
灰飞虱G蛋白偶联受体基因的鉴定与分析
2022-08-11 03:47徐圣张怡萍韩召军吴敏
南京农业大学学报 2022年4期
徐圣,张怡萍,韩召军,吴敏
(南京农业大学植物保护学院,江苏 南京 210095)
G蛋白偶联受体(G protein coupled receptor,GPCR)是一类高度保守的膜蛋白,广泛存在于动物、植物和微生物中。GPCR的数目和种类众多,但是具有共同的特征拓扑结构:膜外N末端、7个跨膜螺旋结构域、膜内C末端、连接跨膜结构域的3个胞内环和3个胞外环[1-2]。Kolakowski[3]将GPCR分为6个家族:类视紫红质受体家族(A家族)、类分泌素受体家族(B家族)、代谢型谷氨酸受体家族(C家族)、真菌信息素P-和α-因子受体(D家族)、真菌A-和M-因子受体(E家族)和来自网柄菌的cAMP 受体(F家族)。迄今为止,昆虫中尚未有上述D、E和F家族成员的报道。昆虫中存在卷曲(frizzled)受体、平滑(smoothened)受体、bride of sevenless(boss)和stan受体等,它们与上述6个家族的GPCR序列相似性低,但是具有典型的GPCR拓扑结构,这些新型GPCR在果蝇中被归为非典型七次跨膜蛋白(D家族)[4]。因此,目前昆虫GPCR分为A、B、C和D四个家族[4-5]。
GPCR是联系细胞内、外信号的重要桥梁。GPCR被细胞外信号(胞外配体)激活后,在细胞内部启动第二信使引发级联反应,将信号从细胞外部传递到内部,并通过与其他信号通路之间的相互作用调控各种生物学功能,最终对生物体的生长发育、生殖、滞育、摄食、代谢以及行为等起到调控作用[6]。
灰飞虱(Laodelphaxstriatellus)是一类重要的农业害虫。灰飞虱的基因组是2019年发表的,总大小约为464 Mb,预计编码17 736个蛋白[15]。由于作者没有将基因注释信息公布,因此无法直接利用灰飞虱基因组数据搜索到灰飞虱的GPCR基因。截止到2021年9月,GenBank中仅能搜索到3个已注释的灰飞虱GPCR基因(登录号:BAO03861.1、BAO03862.1和BAO03863.1)。针对目前灰飞虱GPCR基因信息不明了的现状,我们利用灰飞虱基因组和转录组数据进行了搜索和鉴定,共鉴定到114个灰飞虱GPCR基因。本研究结果不仅完善了灰飞虱GPCR基因信息,而且为进一步研究灰飞虱GPCR的功能奠定了基础。
1 材料与方法
1.1 灰飞虱GPCR的鉴定方法
以下载的灰飞虱基因组数据(https://www.ncbi.nlm.nih.gov/assembly/GCA_003335185.2)和本试验前期测得的灰飞虱转录组数据构建本地数据库。以黑腹果蝇[4,16-17]和豌豆蚜[5,16]的GPCR序列作为参考序列,在本地数据库中进行TBLASTn和BLASTp搜索,得到灰飞虱候选GPCR基因集,在候选基因中去除冗余序列后,使用TMHMM(v2.0)(http://www.cbs.dtu.dk/services/TMHMM/)去除不含跨膜螺旋结构特征的候选基因,即获得灰飞虱GPCR基因。将所有鉴定的灰飞虱GPCR基因序列比对到NCBI的非冗余蛋白数据库,与已公开的其他物种GPCR序列进行比对。
1.2 灰飞虱GPCR序列分析
