槐及其他豆科植物的KTI基因编码蛋白的同源性,从而构建系统进化树(表4)。

表4 生物信息学分析工具及其网址
1.2.4 刺槐KTI基因的组织表达分析 将刺槐幼苗于铅(2 000 mg/kg)、镉(80 mg/kg)胁迫下分别处理3 d及45 d,以不作处理的植株为空白对照(0 d)。取刺槐幼苗叶片迅速放入-80℃液氮中冻存,并进行RNA提取及反转录试验。
根据KTI基因的全长序列,以刺槐Actin基因为内参,使用NCBI中的Primer designing tool设计实时荧光定量PCR(qRT-PCR)引物(F:5′-TTTCCCAAGAGAATGGGTGGG-3′,R:5′-GGTGGAGCAGTGATTGTGGG-3′),扩增大小为135 bp。扩增体系(10μL):TB Green Premix Ex TaqⅡ(Tli RNaseH Plus)(2×)5μL,引物各0.5μL,cDNA 0.5μL,ddH2O 3.5μL。扩增程序为:95℃30 s;95℃5 s,50℃30 s,共40个循环;95℃5 s,65℃5 s,95℃5 s。采用2-ΔΔCt算法计算KTI基因的表达水平。
2 结果与分析
2.1 刺槐KTI基因全长的克隆及ORF预测
通过对刺槐KTI基因3′端和5′端进行PCR扩增,得到3′端序列长度约500 bp,5′端序列长度约为400 bp(图1a)。测序结果显示KTI基因长度为633 bp(图1b),经过拼接得到刺槐KTI基因序列全长为745 bp(图2),将此基因命名为RpKTI。通过开放阅读框ORF预测(图3),RpKTI基因共编码210个氨基酸,起始密码子为ATG,该序列的最后一个密码子为GTT,终止密码子为TGA。编码区两侧为11 bp(735~745 bp)的5′端非翻译区和101 bp(1~101 bp)的3′段非翻译区。

图1 刺槐KTI基因的PCR扩增

图2 刺槐KTI基因拼接序列全长

图3 刺槐KTI基因的开放阅读框(a)及氨基酸序列(b)分析
2.2 RpKTI基因的基本理化性质
