
基于选择信号分析揭示猪终端父本群体间性状趋同的关键基因
2022-08-23 02:39李望娇董文君李新云赵书红马云龙
中国畜牧兽医 2022年8期
李望娇,彭 夏,宋 徽,董文君,李新云,赵书红,马云龙
(华中农业大学,农业动物遗传育种与繁殖教育部重点实验室,武汉 430070)
中国地方猪种质资源十分丰富,约占世界总数的30%,且具有抗逆性强、耐粗饲和性成熟早等特点,然而多数品种存在脂肪含量高、饲料利用率低等缺点[1]。随着中国居民消费习惯的变化,瘦肉型猪的市场需求日益增长,在20世纪70年代至20世纪80年代,中国逐渐开始引入皮特兰猪和杜洛克猪等瘦肉型猪种在生猪繁育体系中作为终端父本使用[2-3]。在生猪生产中,皮特兰猪与杜洛克猪作为终端父本,其生长速度和饲料利用率长期受到高强度人工选择作用,且选育方向相似。人工选择作用在基因组上留下的特征痕迹称之为选择信号(selection signature)[4],针对商业猪种进行选择信号检测,可以筛选出与猪重要经济性状相关的分子标记和候选基因,为猪重要经济性状遗传改良提供一定的参考依据。目前,在猪上进行选择信号检测的研究报道了一系列重要的选择信号候选区域,在不同的繁殖力群体中ESR、AREG、PRLP等多个基因受到了不同程度的选择作用[5-6];BARX2基因作为肌肉生长、再生和维护的重要调节基因,在约克夏猪与长白猪中均受到选择[7];影响猪椎骨数及体长的多个基因(如NR6A1、PLAG1、LCORL)在欧洲商业猪和大多数欧洲地方猪群体中受到了强烈选择[8];ADAMTS12、SIM1和NOS1基因显示了藏猪自然选择的特征,可能与高海拔适应性相关,在具有带型毛色的中国地方猪EDNRB基因位点上发现了强烈的定向选择信号,表明EDNRB基因是中国地方猪白色带型的潜在候选基因[9]。
随着畜禽选择信号研究的深入,近年来群体间平行选择(parallel selection)信号的检测逐渐成为群体遗传学领域研究趋同遗传基础的重要方向[10]。Rehkamper等[11]研究发现,鸟类与哺乳动物大脑组织在结构和功能上具有大量相似性,主要表现于端脑中多模态信息融合能力方面,揭示了脊椎动物间发生了平行进化。此外,在人、三刺鱼及秀丽隐杆线虫等物种中也发现诸多关于平行选择的证据[12-14]。