使用 NCBI CD-Search(http://www.ncbi.nlm.nih.gov/Structure/cdd/wrpsb.cgi)搜 索 保 守 结 构 域 数 据 库(conserved domain database,CDD),对灰飞虱GPCR基因序列进行跨膜结构域分析以初步判断其所属家族和亚家族;将其他昆虫的GPCR蛋白序列和灰飞虱GPCR序列一起采用Clustal W(http://www.ebi.ac.uk/Tools/msa/clustalw2/)软件进行多重比对,使用软件MEGA 6的邻接法(Neighbor-joining,NJ)和最大似然法(maximum likelihood,ML)生成系统发育树,NJ树采用校验参数Bootstrap重复1 000次,选择Pairwise deletion估算进化距离;ML树采用校验参数Bootstrap重复500次,Poisson模型选择Complete deletion估算进化距离,其他参数均采用默认设置。结果部分为NJ树。
2 结果与分析
2.1 灰飞虱GPCR基因鉴定
本文共鉴定了114个灰飞虱GPCR基因。其中3个已经在GenBank中报道,其余111个是新鉴定的灰飞虱GPCR基因。111个GPCR中有109个是从基因组数据库中搜索得到,另外2个GPCR基因(登录号:OK323198和OK323199)是从转录组数据中鉴定获得。从灰飞虱基因组中鉴定的109个基因中有5条序列(RZF36065.1、RZF36806.1、RZF43740.1、RZF48464.1和RZF45977.1)的完整性低于转录组中的对应序列,于是用转录组数据补充完整了这5条GPCR基因序列。
通过跨膜结构域分析发现:114个灰飞虱GPCR基因中有101个具有完整的7跨膜螺旋结构(占88.6%),其余13个GPCR可能是由于序列缺失,7跨膜螺旋结构并不完整。这13个GPCR基因中有7个具有6个跨膜螺旋结构(占5.2%),3个具有5个跨膜螺旋结构(占2.6%),3个只有4个跨膜螺旋结构(占2.6%)。
根据序列的特征结构,这114个GPCR分别归属于4个家族:81个A家族成员、19个B家族成员、7个C家族成员和7个D家族成员。灰飞虱A家族的GPCR数量最多,尤其是其中的神经肽和蛋白激素受体亚家族数量较多。该结果与其他已发表的昆虫GPCR基因分布情况类似(表1)。灰飞虱GPCR基因的GenBank登录号、基因名称、跨膜次数见表2。
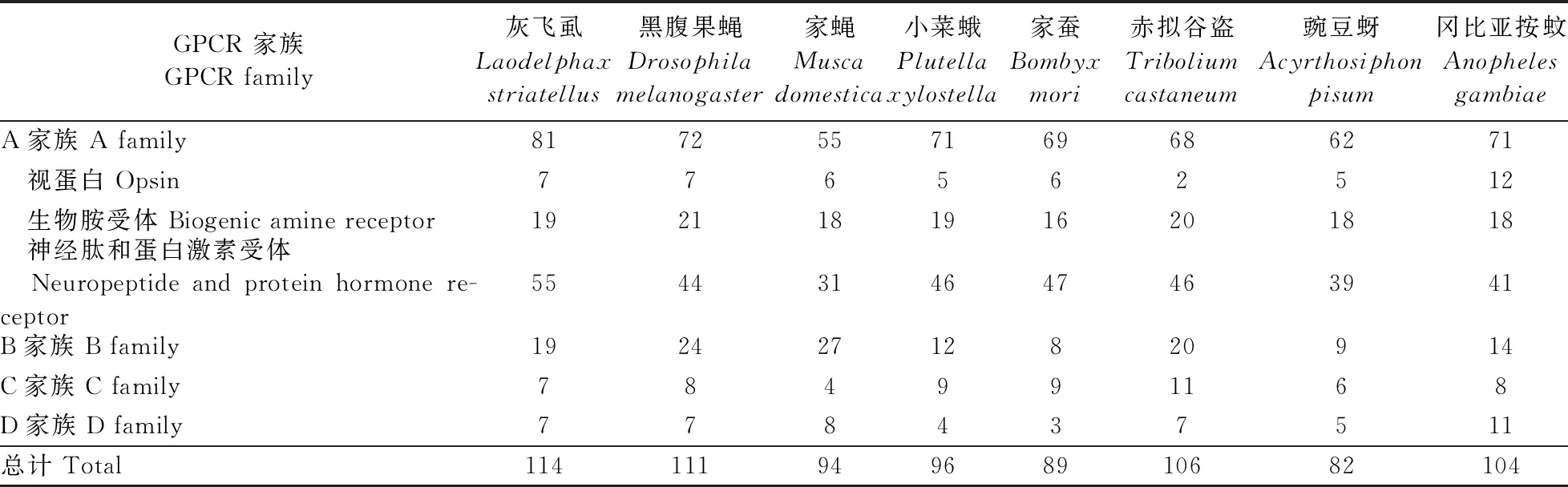
表1 灰飞虱与其他昆虫的GPCR基因数量Table 1 The number of GPCR genes in Laodelphax striatellus and other insects

表2 灰飞虱GPCR的鉴定Table 2 Identification of GPCR in L.striatellus

续表2 Table 2 continued

续表2 Table 2 continued
2.2 灰飞虱GPCR序列分析
2.2.1 灰飞虱GPCR A家族A家族GPCR可进一步分为视蛋白(opsin)、生物胺受体(biogenic amine receptor)及神经肽和蛋白激素受体(neuropeptide and protein hormone receptor)亚家族[4-5,11]。灰飞虱中鉴定了81个A家族GPCR,其中7个是视蛋白受体,19个是生物胺受体,55个是神经肽和蛋白激素受体。
昆虫的视蛋白有R型、C型和RGR/Go型[18-19]3类。R型视蛋白又可分为紫外光波敏感型视蛋白(ultraviolet sensitive opsin,UVO)、长波长敏感视蛋白(long wavelength sensitive opsin,LWO)、蓝光敏感视蛋白(blue sensitive opsin,BLO)、视紫质视蛋白(rhodopsin)和节肢动物视蛋白(arthropsin)[20-21]。灰飞虱中鉴定了5个R型视蛋白(2个UVO、1个LWO、1个Rhodopsin和1个Arthropsin),1个C型视蛋白与1个 RGR/Go型视蛋白,未发现BLO。将这7个灰飞虱视蛋白与其他物种的视蛋白进行序列比对并构建系统发育树(图1)。R型、C型和RGR/Go型3类视蛋白各自聚成1个进化支,R型视蛋白进化支中又细分为UVO、 LWO、BLO、Rhodopsin和Arthropsin 5个小支。