RpKTI基因共编码210个氨基酸,其中含有不带电荷的极性亲水氨基酸(Y、S、T、C、N、Q)50个,占比23.8%;非极性疏水氨基酸(A、G、V、L、I、P、F、M、W)119个,占比56.7%;酸性氨基酸(D、E)25个,占比11.9%;碱性氨基酸(K、H、R)16个,占比7.6%。甘氨酸(Gly)数量最多,占全部氨基酸的11.4%(表5)。理论等电点PI为4.51,相对分子质量为22 763.87 Da,原子总数3 195,分子式为C1039H1586N258O307S5。不稳定系数(instability index)为38.81(<40),脂肪系数为89.05,总平均亲水性(GRAVY)值为0.072。因此,预测此蛋白是一个稳定的酸性疏水蛋白。
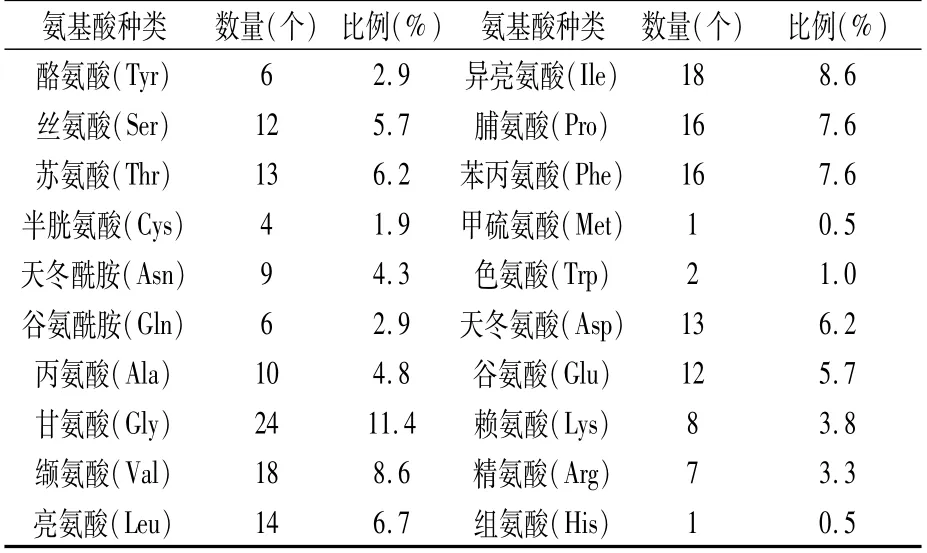
表5 RpKTI基因编码蛋白的氨基酸组成
2.3 RpKTI基因编码蛋白的亲/疏水性预测
通过在线ProtScale工具对RpKTI基因编码蛋白的亲/疏水性进行预测。由图4可知,在氨基酸序列的第12位亮氨酸(Leu)具有最大值2.8,疏水性最强,肽链的第182位谷氨酸(Glu)具有最小值-3.2,亲水性最强。整体看疏水氨基酸与亲水氨基酸分布较为均匀。结合ProtParam预测结果,此蛋白为疏水蛋白。在第13位氨基酸位置出现一个较强的疏水区域(score>1.5),推测此位置可能存在一个跨膜结构。
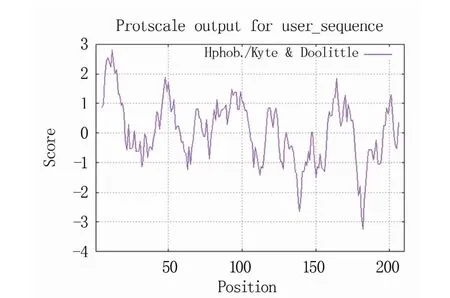
图4 RpKTI基因编码蛋白的亲/疏水性
2.4 RpKTI基因编码蛋白的跨膜结构域预测及信号肽分析
通过TMHMM对RpKTI基因编码蛋白的跨膜结构域进行预测,结果(图5a)表明,RpKTI基因编码蛋白可能存在1个跨膜结构域,此跨膜结构位于氨基酸序列N-端的第3~23位氨基酸,由膜内到膜外,且可能含有信号肽。经过SignalP-4.1分析发现,该蛋白中存在信号肽,信号肽切割位点位于第25位氨基酸(即C值最高点,图5b)。1~24位氨基酸为此蛋白的信号肽区域,信号肽序列为MKPAFITLSFLLFAFITNLQLTFS。
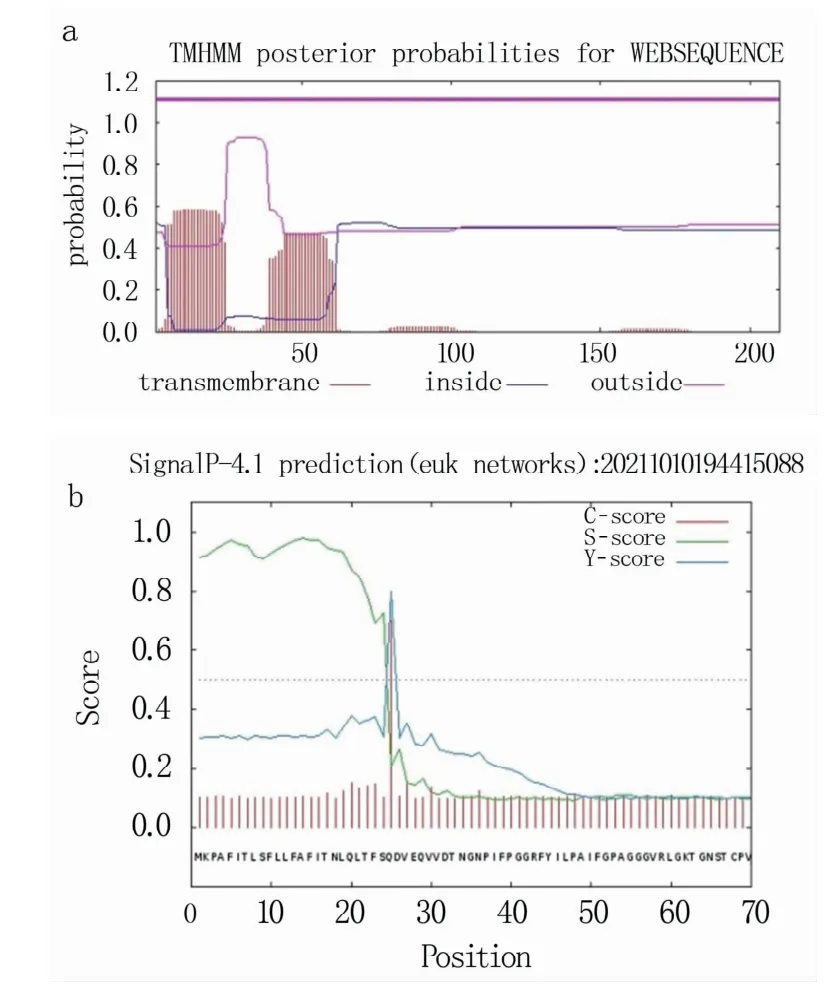
图5 RpKTI基因编码蛋白的跨膜结构域(a)和信号肽位点(b)