在畜禽相关研究方面,在蛋鸡和肉鸡群体中检测到多个与体型外貌、生产性能相关的平行选择候选基因,包括WWP1、BCDO2、TSHR、AGTR2和OPG等[[15-16];在杜洛克猪和大白猪群体间检测到67个平行选择区域,并挖掘出多个调节猪生长发育、体型大小等性状的候选基因[17]。
本研究拟通过对皮特兰猪、杜洛克猪2个商业猪种进行选择信号检测,揭示其作为终端父本时重要经济性状在人工选择作用下的基因组变化特征,利用检测到的群体间平行选择候选区域,结合生物信息学分析揭示表型趋同的功能候选基因,为进一步解析皮特兰猪、杜洛克猪2个品种重要经济性状的遗传基础,探讨表型趋同的遗传机制以及商业猪重要经济性状的遗传改良提供一定的参考依据。
1 材料与方法
1.1 材料
本研究共选取2 165头猪作为研究对象,样本分别来自中国南方2个生猪企业的4个群体,包括来自A猪场的376头皮特兰群体(PP)和451头杜洛克品系Ⅰ群体(DDⅠ),以及来自B猪场的841头杜洛克品系Ⅱ群体(DDⅡ)和497头杜洛克品系Ⅲ群体(DDⅢ)。对每头猪采集耳组织样,-20 ℃保存,用于提取全基因组DNA。
1.2 方法
1.2.1 质量控制及基因型填充 使用全基因组芯片进行基因分型,获得4个群体的50K SNP芯片数据。使用Plink软件[18]对50K SNP芯片数据进行质量控制,条件为:①SNP检出率(call rate)>90%;②样本的检出率>90%;③最小等位基因频率(minor allele frequency,MAF)为0.01。利用Beagle软件[19]对缺失的基因型进行填充,对填充后的群体计算个体间的亲缘关系系数(PI_HAT),并质控掉亲缘关系较近(PI_HAT>0.5)的个体。
1.2.2 选择信号检测 利用综合单倍型评分(iHS)[20]与等位基因频率差(△AF)2种方法分别进行群体内及群体间选择信号检测,△AF定义为2个群体间等位基因频率差的绝对值。研究利用R语言中的REHH包[21]及Plink软件,按照100 kb窗口、50 kb步长在全基因组范围内划分窗口进行选择信号检测,对窗口统计量平均值进行秩排序,取其前5%的窗口作为选择信号显著候选区域。