图1 基于氨基酸序列构建的灰飞虱和其他物种视蛋白的系统发育树Fig.1 Phylogenetic tree of opsins of L.striatellus and other species based on amino acid sequences Ls:灰飞虱Laodelphax striatellus;Ap:豌豆蚜Acyrthosiphon pisum;Dm:黑腹果蝇Drosophila melanogaster;Ev:假眼小绿叶蝉Empoasca vitis;Am:意大利蜜蜂Apis mellifera;Sf:秋赤蜻Sympetrum frequens;Lf:蜻蜓Ladona fulva;Oa:白尾灰蜻Orthetrum albistylum;Tc:赤拟谷盗Tribolium castaneum;Mp:桃蚜Myzus persicae. 下同。The same as follows. 以黑腹果蝇CG4521_mthl1作为发育树的外群。D. melanogaster CG4521-mthl1 receptor was the outgroup of the phylogenetic tree.
生物胺包括章鱼胺、酪胺、多巴胺、5-羟色胺和乙酰胆碱等。它们可以调节昆虫的许多生理过程,例如:昼夜节律、内分泌、学习和记忆等[8,22]。灰飞虱中鉴定了19个生物胺受体:6个章鱼胺受体(octopamine receptor)、5个5-羟色胺受体(5-hydroxytryptamine receptor)、3个多巴胺受体(dopamine receptor)、2个酪胺样受体(tyramine like receptor)、2个毒蕈碱型乙酰胆碱受体(muscarinic acetylcholine receptor,mAChR)和 1个孤儿受体(orphan receptor)。将这19个受体与黑腹果蝇和豌豆蚜的生物胺受体进行序列比对并构建系统发育树(图2)。灰飞虱比黑腹果蝇多了2个章鱼胺受体基因(登录号:RZF38146.1和RZF37066.1),少了1个酪胺样受体和1个毒蕈碱型乙酰胆碱受体基因;灰飞虱比豌豆蚜多了1个章鱼胺受体,2个5-羟色胺受体和1个多巴胺受体。灰飞虱中没有鉴定到多巴胺/蜕皮激素(dopamine/ecdysteroid,DopEcR)受体基因。
在白羽肉鸡养殖过程中,由于对白羽肉鸡相关传染性疾病的认识不到位,不重视白羽肉鸡相关传染性疾病的免疫接种,导致相关传染性疾病的发生与流行。一些传染性疾病在肉鸡养殖过程中流行,如新城疫、禽流感、传染性支气管炎等。这些传染性疾病主要以温和型发生与流行,发病症状较轻,死亡率较低,且不易诊断。这使传染性疾病成为潜在威胁,当外界环境条件改变时,可引起发病甚至死亡[2]。
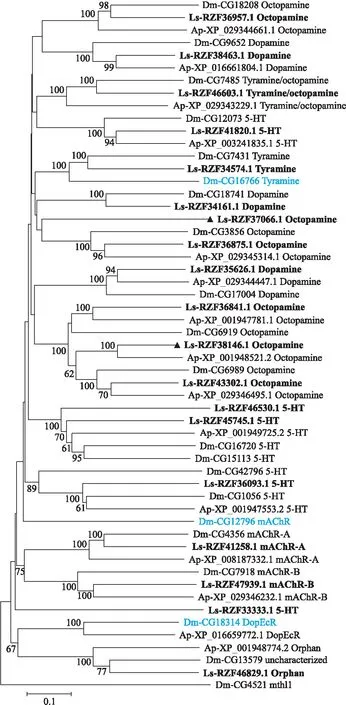
