
卧龙国家级自然保护区川金丝猴的群落环境分析
2022-08-11 02:35刘卓涛王晓娟周厚熊施小刚李兆元
野生动物学报 2022年3期
刘卓涛 钟 义 王晓娟 李 谦 周厚熊 施小刚 李兆元*
(1.西南林业大学地理与生态旅游学院,昆明,650224;2.中国野生动物保护协会,北京,100714;3.昆明逸境生态有限公司,昆明,650224;4.卧龙国家级自然保护区管理局,汶川,623006)
川金丝猴(Rhinopithecusroxellana)属于灵长目(Primates),猴科(Cercopithecidae),是中国特有物种,主要分布于陕西秦岭、湖北神农架、甘肃南部以及四川邛崃山、凉山和岷山几大山系[1-2],目前已被IUCN列为濒危(EN)物种,是国家一级重点保护野生动物[3]。川金丝猴主要取食植物的叶、芽、果、树皮以及地衣等,食谱存在季节性变化[4-5]。
种间生态学关系是群落生态学研究的核心,开展种间空间关联的研究是发现种间生态学关系全面且有效的方法。当代种间空间关联研究主要集中在植物群落,对动物群落的研究较少,涉及的类群有鱼类[6-7]、啮齿类[8]、鸟类和灵长类[9],这些研究局限于种对关系的空间关联分析,没有对群落中的物种网络进行多维度的比较分析。基于这种研究现状,本课题组从2018年开始,用红外相机数据计算同一研究区中物种的佛爱系数以衡量种间关联性,以此构建四川卧龙地区生物群落中的地栖动物关联物种网络。初步分析发现,卧龙地区存在2个地栖温血物种网络,一个包括羚牛(Budorcastaxicolor)等20种动物,位于低海拔地区(海拔<3 531 m)[10];另一个包括雪豹(Pantherauncia)等13种动物,分布在高海拔地区(海拔3 531~4 430 m)[11]。川金丝猴生活于低海拔网络中,本研究基于物种关联网络分析其群落环境,旨在丰富群落生态学的理论知识,为保护管理措施的改进提供依据。
1 研究区概况
卧龙自然保护区位于四川省阿坝藏族羌族自治州汶川县西南部(30°45′—31°25′ N,102°52′—103°24′ E),属于邛崃山系东麓,总面积2 000 km2,地势西北高、东南低,相对高差5 100 m[12]。保护区1月平均气温为-1.7 ℃,7月17 ℃;年降水量为861.8 mm,集中在5—9月[13]。巨大高差形成丰富的立体气候环境和发育良好的垂直植被带:从山谷到山顶依次为亚热带(1 600 m以下)、温带(1 600~2 600 m)、寒温带(2 600~3 600 m)、寒带(3 600~4 400 m)、高寒带(4 400~5 000 m)和极高山寒冻冰雪带(>5 000 m),植被依次为常绿阔叶林、常绿落叶阔叶混交林、落叶阔叶林、温性针叶林、温性针阔混交林、寒温性针叶林、耐寒灌丛、高山草甸和高山流石滩稀疏植被[14]。丰富的植被和生境类型孕育着丰富的动物种类,有脊椎动物504种,其中国家一级重点保护野生动物14种,二级保护动物51种[15]。代表物种有大熊猫(Ailuropodamelanoleuca)、川金丝猴和雪豹等。
2 研究方法
2.1 数据采集与提取
采用红外线相机调查方法采集数据。卧龙自然保护区是一个山地环境,许多区域人无法进入,根据卧龙自然保护区的边界,利用ArcGIS生成覆盖保护区的1 km×1 km的公里网格,在可通达的地方共布置了400台红外相机,每个网格安装了1台以上的相机。沿着海拔随机抽取60个相机位点,覆盖1 600~4 400 m,这个范围大致可以分为3个区域:高海拔区域(3 100~4 400 m)、中海拔区域(2 500~3 400 m)和低海拔区域(1 600~2 800 m)。相机固定在距离地面50~80 cm的树干或其他固定物上,镜头基本与地面保持平行,工作时间为2017年2月—2018年4月。将相机数据回收后,根据相机编号与内存卡编号,逐一查看照片和视频,对物种进行分类与鉴定。物种鉴定参照《中国兽类野外手册》[16]和《中国鸟类野外手册》[17]。