1.2.3 平行选择信号候选区域鉴定 结合iHS和△AF进行皮特兰猪和杜洛克猪群体间的平行选择信号检测。4个群体两两组合,以PP、DDⅠ群体为例,利用bedtools软件将iHS、△AF 2种统计量所对应的选择信号候选区域按照窗口左右200 kb的范围进行合并。提取iHSPP和iHSDDⅠ重叠区域,再提取iHSPP_iHSDDⅠ与△AFPP-DDⅠ的重叠区域,所得区域即定义为PP和DDⅠ 2个群体间的平行选择候选区域。
1.2.4 选择信号候选区域注释 基于定位到的平行选择候选区域,利用Ensembl[22]平台的BioMart(http:∥www.biomart.org)软件进行基因挖掘;使用DAVID[23]开展基因集进行富集分析,显著富集条目定义为P<0.05;基于Ensembl注释的基因,通过NCBI数据库对其进行功能注释,挖掘与猪繁殖、生长和肉质等性状相关的候选基因。
2 结 果
2.1 数据质量控制
皮特兰猪和杜洛克猪4个群体的SNP数据质量控制结果见表1。由表1可知,质控后DDⅠ群体获得的SNPs数目最多(44 056个),DDⅢ群体获得的SNP数目最少(37 323个)。2 165个样本中通过样本检出率的个体共2 140个,对其经亲缘关系系数过滤(PI_HAT>0.5)后,最终得到1 003个亲缘关系较远个体。
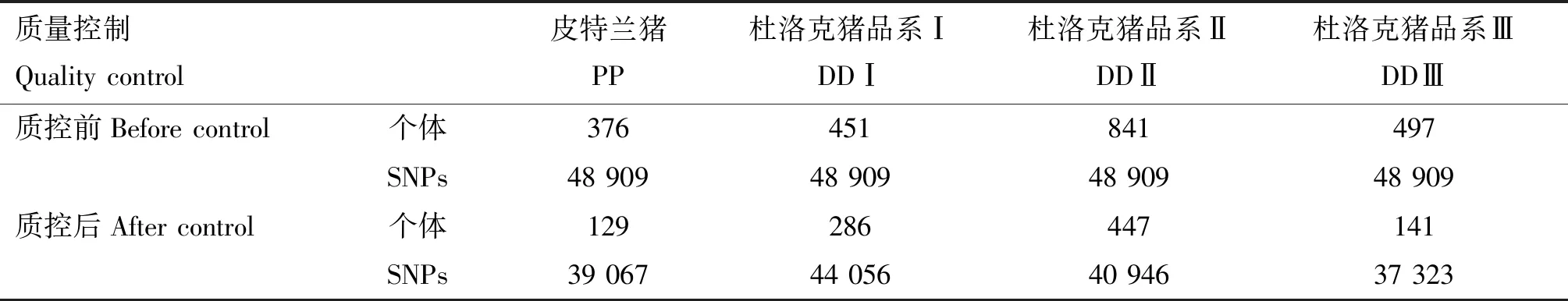
表1 皮特兰猪和杜洛克猪群体质量控制结果
2.2 群体内选择信号候选区域检测
基于iHS方法群体内选择信号检测发现,在PP群体的25 091个iHS统计量中有1 254个统计量达到显著水平,长度为119.59 Mb,约占全基因组总长的4.94%;在DDⅠ群体的27 195个iHS统计量中有1 359个统计量达到显著水平,长度为129.60 Mb,约占全基因组总长的5.35%;在DDⅡ群体的27 582个iHS统计量中有1 379个统计量达到显著水平,长度为131.51 Mb,约占全基因组总长的5.43%;在DDⅢ群体的22 398个iHS统计量中有1 120个统计量达到显著水平,长度为106.81 Mb,约占全基因组总长的4.41%(图1)。
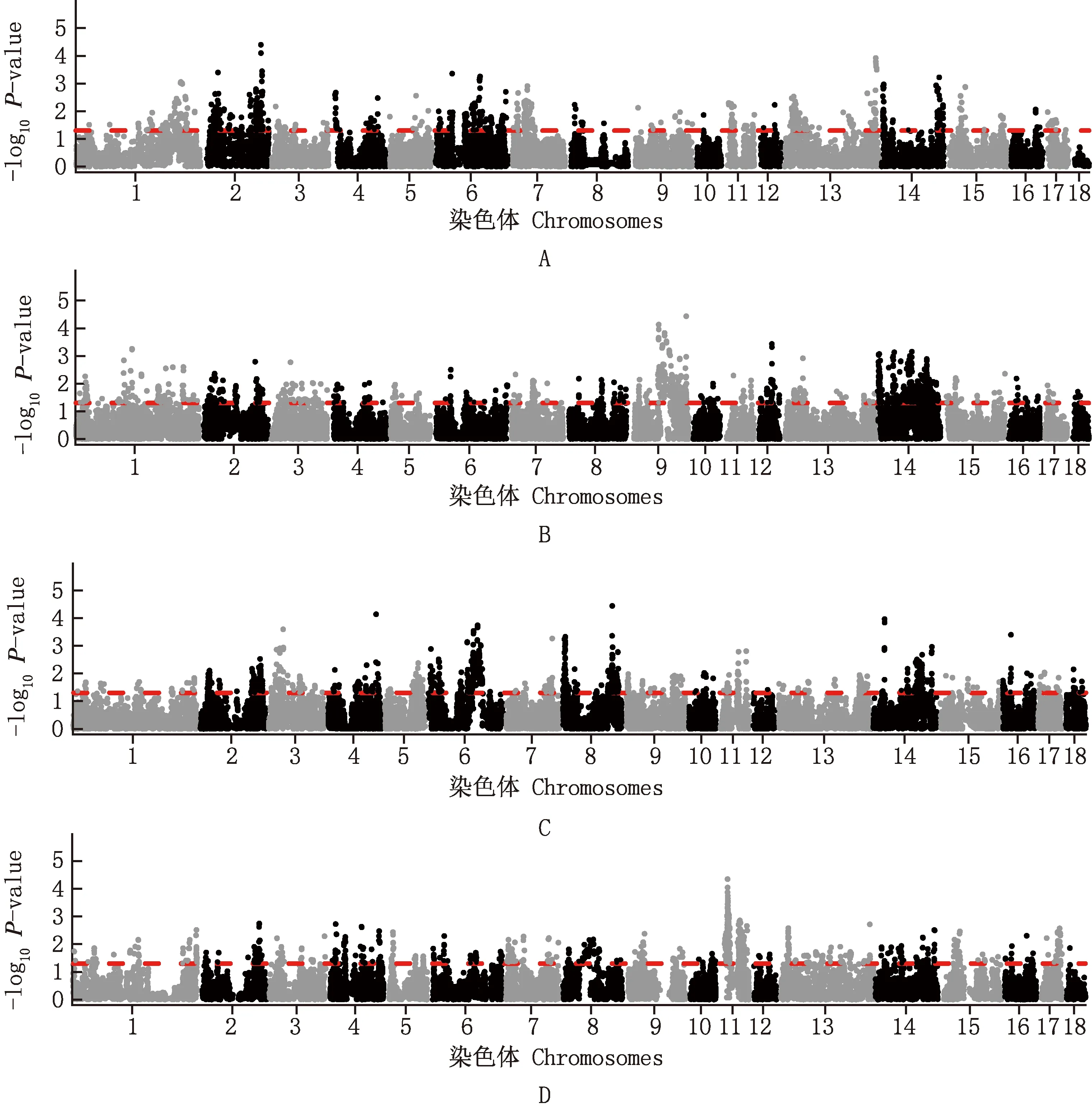