2.5 RpKTI基因编码蛋白的磷酸化位点及糖基化位点
蛋白质翻译后的磷酸化及糖基化分析结果表明RpKTI基因编码蛋白有11个磷酸化位点(图6a),其中有7个丝氨酸(Ser)位点、3个苏氨酸(Thr)位点和1个酪氨酸(Tyr)位点。N-糖基化位点位于第65位氨基酸天冬酰胺(Asn,图6b)。

图6 RpKTI基因编码蛋白的磷酸化位点(a)和糖基化位点(b)
2.6 RpKTI基因编码蛋白的二级结构预测
SPOMA分析结果(图7)表明,RpKTI基因编码蛋白的二级结构由31个氨基酸构成α-螺旋(alpha helix;14.76%),32个氨基酸构成β-转角(beta turn;15.24%),68个氨基酸构成延伸链(extended strand;32.38%),以及79个氨基酸构成无规则卷曲(random coil;37.62%)。在此蛋白的二级结构中,以延伸链和无规则卷曲为主。
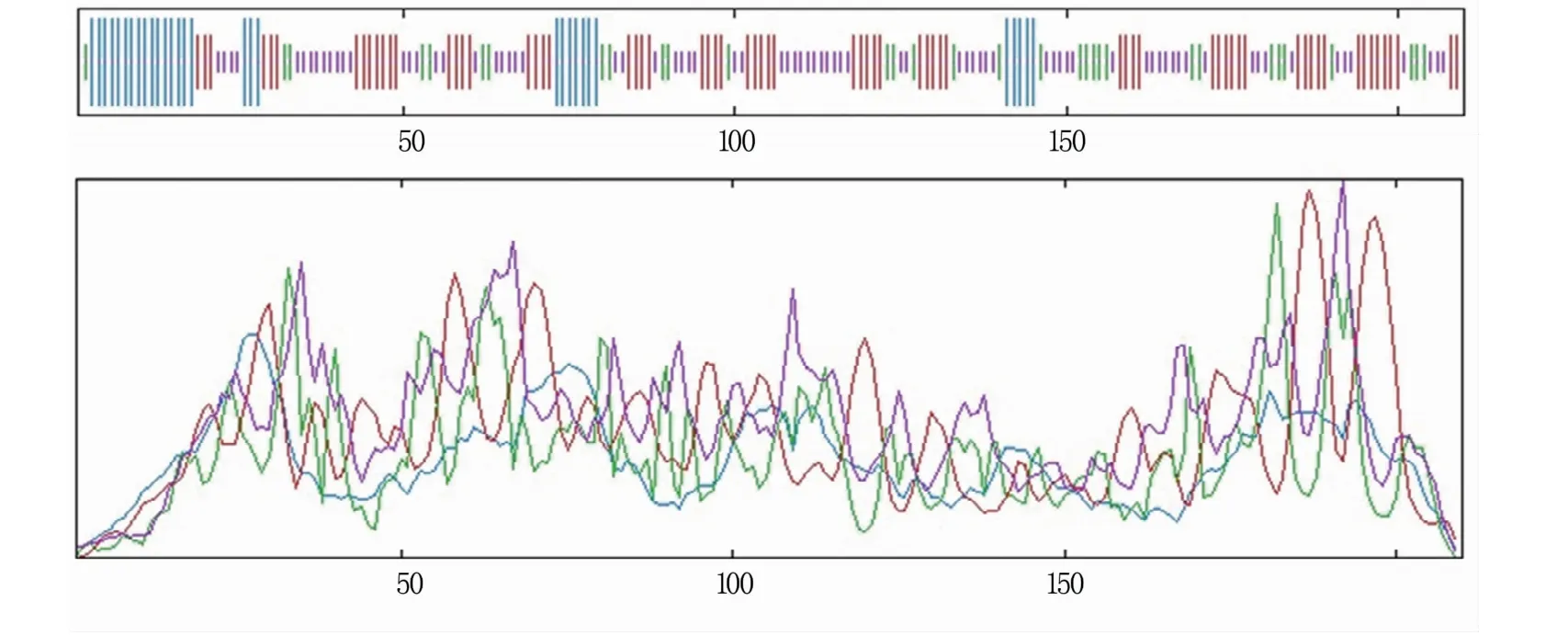
图7 RpKTI基因编码蛋白的二级结构
2.7 RpKTI基因编码蛋白的三维结构同源建模
建模结果(图8)表明,第27~203位氨基酸参与建模,以5xoz.1.A为模板,两者一致性可达66.85%,GMQE值为0.77。从图8a可知,此蛋白主要是由无规则卷曲及延伸链构成,与二级结构预测结果一致。拉式图(Ramachandran Plots)的Ramachandran Favoured值为92.57%,表明预测构象合理可用(图8b)。
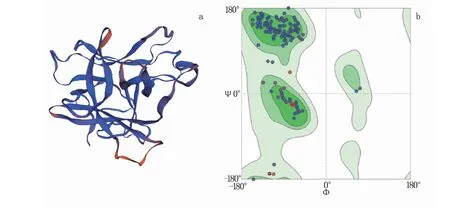
图8 RpKTI基因编码蛋白的三维结构(a)和拉式图(b)
2.8 RpKTI基因编码蛋白的系统进化树
通过对RpKTI基因编码蛋白进行Blastp比对(图9),此蛋白氨基酸序列中具有STI[soybean trypsin inhibitor(Kunitz)family of protease inhibitors]结构域,属于STI超级家族。