2.2 数据分析
2.2.1 种对关联性分析及网络构建
在进行种对关联性分析时,采用经过数理论证的统计学方法来衡量种对关联性,选取具有显著正相关的种对进行分析。由于各台红外相机获得的是二分型数据,因此采用佛爱系数(rø)计算物种间关联系数[18]。
2.2.2 非对称关联分析
本研究数据为二分型,为了探讨物种间的非对称关联,先建立列联表,分析物种两两间的非对称关联,列联表与佛爱系数列联表完全一致,用兰布达系数(LB)[18]计算物种A(从偏利中不获益,甚至受害)对物种B(从偏利中获益的物种)的预测性,即预测率上限。
(1)
式中:j为列数;i为行数;N为相机位点数;nMj为第j列的最大列频率;max(Ri)为最大行总和。
兰布达系数的显著性检验,首先计算LB的变异性,如下:

2∑′nMj)]/[N-max(Ri)]3
(2)
式中:∑′nMj为落在max(Ri)所在行的最大列频率之和。若落在该行上的最大列频率只有1个,则∑′nMj=nMj。最后计算z,对于LB,有:
(3)
式中:λB0为预测物种B的出错率下降幅度等于某个具体数值,本研究取对应显著性P=0.05时z值的λB0。
(4)
式中:LB为公式1计算值,zP=0.05可以从统计学书籍的数表中查阅获得。计算得出的λB0代表物种A对物种B有显著性意义的预测率下限。
2.2.3 物种关联复杂性分析
用香农维纳指数(Shannon-Wiener)来衡量物种关联网络的整体复杂性和物种关联复杂性,计算如下:
(5)
(6)
当分析特定物种关联复杂性时,式中,H′为该物种的香农维纳指数,S为与该物种存在直接正关联的种对总数,Pi为种对i的关联系数占该物种关联系数总和的比例。当分析整个网络的关联复杂性时,式中H′为该网络的香农维纳指数,S为该网络所有存在显著正关联的种对总数,Pi为种对i的关联系数占该网络所有关联系数总和的比例。
为了进一步分析物种对整个网络维系的作用以及网络中物种的层级,进行如下计算:
(7)
式中:C为该物种对网络的贡献率;H′值越大,则物种空间关系越复杂。特定物种的香浓维纳指数占网络香浓维纳指数的比例用做该物种对网络稳定性的贡献率,比例越大,贡献率越高,物种对群落稳定性的维系能力就越强。对于网络中物种的层级,将各物种关联复杂性对整个网络复杂性的贡献率进行平均,然后将贡献率高于平均值的物种归为网络的内层物种,低于平均值的物种归为外层物种。
3 结果与分析
3.1 种间关联性
红外相机共记录到35种地栖动物,构成2个物种关联网络。在川金丝猴所在的网络(图1)中,将各物种关联复杂性对整个网络复杂性的贡献率进行平均,然后将贡献率低于平均值的物种归为网络的外周物种(表1),包括藏酋猴(Macacathibetana)、花面狸(Pagumalarvata)、羚牛、豹猫(Prionailurusbengalensis)、勺鸡(Pucrasiamacrolopha)、狼(Canislupus)、红腹锦鸡(Chrysolophuspictus)和中华斑羚(Naemorhedusgriseus),这些物种的显著正关联物种对<5个,关联系数平均为0.36(0.31~0.41),香农维纳指数平均值为0.57(0~1.38)。贡献率高于平均值的物种归为核心物种,包括毛冠鹿(Elaphoduscephalophus)、黄喉貂(Martesflavigula)、水鹿(Rusaunicolor)、野猪(Susscrofa)、小熊猫(Ailurusfulgens)、红腹角雉(Tragopantemminckii)、川金丝猴、中华鬣羚(Capricornismilneedwardsii)、豪猪(Hystrixbrachyura)、林麝(Moschusberezovskii)、大熊猫和黑熊(Ursusthibetanus),这些物种的显著正关联物种对≥5个,关联系数平均为0.49(0.29~0.72),香农维纳指数平均值为2.01(1.61~2.38),其中,川金丝猴的香农维纳指数为1.92,贡献率为6.75%,处于第7位,在核心物种中处在中间位置,表明川金丝猴是群落维持的重要物种之一,但不是最重要的物种。