A~D,PP、DDⅠ、DDⅡ、DDⅢ群体A-D,PP,DDⅠ,DDⅡ and DDⅢ populations,respectively图1 皮特兰猪和不同品系杜洛克猪群体内信号检测结果Fig.1 Ingroup selection signature detection results of Pietrain pigs and different strains of Duroc pigs
2.3 群体间选择信号候选区域鉴定
由表2可知,基于△AF方法群体间选择信号显著候选区域共有9 579个,总长度为913.50 Mb,其中,PP-DDⅠ、PP-DDⅡ、PP-DDⅢ、DDⅠ-DDⅡ、DDⅠ-DDⅢ、DDⅡ-DDⅢ群体对分别检测到1 648、1 541、1 467、1 689、1 616和1 618个基因组候选区域,分别约占基因组总长的6.49%、6.07%、5.78%、6.65%、6.38%和6.38%。
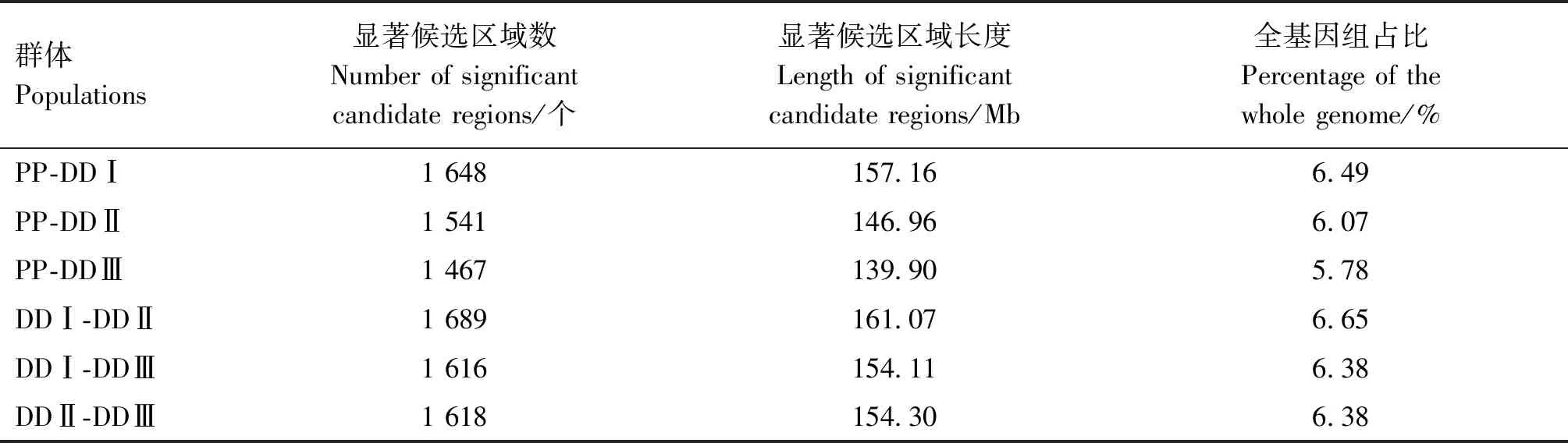
表2 皮特兰猪和不同品系杜洛克猪群体间选择信号检测结果
2.4 平行选择候选区域鉴定
将iHS统计量按照窗口左右各200 kb的范围进行合并处理,共检测出1 591个群体内选择信号候选区域,其中PP群体390个,长度为99.75 Mb;DDⅠ群体460个,长度为110.82 Mb;DDⅡ群体400个,长度为106.86 Mb;DDⅢ群体341个,长度为86.93 Mb(表3)。
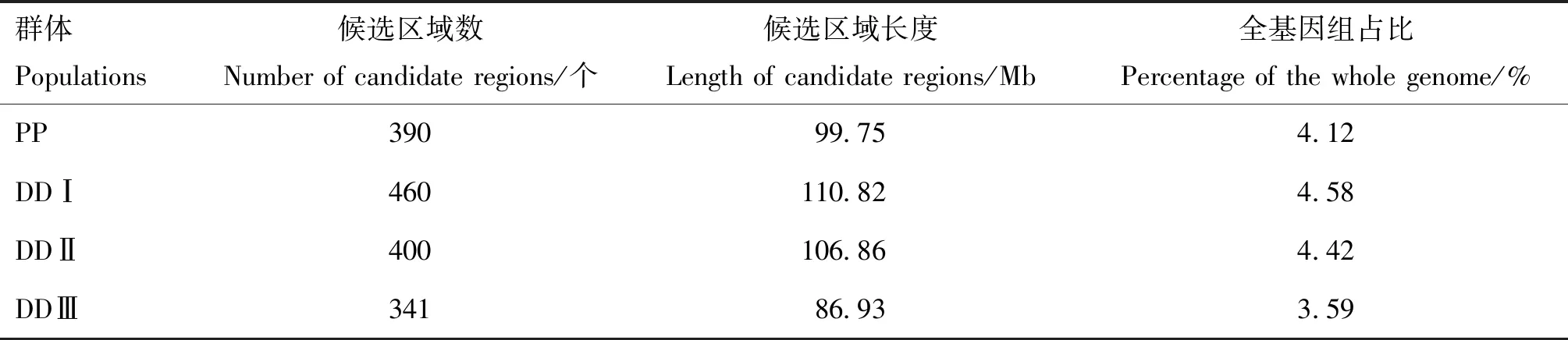
表3 群体内选择信号候选区域
将△AF统计量按照窗口左右各200 kb的范围进行合并处理,共发现3 466个群体间选择信号候选区域,其中PP-DDⅠ群体间有851个候选区域,长度为136.14 Mb;PP-DDⅡ群体间有795个,长度为127.28 Mb;PP-DDⅢ群体间有733个,长度为121.74 Mb;DDⅠ-DDⅡ群体间有798个,长度为137.66 Mb;DDⅠ-DDⅢ群体间有744个,长度为136.42 Mb;DDⅡ-DDⅢ群体间的候选区域有678个,长度为127.36 Mb(表4)。

表4 群体间选择信号候选区域