选取相思子(Abrus precatorius)、黧豆(Mucuna pruriens)、蔓花生(Arachis duranensis)、花生(Arachis hypogaea)、木豆(Cajanus cajan)、豌豆(Pisum sativum)6种与刺槐同源性较高的豆科植物KTI基因氨基酸序列,采用DNAMAN软件进行多序列比对,同源性为72.64%,由图10可知,KTI基因在豆科植物中相对保守。
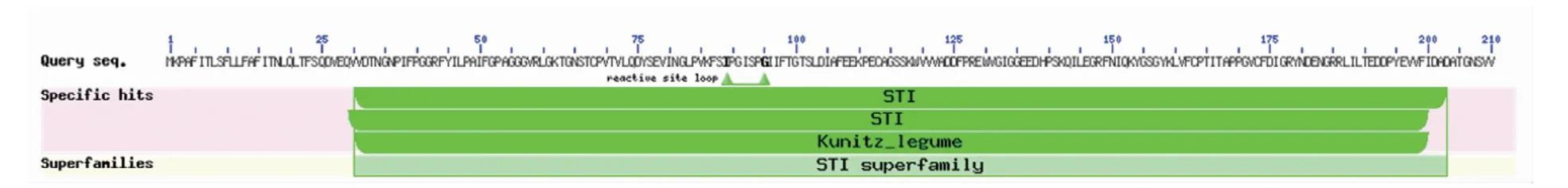
图9 RpKTI基因编码蛋白的保守结构域

图10 RpKTI基因的氨基酸序列与其他植物KTI的同源比对
通过MEGA-X软件的邻位归并法(Neighborjoining,NJ)对豆科植物Kuntiz型胰蛋白酶抑制剂基因(KTI)构建系统进化树(图11),发现刺槐与蒺藜苜蓿位于同一分支,亲缘关系较近,而与鹰嘴豆(Cicer arietinum)、蔓花生(Arachis duranensis)及花生(Arachis hypogaea)的亲缘关系较远。
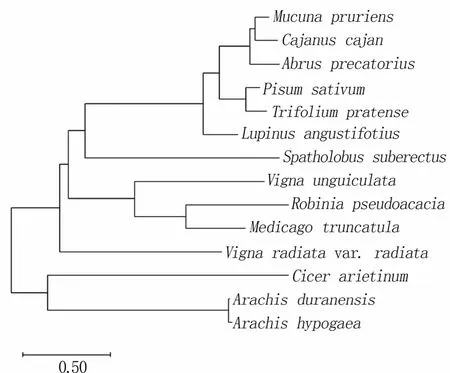
图11 豆科植物KTI基因的系统进化树
2.9 RpKTI基因的相对表达量
如图12所示,在铅镉混合胁迫时,随处理时间的增加,RpKTI基因的表达量先增加后下降,在混合胁迫处理3 d时基因的相对表达量达到最高,约为对照(0 d)的3倍,而在混合胁迫45 d时又明显下降。
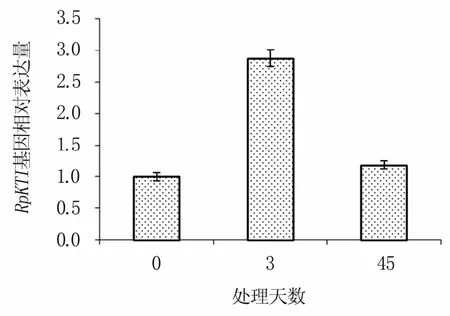
图12 铅镉混合胁迫下RpKTI基因的相对表达量
3 讨论与结论
Kunitz型胰蛋白酶抑制剂广泛存在,在豆科植物中的含量最多,目前已在多种植物中进行了分离及深入研究,如大豆、杨树、烟草等[19-21]。研究表明,KTI基因参与多种生物胁迫及非生物胁迫的响应,如抗虫、耐盐碱等。
实验室前期的转录组分析显示,RpKTI基因的表达量在刺槐应答铅镉胁迫过程中出现明显的上调,表明RpKTI基因可能参与了刺槐对铅镉的胁迫应答过程。