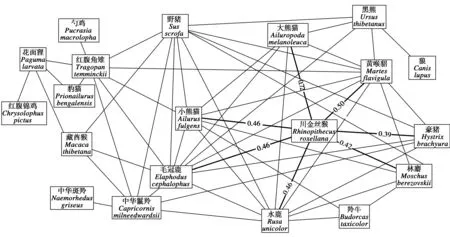
图1 卧龙国家级自然保护区物种空间关联网络结构

表1 卧龙低海拔物种关联网络中香农维纳指数及贡献率
在这个网络中,有7个物种与川金丝猴存在直接显著正关联,分别为毛冠鹿(rø=0.43,χ2=9.55,P<0.01)、黄喉貂(rø=0.50,χ2=12.89,P<0.01)、水鹿(rø=0.46,χ2=10.36,P<0.01)、小熊猫(rø=0.46,χ2=10.53,P<0.01)、豪猪(rø=0.39,χ2=7.41,P<0.01)、大熊猫(rø=0.72,χ2=27.68,P<0.01)与林麝(rø=0.42,χ2=8.47,P<0.01)。
3.2 关联物种间的非对称关系
在与川金丝猴发生直接空间关联的7个物种中,种对间出现非对称空间关联:黄喉貂、林麝、小熊猫和大熊猫对川金丝猴有预测,预测率分别为13%~39%、9%~30%、10%~35%和47%~65%;川金丝猴对大熊猫和黄喉貂有预测,预测率分别为31%~58%和3%~33%(表2)。在这些非对称关联中,川金丝猴和黄喉貂,以及川金丝猴和大熊猫之间存在分布上的相互预测性,或者是相互吸引对方,其中,大熊猫对川金丝猴的预测性最强。

表2 川金丝猴与其他直接关联物种的预测性(兰布达系数检验)
4 讨论
4.1 川金丝猴的互利关系
研究结果显示,大熊猫和黄喉貂都与川金丝猴有相互预测性,说明川金丝猴与大熊猫和黄喉貂在分布上存在相互吸引性,它们之间可能存在互利生态学关系。
4.1.1 川金丝猴-大熊猫
川金丝猴和大熊猫都偏向选择针阔混交林[19-20],川金丝猴主要在乔木林冠中活动[21],而大熊猫主要在林下灌丛中活动,活动基底实际上分离。川金丝猴的主要食物在大熊猫活动区域的乔木林中十分丰富[4,22],但由于食性不同,2个物种在食物上也不存在竞争关系。大熊猫觅食会降低竹林密度[23],促进乔木生长,从而有利于川金丝猴,这可能是大熊猫对川金丝猴在分布空间上产生吸引力的原因,即川金丝猴倾向于分布在有大熊猫的区域中。而川金丝猴的取食对乔木冠层郁闭度存在稀疏化作用,有利于竹子生长,为大熊猫提供更为丰富的食物资源,这可能是川金丝猴对大熊猫在分布空间上产生吸引力的原因,即大熊猫倾向于分布在有川金丝猴分布的区域中。通过上述机制,川金丝猴和大熊猫实现生存上的互利,因此,对任何一个物种的有效保护都会有利于对另一个物种的保护。
4.1.2 川金丝猴-黄喉貂
黄喉貂为杂食性食肉物种,食物包括昆虫、鸟类、哺乳类和植物,尤其是浆果[24],在果成熟的秋季优先食果[25]。2种动物都会取食猕猴桃科(Actinidiaceae)和蔷薇科(Rosaceae)的果[4,25],黄喉貂食果有利于这些植物的散布[26],对以乔木为生活基底的川金丝猴的生存是有利的。食性需求的相似性导致2个物种倾向于分布在相同的地理空间中,并形成统计学上的空间关联性。当一方的存在为另一方带来好处时,获益方在分布上倾向于出现在对方的空间里,从而形成非对称空间关联;当双方的存在都为对方带来好处时,2个物种都倾向于分布在对方的空间中。但是,从引述的生态学研究文献中只发现黄喉貂的活动对川金丝猴的意义,未发现川金丝猴对黄喉貂的意义,与本研究结果所预测的互利生态学关系不同。因此,笔者认为川金丝猴和黄喉貂之间存在着目前未知的互利生态学关系,有待生态学者进一步研究发现。
4.2 川金丝猴的偏利关系
研究结果显示,林麝和小熊猫都对川金丝猴有预测性,说明川金丝猴尾随林麝与小熊猫分布,并从与这2个物种的关系中获益,存在偏利关系。
4.2.1 川金丝猴-林麝
