
丹顶鹤的保护遗传学与保护基因组学研究
2022-08-11 02:24李菲凡胡东宇
野生动物学报 2022年3期
李菲凡 胡东宇
(沈阳师范大学,辽宁古生物博物馆,辽宁省东北亚生物进化与环境变迁重点实验室,沈阳,110034)
丹顶鹤(Grusjaponensis)为鹤形目(Gruiformes)鹤科(Gruidae)大型鸟类,目前被世界自然保护联盟(IUCN)濒危物种红色名录列为易危(VU)物种,也是国家一级重点保护野生动物[1],野外现存数量不足4 000只[2]。
作为一地区性分布物种,历史上丹顶鹤夏季广泛繁殖于东北亚地区的大型湿地,冬季迁徙到长江中下游地区、朝鲜半岛和日本本州甚至更南部地区越冬[3-5],目前野生丹顶鹤分为大陆和岛屿2个种群。大陆种群(约2 000只)夏季主要在俄罗斯远东与中国东北交界地区繁殖,冬季在中国东部沿海和朝鲜半岛的非军事区越冬。黑龙江省境内的松嫩平原和三江平原湿地是国内丹顶鹤的主要繁殖地,江苏省盐城沿海地区和山东省境内的黄河三角洲地区是国内丹顶鹤的主要越冬地[2,4,6]。岛屿种群(约1 900只)终年生活于日本北海道东部地区,春秋仅有越冬地和繁殖地间的短距离移动,所以也被称为日本北海道种群[2-3,6]。
由于大规模的土地开发和大量捕杀,至19世纪末,分布于日本的丹顶鹤数量急剧减少,曾一度被认为已经灭绝。目前的岛屿种群来自20世纪20年代在北海道东部的未开发湿地中仅存的30余只丹顶鹤[3]。经过保护,特别是50年代开始的冬季人工投食,岛屿种群数量逐渐得到恢复,至2020年发展到1 900只[7]。由于从小种群发展而来,一定程度上经历了种群瓶颈效应,因此岛屿种群的遗传多样性一直令人担忧[6-8]。对大陆种群实施的保护始于20世纪80年代,在我国境内越冬的丹顶鹤于20世纪末增加到接近1 200只,此后不断减少,目前已不到500只[9],同时栖息地也在不断地缩小和破碎化[10-12],大陆种群是否会重蹈岛屿种群的覆辙也令人担忧。
遗传多样性是物种演化的基础,保护遗传学研究为濒危物种保护提供理论依据。在过去的丹顶鹤保护过程中,虽然面临着样本获取困难和测试费用昂贵等难题,但还是有一些与保护遗传学相关的研究被开展。21世纪以来,随着高通量测序技术,特别是二代和三代测序技术的应用,保护基因组学研究应运而生,为濒危物种的科学保护提供了前所未有的机遇[13]。本文回顾了过去40年对丹顶鹤全面实施保护以来,在丹顶鹤保护遗传学与保护基因组学研究上已取得的成果,探讨了目前丹顶鹤大陆种群遗传多样性保护所面临的挑战,以及新的基因组学技术对今后丹顶鹤保护的潜在作用。
1 丹顶鹤的保护遗传学研究
保护遗传学(conservation genetics)是保护生物学和分子遗传学相互渗透、结合而产生的一门分支学科,已成为保护生物学研究的一个核心部分[14],主要研究内容包括:物种的分类地位、种群遗传结构和遗传多样性、近亲繁殖、遗传变异、基因流、杂交、迁移、亲系关系、有效种群大小、种群的亚分化和进化显著单元等方面[14]。在过去40年里,丹顶鹤保护遗传学研究主要集中在系统演化、种间关系和物种的遗传多样性等方面。
1.1 丹顶鹤在鹤类系统演化中的位置
关于丹顶鹤所在鹤科15种鹤的种间关系,前后有通过DNA-DNA杂交[15]、蛋白质电泳分析[16]、线粒体细胞色素b基因序列[17-18]、线粒体细胞色素b和NADH脱氢酶亚基6基因序列测序[19]、多基因(4个线粒体基因和3个核基因)序列测序[20-21]和线粒体全基因组序列测序[22]等进行的多项研究,这些研究得出了基本一致的结果(图1)。系统发育分析进一步支持了鹤科的单系性,并且其内部存在着种组(species group)分化,丹顶鹤、灰鹤(G.grus)、白头鹤(G.monacha)、美洲鹤(G.americana)、黑颈鹤(G.nigricollis)形成了一个种组,被称为美洲鹤种组(species group Americana)[22]。基于线粒体DNA序列,对鹤科各物种的分异时间所做的贝叶斯估计显示,各物种的分异发生在新近纪,丹顶鹤于7.9百万年~9.0百万年前最早从美洲鹤种组分出(图1)。主要用鹤属线粒体全基因组进行的系统演化分析[23]也得出了相同的结果。