本研究成功从刺槐中克隆得到RpKTI基因全长序列,其开放阅读框ORF长度为633 bp,共编码210个氨基酸,相对分子量为22 763.87 Da,与决明(26.68548 kDa)、鹰嘴豆(25.7 kDa)、黑豆(23.9 kDa)、蒙古沙冬青(21.5 kDa)等豆科植物的KTI基因分子量大小相近[22-25],属于STI家族。RpKTI基因编码蛋白的信号肽序列位于整个氨基酸序列的第1~24位;具有跨膜结构和信号肽,属于分泌蛋白;磷酸化主要集中在Ser、Thr、Tyr氨基酸位点。糖基化根据被修饰的类型可分为4类,以N-糖基化类型最多见,一般在Asn-X-Ser/Thr模式序列上出现[26]。RpKTI基因编码蛋白含有磷酸化位点11个,其中Ser位点最多,可能参与蛋白质的磷酸化调控;N-糖基化位点1个,位于第65位Asn氨基酸位点;RpKTI基因编码蛋白的二、三级结构主要以延伸链(32.38%)和无规则卷曲(37.62%)为主,而无规则卷曲影响着蛋白质的空间构象及其功能[27]。RpKTI基因的氨基酸序列与其他豆科植物的同源性为72.64%,可能KTI基因在豆科植物中是一个较为保守的基因。系统进化树发现其与蒺藜苜蓿的亲缘关系更近。
为了进一步了解RpKTI基因的表达水平,通过qRT-PCR分析了RpKTI基因在铅镉胁迫下的表达量,在铅镉混合处理3 d时,叶中的表达量最高,随着胁迫时间的延长,对胁迫的响应呈下降趋势,可能在此状态下,植株已经对铅镉的吸收达到饱和状态,并对铅镉胁迫产生耐性[28],其具体功能表达还需进一步试验分析。
通过对RpKTI基因的克隆、生物信息学分析及表达分析,为后期RpKTI基因的功能验证、解析其与刺槐铅镉耐受性及其作用机理提供了理论依据。
猜你喜欢
分子催化(2022年1期)2022-11-02
中国农业科学(2022年16期)2022-09-19
中国典型病例大全(2022年11期)2022-05-13
健康体检与管理(2022年4期)2022-05-13
医学概论(2022年4期)2022-04-24
中国药房(2022年7期)2022-04-14
现代青年·精英版(2022年3期)2022-04-06
电脑报(2020年40期)2020-11-06
农民致富之友(2019年26期)2019-10-11
小溪流(故事作文)(2018年10期)2018-12-28