川金丝猴和林麝的空间关联系数为0.42,林麝对川金丝猴的预测率为9%~30%,表明川金丝猴从与林麝的关系中获益。川金丝猴和林麝主要分布在中海拔地区的针阔混交林中,偏爱乔木郁闭度适中、乔木密度较高、灌木及竹林适中且隐蔽性好的区域[27-28],并以木本植物为食[4,29],冬季选择积雪较浅的阳坡或半阳坡[30-31],可能是这些因素导致2个物种产生空间重叠。已有研究并没有揭示川金丝猴和林麝间任何偏利生态学关系,川金丝猴从与林麝的关系中受益的机制有待进一步观察。
4.2.2 川金丝猴-小熊猫
川金丝猴和小熊猫的关联系数(rø=0.46,χ2=10.53,P<0.01)较强,而且存在非对称关联(LB=0.348,λB0=0.102,P≤0.05),小熊猫对川金丝猴存在10%~35%的预测率,表明小熊猫的存在对川金丝猴有利,因此川金丝猴依据小熊猫而分布。川金丝猴与小熊猫都偏向选择针阔混交林下灌丛[19-20],然而,2个物种的活动基底不同,川金丝猴主要在乔木林树上活动[21],小熊猫则在林下竹丛中。食性差异降低了2种动物的竞争性。竹子生长会抑制乔木幼苗的生长[32]。小熊猫倾向于进入竹子密度较高、乔木相对较少的区域活动[33],通过取食竹子将竹林稀疏化[23],继而促进乔木生长,创造对川金丝猴有利的生境。笔者推测这种机制可能导致川金丝猴倾向于分布在有小熊猫活动的区域,从而形成非对称空间关联。
4.3 川金丝猴与其他物种的关系
川金丝猴还与水鹿、毛冠鹿和豪猪存在对等空间关联。在对水鹿的群落环境研究中发现,水鹿和川金丝猴由于活动基底和食物分布空间的相似性产生了空间关联,从而构成二者间的生态学关系[34]。
4.3.1 川金丝猴-毛冠鹿
毛冠鹿主要取食草本植物和竹子,在灌丛生长的季节里,为了食物与躲避天敌,毛冠鹿进入灌丛较密的区域活动,取食竹笋[35],这些区域也是川金丝猴冬春季的觅食场所[36],这可能是出现空间关联的部分原因。
4.3.2 川金丝猴-豪猪
豪猪主要选择森林、灌丛和草原等区域活动,主要取食植物根、块茎、树皮、叶以及落地的果[16,37],与川金丝猴的食性有一定程度相同。食性相似性导致2个物种出现稳定但不太强的空间关联。
4.4 川金丝猴在群落维持中的作用
川金丝猴作为核心物种,直接关联的物种有7个,各关联系数均较高,是维持群落的重要物种之一。在物种网络结构中,川金丝猴与大熊猫和黄喉貂存在互利生态学关系,对这3种动物中任何一种的有效保护可能惠及其余两种。川金丝猴从与林麝、小熊猫的生态学关系中获益,对林麝和小熊猫的有效保护有助于川金丝猴的生存。川金丝猴与水鹿、毛冠鹿和豪猪之间存在适度生态位分离,同时又有资源竞争。通过与其他物种的非对称关联,川金丝猴在加强群落内种间关联力度、群落内聚力度及稳定性具有重要作用,而基于对称关联,水鹿、毛冠鹿和豪猪过度的种群增长可能会排挤川金丝猴的生存空间。川金丝猴不是一个仅从群落中获益的物种,对维持群落也具有重要意义,出于物种多样性保护的目的,与强调对川金丝猴的单种保护措施相比,综合且平衡的保护措施可能是更好的保护策略。
致谢:卧龙国家级自然保护区王鹏彦、胡强、王茂麟、林红强,以及四川省林业和草原局古晓东给予了数据采集中的大力支持,姜楠、吴艳蓉和杨虎参与了数据采集和处理,刘萍在外业数据采集中提供后勤保障支持。特此感谢!
猜你喜欢
社会科学战线(2022年9期)2022-10-25
小哥白尼(野生动物)(2022年6期)2022-08-17
中学生数理化(高中版.高考数学)(2022年4期)2022-05-25
交通科技与管理(2022年8期)2022-05-07
云南画报(2021年10期)2021-11-24
学苑创造·A版(2021年9期)2021-09-16
东方少年·布老虎画刊(2021年1期)2021-06-22
广东教学报·教育综合(2020年9期)2020-04-17
学生天地·小学低年级版(2018年12期)2018-01-15
作文周刊·小学一年级版(2017年35期)2017-10-18