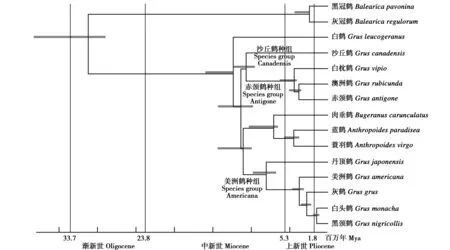
图1 基于全线粒体DNA序列的鹤类系统树(基于Krajewski等[22]修改)
1.2 岛屿种群与大陆种群间遗传多样性差异
关于丹顶鹤岛屿种群与大陆种群间的遗传多样性差异,目前主要通过线粒体DNA片段测序和以微卫星为主的核遗传标记分析进行研究。各种研究共同表明,丹顶鹤岛屿种群与大陆种群(不论是野生种群,还是人工饲养种群)相比,具有较低的遗传多样性水平,并且在过去的近百年间一直维持着一个较低的遗传变异水平。
1999年,日本学者Hasegawa等[8]对来自日本各动物园、水族馆的丹顶鹤等鹤类的线粒体DNA控制区(D环区)全DNA序列进行了测序,比较种间和种内的序列变化,并使用其中最易变的418 bp序列评估丹顶鹤大陆与岛屿种群间的遗传多样性差异。研究显示,丹顶鹤、灰鹤和白头鹤的种内遗传距离相似。在丹顶鹤大陆种群中发现了7个D环单倍型,而岛屿种群只有2个(Gj1、Gj2),两者间不存在相同的单倍型。简约网络(parsimony network)分析显示,岛屿种群的单倍型并没有形成独立的分支,系统发育树也显示2组单倍型没有按种群分成2个单系群,强烈地支持了2个种群属于一个谱系,无显著的遗传结构分化。Hasegawa等[8]认为,岛屿种群的单倍型较少,可能是由该种群从少数个体恢复而来所产生的“瓶颈”效应所致。此后,一个覆盖了岛屿种群各主要繁殖地的更大样本的测序分析证实,岛屿种群仅有Gj1和Gj2两个单倍型,并且以Gj2为主,Gj1单倍型在岛屿种群中的分布存在着地域局限性[24]。为了解岛屿种群遗传结构在时间上的变化,Akiyama等[25]从日本各博物馆、研究机构等获取1878—2001年保存的填充标本的羽毛,并使用1970—2014年在岛屿种群主要繁殖地区采集的幼鸟或亚成鸟血液或组织样本,对线粒体D环区DNA进行测序,在已知的2种单倍型以外,又发现了一种新的单倍型Gj13,来自1997年和2007年的各1份样本。分析结果表明:近百年来,Gj1和Gj2单倍型的频率没有明显变化,Gj2频率最低时(20世纪90年代)也达到了87.5%,Gj1频率最高时(20世纪80年代)仅11.1%,即岛屿种群一直维持着低遗传变异水平,而极其稀少的Gj13目前是否还存在尚不清楚。这也意味着岛屿种群的遗传多样性几乎不可能靠自身独立恢复,因此建议促进大陆和岛屿种群间的遗传交流,如人工引入大陆种群个体提高岛屿种群的遗传多样性水平[25]。
与使用线粒体DNA片段的分析同期,核遗传标记也被用于丹顶鹤大陆与岛屿种群的遗传多样性分析(表1)。Hasegawa等[26]从来自日本各动物园、水族馆饲养的大陆个体和北海道的野生幼鹤的DNA中分离出7个微卫星基因座,都表现出明显的等位基因变异,大陆个体表现为2~10个等位基因,岛屿种群则为1~4个等位基因,显示大陆种群存在着明显的等位基因变异,而岛屿种群的等位基因变异幅度相当小。对我国扎龙保护区野生、散养和圈养的3个丹顶鹤小种群应用6个微卫星分子标记位点的分析表明,3个小种群均表现出较高且相近的遗传变异水平[27];另外,从该保护区野生或散养个体的羽毛和蛋壳中,成功分离出12个微卫星基因座,并证实包括这些基因座在内的18个微卫星标记的每个组合对种群遗传分析都是有效的[28]。对采自我国盐城保护区的野生越冬丹顶鹤的羽毛样本选择6个微卫星基因座进行基因分型,获得了47个等位基因,所有微卫星位点均高度多态;同时,该研究对上述样本的线粒体细胞色素b基因序列也进行了分析,检测出12个单倍型,系统发育分析显示样本的同源性较低[29]。基于34个基因座,对来自成都、上海、重庆和秦岭动物园的丹顶鹤样本检测,发现其基因分型在209个等位基因中表现出高度的多态性[30]。上述研究均显示大陆种群具有高度的遗传多样性。