利用bedtools软件在4个群体中共挖掘到52个平行选择候选区域(54个区域中包含2个重复区域),其长度约为4.67 Mb,值得注意的是,在DDⅠ-DDⅢ群体间检测到的5个平行选择候选区域中,位于11及14号染色体的2个区域分别在PP-DDⅢ、DDⅠ-DDⅡ群体间被检测到,长度均约为0.095 Mb(图2)。
2.5 平行选择信号候选区域基因功能注释
利用BioMart软件挖掘平行选择区域的基因,注释到88个平行选择候选基因。DAVID数据库中GO功能富集结果显示,DDⅠ-DDⅡ群体间的平行候选基因LIPA、LIPM参与脂质分解过程;PP-DD Ⅰ群体间平行选择候选区域功能注释结果见表5,挖掘出多个与生殖系统发育(HSD17B4、FAM170A、AXDND1)、调节脂质代谢和脂肪酸代谢(HSD17B4、SOAT1、COMMD6、CDCP1、ACSL5)、参与内质网修饰(ZDHHC6)等生物过程相关的候选基因;PP-DDⅡ群体间注释到4个平行选择候选基因(GALNT18、FER、LDLRAD4、PLCB4),参与调节机体生长发育及细胞增殖等;PP-DDⅢ群体间挖掘出多个与脂肪沉积相关的基因(KLHDC3、MEA1、GPC6);DDⅠ-DDⅡ群体间挖掘到33个平行选择候选基因;DDⅠ-DDⅢ群体间挖掘到8个候选基因;DDⅡ-DDⅢ群体间挖掘到11个候选基因,主要调控猪的胴体性状(ACOXL、ZNF638、GCP6、SUPV3L1、LIPN、LIPM、ACTA2、LIPA、FFAR4、PDE6C、SMARCD1、SNCG)以及繁殖性状(TMEM132D、VPS26A、RBP4、GDNF、SPACA3、BMPR1A)。

A~F,PP-DDⅠ、PP-DDⅡ、PP-DDⅢ、DDⅠ-DDⅡ、DDⅠ-DDⅢ和DDⅡ-DDⅢA-F,PP-DDⅠ,PP-DDⅡ,PP-DDⅢ,DDⅠ-DDⅡ,DDⅠ-DDⅢ and DDⅡ-DDⅢ,respectively图2 群体间平行选择候选区域数目Fig.2 Number of candidate regions for intergroup parallel selection
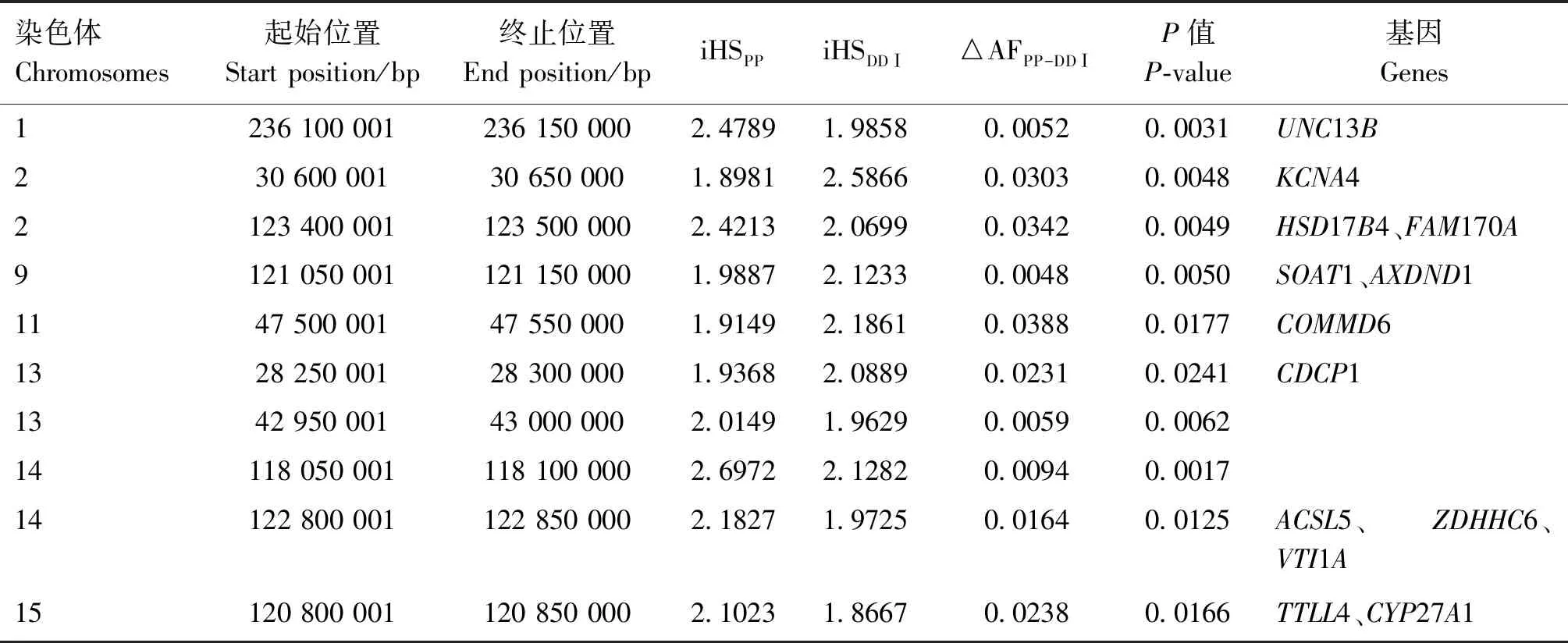
表5 PP-DDⅠ群体间平行选择候选区域和候选基因
3 讨 论