图2 基于氨基酸序列构建的灰飞虱及其他物种生物胺受体的系统发育树Fig.2 Phylogenetic tree of biogenic amine receptors of L.striatellus and other insect species based on amino acid sequences 黑色三角代表灰飞虱比黑腹果蝇多的章鱼胺受体;蓝色字体表示黑腹果蝇多于灰飞虱的GPCR。The black triangles represent more octopamine receptors in L.striatellus than in D. melanogaster. The blue fonts represent more GPCR in D. melanogaster than in L.striatellus.
神经肽和蛋白激素受体是GPCR受体中最大的亚家族。灰飞虱中共鉴定了55个神经肽和蛋白激素受体。从图3可知:灰飞虱的类速激肽受体、激肽受体、RYamide受体、Capability受体、脂动激素受体、甲壳动物心肌肽受体和糖蛋白激素受体基因存在明显的基因重复现象,这些受体基因数多于黑腹果蝇的同源受体基因数;灰飞虱中有黑腹果蝇中没有的促咽侧体激素受体、Elevenin受体和精氨酸加压素样肽受体;灰飞虱中没有鉴定到与黑腹果蝇同源的Trissin受体,豌豆蚜和褐飞虱中同样未鉴定到该受体;灰飞虱抑咽侧体激素受体、硫激肽受体和焦激肽受体基因数量都少于黑腹果蝇中相应受体基因数量。

图3 基于氨基酸序列构建的灰飞虱和其他昆虫神经肽(A)与蛋白激素受体(B)的系统发育树Fig.3 Phylogenetic trees of neuropeptide(A)and protein hormone receptors(B)of L.striatellus and other insect species based on amino acid sequences Nl:褐飞虱Nilaparvata lugen. 黑色三角代表灰飞虱比黑腹果蝇多的神经肽与蛋白激素受体;蓝色字体表示黑腹果蝇多于灰飞虱的GPCR。The black triangles represent more neuropeptide and protein hormone receptors in L.striatellus than in D.melanogaster. The blue fonts represent more GPCR in D. melanogaster than in L.striatellus.
2.2.2 灰飞虱GPCR B家族GPCR B家族受体是一类具有长N-末端结构域的受体。昆虫GPCR B家族可以进一步分为B1、B2和B3三个亚家族[23]。灰飞虱中共鉴定了19个B家族GPCR,其中7个属于B1亚家族,3个属于B2亚家族,9个属于B3亚家族。
灰飞虱中有7个B1亚家族受体:1个促肾上腺皮质激素释放因子(corticotropin releasing factor,CRF)受体、2个降钙素(calcitonin)受体、2个类甲状旁腺激素受体(parathyroid hormone receptor like,PTHRL)、1个神经元表达色素扩散因子(pigment dispersing factor,PDF)受体和1个孤儿受体。灰飞虱中发现了2个类甲状旁腺激素受体基因(RZF39724.1和RZF48464.1)与褐飞虱的类甲状旁腺激素受体基因B6(BAO01106.1)和B7(BAO01107.1)[24]的序列相似性高达94.86%和96.09%;黑腹果蝇和豌豆蚜中都没有发现这类受体基因。
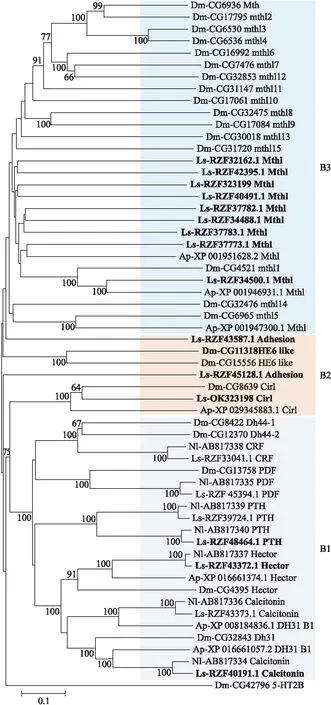