表1 不同研究分离的丹顶鹤种群微卫星基因座的变异分析
1.3 亲子鉴定与家族区域地理分布

利用1995—2006年在北海道不同巢址采集的幼鹤血液样本和12个多态性微卫星基因座,对北海道3个主要繁殖区的种群遗传结构的研究表明,3个区域种群间的遗传多样性没有显著差异,但似乎不是随机婚配的种群[31]。空间自相关分析(spatial autocorrelation analysis)显示,尽管丹顶鹤有很强的飞行能力,近距离(0~15 km)样本间有明显的正亲缘关系,而远距离(155~205 km)样本间则有明显的负亲缘关系,说明在近距离的空间尺度下仍存在距离隔离。在出生地的留恋(natal philopatry)影响下的扩散和繁殖地选择可能是这种距离隔离的主要成因[31]。
2 丹顶鹤的保护基因组学研究
保护基因组学(conservation genomics)本质上是保护遗传学理论和基因组学技术的融合。它通过高通量测序技术,尤其是二代、三代测序技术的应用,推动了保护遗传学研究内容和手段的革新性变化[33-34]。保护基因组学除了在传统的保护遗传学问题上能够提供更加精准的研究结果以外,还能对物种的演化历史、种群的局域适应、遗传混合、杂交、远交衰退和基因渐渗等方面提供更加深入的研究[33-34]。
目前,有关丹顶鹤的保护基因组学研究还处于起步阶段。近年有2项研究[22-23]分别测定了丹顶鹤线粒体全基因组序列,得到了基本一致的结果。测定的序列全长分别为16 715、16 727 bp,有13和14个蛋白质编码基因、22个tRNA基因和2个rRNA基因。2020年,丹顶鹤的全基因组序列被发表,用于研究基因与鸟类长寿的关系,这项研究鉴定出了多个与丹顶鹤长寿相关的候选基因,包括在代谢和免疫途径中被正向选择的基因,提供了低代谢率和长寿相关的基因证据[35]。通过与其他鸟类全基因组的比较进化分析(comparative evolutionary analysis),估测出丹顶鹤全基因组大小为1.146 Gb,鉴定出约3 600万个单核苷酸变异(SNV),核苷酸多样性为每位点1.29×10-3个核苷酸取代。与无危鸟类物种的核苷酸多样性中位数(每位点2.49×10-3个核苷酸取代)相比,丹顶鹤的核苷酸多样性水平低,在遗传上与濒危物种一致。用成对顺序马尔科夫聚类(PSMC)分析方法对丹顶鹤的历史种群数量波动进行了模拟,显示从上新世晚期到更新世早期(约40万年前),丹顶鹤的有效种群数量(Ne)减少;在晚更新世的最后一个冰期末期(70万~10万年前)增加。Ne在大约8万年前达到峰值,预计约7.0×104[35]。
3 结论与展望
3.1 结论
回顾40年来的丹顶鹤保护,保护遗传学研究主要集中在物种的系统演化和遗传多样性,以及岛屿和大陆种群间的遗传多样性差异方面。在技术应用上,证实了AFLP和微卫星分子标记可用于丹顶鹤亲缘关系鉴定。保护基因组学研究虽历史较短,但已完成了线粒体全基因组和全基因组序列的测定。以上研究显示:(1)现存15种鹤类组成了一个单系群,丹顶鹤属于美洲鹤种组,是该种组中最早分化出来的物种。从分化时间来说,该种组称为丹顶鹤种组应该更为恰当。(2)丹顶鹤物种的核苷酸多样性水平较低,在遗传上与濒危物种一致;岛屿和大陆种群之间,虽然不存在遗传结构上的显著分化,但岛屿种群的遗传多样性明显低于大陆野生和人工饲养种群,近百年间一直维持着较低的遗传变异水平,表明该种群独立恢复或提高遗传多样性水平的可能性很低。
虽然丹顶鹤的保护遗传学和保护基因组学研究已经取得了一些重要的进展,但在许多方面开展的工作还很少,甚至还没有涉及,如大陆种群内部各区域分布种群的遗传多样性分化和各区域种群间的基因交流等。新的基因组学技术可为上述保护遗传学问题提供更加精准的研究结果,使之前难以开展的大规模系统性研究变成可能。为此,我们对目前迫切需要解决的丹顶鹤保护遗传学问题和未来的保护基因组学研究作出以下探讨和展望。
3.2 展望
3.2.1 关于岛屿种群的遗传多样性的恢复