为研究皮特兰猪和杜洛克猪作为终端父本在人工选择作用下性状趋同的基因组变化特征,本研究基于来自A猪场的PP、DDⅠ群体和来自B猪场的DDⅡ、DDⅢ群体的50K SNP芯片数据进行选择信号检测,通过整合iHS和△AF在PP、DDⅠ、DDⅡ和DDⅢ 4个群体中的选择信号检测结果,挑选至少在每2个群体中都存在的重叠区域,共得到52个性状趋同的基因组区域,其中,皮特兰猪与3个杜洛克猪群体间共检测到21个候选区域,而3个杜洛克猪群体间共筛选出33个平行选择的基因组候选区域,表明由于品种内不同品系间具有更高的遗传背景,因此性状趋同的基因组特征一致性更高。然而,PP与DDⅠ群体间共检测到10个候选区域,表明不同品种在相似育种方向的驱动下仍然能够形成较高的基因组特征相似。此外,DDⅠ和DDⅡ群体间共筛选出18个候选区域,与两群体均属于美系杜洛克猪密切相关;尽管DDⅠ和DDⅢ群体均属于杜洛克猪品种,但DDⅢ群体为丹系杜洛克猪且来源于B猪场,在DDⅠ和DDⅢ群体间仅检测到5个性状趋同候选区域,表明不同品系间杜洛克猪群体的人工选择方向可能存在一定的差异。
本研究对52个性状趋同区域进行基因注释,共挖掘出88个受选择的候选基因,主要参与调节脂肪沉积、生长发育、繁殖、免疫等性状。GO功能富集到脂质分解过程,涉及LIPA、LIPM2个候选基因。研究表明,小鼠酸性脂肪酶基因受组织mRNA表达差异的影响,LIPA基因表现为广泛的组织表达,而LIPM基因表达受限,仅在表皮组织中表达[24-25];ZNF638、FFAR4、PDE6C基因共同参与调节脂肪细胞的发育和分化[26-28];ACOXL、ACSL5、CDCP1、COMMD6、HSD17B4、SOAT1、PPP2R5D、KLHDC3和ACTA2基因通过调节脂质代谢影响猪的脂肪沉积,进而影响猪的肌内脂肪含量[29-36];GNMT基因通过利用和分配营养素影响肉质性状[37];PTPRM基因可调节肌肉生长发育[38];SLC16A12基因参与肌酸的转运,从而调节机体肌肉的生长发育[39]。猪的肉质与肌内脂肪含量和肌肉密切相关,因此上述基因在4个群体中受到趋同选择。猪的生长速度与瘦肉率长期以来一直受到养殖户和企业的关注,SUPV3L1基因是一种重要的发育调控基因,可调节机体的生长发育,SUPV3L1基因的表达始于囊胚期,在所有胎儿组织和细胞类型中广泛表达,成年小鼠中敲除SUPV3L1基因会导致早衰,包括肌肉、脂肪组织的损失和严重的皮肤异常[40];PLCB4基因能调节猪的生长和骨架,从而影响猪体重和身体构象性状[41];CUL7基因对猪四肢和蹄筋坚固性起着重要作用[42]。