图4 基于氨基酸序列构建的灰飞虱和其他昆虫B家族受体的系统发育树Fig.4 Phylogenetic tree of family B receptors of L.striatellus and other insect species based on amino acid sequences
灰飞虱中鉴定到了3个B2亚家族受体基因(OK323198、RZF45128.1和RZF43587.1)。其中OK323198为Latrophilin like受体基因,它与黑腹果蝇中唯一的Latrophilin like受体基因(CG8639)同源,它们在系统发育树中聚为一小支(图4)。通过序列比对发现,RZF43587.1分别与小蜜蜂(Apisflorea)和湿木白蚁(Zootermopsisnevadensis) B2亚家族基因XP_012350908.1和XP_021914438.1的序列相似性为45.51%和46.67%。RZF45128.1具有GPCR蛋白水解位点(GPCR proteolysis site,GPS)的特征序列、钙结合EGF样结构域(calcium-binding EGF-like domain)的重复序列以及钙黏蛋白串联重复序列结构域(cadherin tandem repeat domain),这些都是B2亚家族中黏附类GPCR(adhension GPCR)的典型特征。
B3亚家族有Methuselah(Mth)和Methuselah like(Mthl)2类受体。灰飞虱中鉴定了9个Mthl基因,没有鉴定到Mth基因。序列比对发现,灰飞虱Mthl基因与果蝇的Mthl基因序列相似度都比较低。序列相似性最高的是灰飞虱RZF34500.1与黑腹果蝇NP_573140.1基因(相似性为40%)。灰飞虱中的Mthl基因数量(9个)少于黑腹果蝇的Mthl数量(16个),但是多于豌豆蚜的Mthl数量(3个)。
2.2.3 灰飞虱GPCR C家族GPCR C家族有γ-氨基丁酸B(GABA-B)受体和代谢型谷氨酸(metabotropic glutamate,mGlu)受体2类。灰飞虱中鉴定了2个GABA-B受体基因和5个mGlu受体基因。与黑腹果蝇相比,灰飞虱中少1个GABA-B受体基因,与豌豆蚜相比,灰飞虱多了2个mGlu受体基因(图5)。
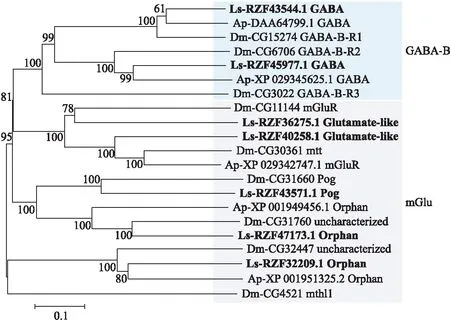
图5 基于氨基酸序列构建的灰飞虱和其他昆虫的C家族受体的系统发育树Fig.5 Phylogenetic tree of family C receptors of L.striatellus and other insect species based on amino acid sequences
2.2.4 灰飞虱GPCR D家族灰飞虱中鉴定出7个GPCR基因D家族,其中4个Frizzled受体基因,1个Smoothened受体基因,1个Stan受体基因和1个Boss受体基因。与黑腹果蝇相比,灰飞虱比黑腹果蝇多了1个Frizzled受体基因。与豌豆蚜相比,灰飞虱多了1个Frizzled受体基因和1个Stan受体基因(图6)。

图6 基于氨基酸序列构建灰飞虱和其他昆虫D家族受体的系统发育树Fig.6 Phylogenetic tree of family D receptors of L.striatellus and other insect species based on amino acid sequences 黑色三角代表灰飞虱比黑腹果蝇多的1个Frizzled受体。The black triangle represents one more Frizzled receptor in L.striatellus than in D. melanogaster.