丹顶鹤岛屿种群的遗传多样性很低,表明19世纪末至20世纪初,岛屿种群的极度减少导致了遗传多样性的严重丧失[26]。同样,由濒临灭绝的小种群恢复而来的美洲鹤,遗传多样性也一直维持在一个低水平状态[36]。在没有显著遗传结构分化的前提下,促进大陆与岛屿种群间的基因自然交流,或通过大陆个体或基因的人工引入方式提高岛屿种群的遗传多样性水平,应该是目前可取且可行的方案,但在实施之前,有必要对丹顶鹤物种的演化历史和大陆种群的遗传结构与遗传多样性进行全面了解和把握,依此制定具有遗传学依据的实施方案。
3.2.2 关于大陆种群内部地理性遗传变异和遗传多样性差异的研究
物种的分布区(繁殖地、越冬地和迁徙路径)是在长期演化过程中形成的,在分布区内又形成了各区域种群,这一区域性及其演化过程应在基因上有所体现[37]。对丹顶鹤岛屿种群的研究也显示:即使在很小的分布区域内,个体的基因交流也不是随机的,雄性个体的出生地留恋性比雌性更强,出生地留恋的栖息地选择可能导致小尺度距离隔离[31]。与岛屿种群相比,大陆种群有广泛的地域分布,通常依据繁殖地、迁徙路线和越冬地的明显差异分成东、西部2个区域种群,即使在各区域种群分布区内,也存在着各繁殖地间的地理隔离。因此,大陆种群内部或许也存在着地理性遗传变异和遗传多样性差异,或者某种特定的偏性扩散模式,对此尚不可知。在大陆种群,特别是我国境内繁殖越冬种群面临缩小和消失的情况下,认识与评价各区域种群的遗传结构特征和进化意义,了解区域种群间以及内部的基因交流方式,对深入认识和有效管理大陆种群、保护种群的进化潜力具有十分重要的意义,并且迫在眉睫[14]。
3.2.3 关于人工种群的谱系调查和遗传管理
目前,国内动物园和鹤类保护区饲养着一个数量超出国内繁殖越冬野生种群的人工种群[38]。对于濒危物种保护来说,人工饲养种群是一种重要的资源储备[13],但目前对这一人工种群的谱系和遗传特征了解得很少,基本上不清楚[30]。作为濒危物种保护的一个重要环节,谱系登记和基因信息库的建设必不可少,以保证放归自然个体的谱系和遗传信息是清楚的,对野生种群的遗传多样性影响是可预判的,从而实现对种群的遗传管理[32]。可以在已有的研究基础上,利用AFLP和微卫星等分子标记技术建立一个统一的标记体系,用于查明各人工饲养种群的谱系,建立基因信息库,进行统一监测和管理。
3.2.4 对保护基因组学研究的展望
目前,基因组学已从结构基因组学转向功能基因组学的后基因组时代,高通量测序技术也日渐成熟,正在成为特定鸟类研究领域的常规工具。低成本、短时间的高通量测序技术使大规模研究成为可能,如单核苷酸多态性(single nucleotide polymorphism,SNP)分析,相比多态性微卫星标记方法,在种群遗传交流、亲缘地理和区域适应,以及物种的演化历史等各研究层次上均具有更高的解析力[39],可用于丹顶鹤的相关研究。Xu等[40]用基因分型测序(genotyping-by-sequencing)方法开发了33个丹顶鹤的SNP标记,未来可将该标记体系用于基因信息库建设、丹顶鹤的遗传多样性保护和管理。此外,基因组学的高通量测序技术可以比保护遗传学方法更好地解析生物结构、功能与环境之间的协同演化关系,如能够提取环境中全部微生物DNA的宏基因组学(metagenomics)研究[34],未来也将成为深入研究丹顶鹤生理、行为、食性、疾病、生长发育和环境适应等课题的重要手段[35,41-42]。
猜你喜欢
格言·校园版(2022年10期)2022-05-22
思维与智慧·上半月(2022年4期)2022-04-08
四川文学(2020年10期)2020-02-06
中国中药杂志(2016年21期)2017-02-16
中国中药杂志(2016年21期)2017-02-16
学苑创造·A版(2016年10期)2016-11-19
上海医药(2016年13期)2016-07-26
儿童故事画报(2015年5期)2015-07-22
时代青年(上半月)(2009年6期)2009-07-31
幼儿智力世界(2006年2期)2006-03-08