3 讨论
关于多基因家族是如何演化的问题,科学家提出了协同进化(concerted evolution)理论和生死进化(birth-and-death evolution)理论等。其中,生死进化理论被认为是引导多基因家族长期进化的主要机制。该理论认为:新的基因是通过基因复制产生的,这些重复的基因一些可以长期留在基因组中,而另一些则被删除或通过有害的突变不能发挥作用[25-26]。昆虫在进化过程中发生了大量的基因重复和丢失事件,基因重复或丢失被认为与基因的新功能和物种的适应性等密切相关[27]。
本研究中,与黑腹果蝇相比,灰飞虱GPCR基因出现了可能的缺失现象。例如:Trissin受体基因、多巴胺/蜕皮激素受体基因和Mth基因在灰飞虱中都没有被鉴定到;酪胺样受体基因、毒蕈碱型乙酰胆碱受体基因、抑咽侧体激素受体基因、硫激肽受体、焦激肽受体基因和GABA-B受体基因的数量都比黑腹果蝇中相应的受体基因数量少。灰飞虱中未找到这些基因可能有3个原因:1)这些GPCR基因真的在灰飞虱基因组中发生了丢失,它们对灰飞虱是非必需的,因此在进化过程中丢失了。2)灰飞虱本身并没有复制产生这些基因,因为半翅目昆虫灰飞虱在进化上是早于黑腹果蝇的[28]。3)由于测序技术和序列拼接的问题导致数据的缺失和不完整,因此无法鉴定得到目标GPCR基因。灰飞虱中这些GPCR基因的缺失是否真正发生丢失仍需要进一步的研究和验证。
本研究中,与黑腹果蝇相比,灰飞虱GPCR各个家族中都出现了基因重复的现象,尤其是A家族神经肽与蛋白激素受体中的脂动激素受体、Capability受体、RYamide受体、甲壳动物心肌肽受体、类速激肽受体、激肽受体和糖蛋白激素受体基因的数量显著多于黑腹果蝇中的同源GPCR基因的数量。已有报道表明,脂动激素及其受体参与调控脂类代谢与糖类的平衡,也通过抑制脂肪体中蛋白质合成而影响生殖行为,还参与逆境生理与免疫反应等生命活动[29]。由于灰飞虱寄主广泛,并能以休眠或滞育状态进行越冬,推测脂动激素受体基因的多拷贝可能增强灰飞虱对不同寄主和温度的适应能力。Capability多肽已被证明能促进不同种类昆虫的马氏管分泌液体[30]。同时Capability及其受体在肌肉调节和渗透调节中发挥重要作用,Capability会协同其他神经肽提高果蝇对低温和干燥环境的耐受能力[31]。因此,我们推测灰飞虱Capability受体基因的扩增,可能有助于灰飞虱对低温环境的适应,协助灰飞虱顺利越冬。目前对RYamide受体的研究表明,RYamide可能在昆虫摄食行为和体内水平衡等生理活动中发挥重要作用[32-33]。因此,推测灰飞虱RYamide受体基因的重复可能在灰飞虱取食行为和对体内水平衡的调控中发挥较大的作用。赤拟谷盗中鉴定了2个甲壳动物心肌肽受体ccapr-1和ccapr-2,它们均具有功能活性,但ccapr-1受体在赤拟谷盗中不发挥作用,可能是一种功能备份[34]。灰飞虱中存在3个甲壳动物心肌肽受体,它们的功能可能更加复杂,仍需进一步的研究。
GPCR信号调控系统十分复杂,参与众多生理过程和行为的调节。本文通过鉴定和分析灰飞虱GPCR受体基因,发现与黑腹果蝇的GPCR相比,灰飞虱GPCR受体基因不仅种类丰富,而且存在较多的基因扩增和可能的丢失情况。本研究不仅完善了灰飞虱GPCR基因的信息,而且为灰飞虱GPCR的功能研究奠定了基础。
猜你喜欢
上海农业科技(2022年4期)2022-08-15
热带作物学报(2022年5期)2022-06-06
热带农业科学(2022年5期)2022-06-02
学苑创造·A版(2022年3期)2022-03-29
中国听力语言康复科学杂志(2021年6期)2021-12-21
福建基础教育研究(2019年2期)2019-09-10
学苑创造·A版(2019年6期)2019-07-11
农业与技术(2019年6期)2019-05-29
福建基础教育研究(2019年2期)2019-05-28
职业·下旬(2009年8期)2009-10-12