皮特兰猪和杜洛克猪作为终端父本,尽管其在总产仔数性状上选择强度始终较低,然而精液品质始终是重点关注的性状类型。研究发现,FAM170A基因可能通过调节其他基因的表达来介导其对精子细胞头形和精子形成的影响[43];AXDND1基因仅在人和小鼠的圆形和细长的精子细胞中表达,可能通过调节精子管的动力学、精子头的形状和精子鞭毛的组装对精子发生和雄性生育起着重要作用[44];KLHDC3与MEA1基因在人与小鼠中的同源基因均被证明与精子生成相关[45];GDNF基因被证明与精原干细胞的维持密切相关[46];SPACA3基因与精子顶体相关,直接影响精子和卵子结合过程,从而影响母畜受胎率[47];SNCAIP和VPS26A基因调节胚胎发育,可间接影响产仔数;RBP4基因可影响猪繁殖性状,相较于纯合子个体,在杂合子个体中表现出较高的产仔数[48]。因此,在生产过程中,终端父本的繁殖性状同第一母本一样一直受到长时间、高强度的人工选择,这些基因的发现可为繁殖性状的遗传标记提供一定参考。
此外,在集约化饲养条件下瘦肉型猪种的抗性实质上长期处于高强度选择压力之下,本研究发现,在猪的选育过程中CAMK4、PTCRA和IFIT5基因受到强烈的选择作用,其中,CAMK4基因是一种多功能丝氨酸/苏氨酸激酶,其通过激活包括T细胞和抗原呈递细胞的各种免疫细胞中的转录因子来调节基因表达[49];PTCRA基因通过编码前T细胞抗原受体α,以在分化的不同阶段调节早期T细胞发育[50];IFIT5基因是先天免疫反应的重要增强因子,在IFIT5基因存在的情况下,宿主细胞的抗病毒反应显著增加[51]。
4 结 论
皮特兰猪和杜洛克猪各群体内选择信号显著候选区域共有5 112个,群体间的选择信号候选区域共有9 579个,在群体之间存在平行选择的候选区域共52个,包含88个受到平行选择的基因;基因富集及功能注释发现这些基因主要与猪的肉质、生长发育及繁殖性状相关,表明高强度的人工选择作用是造成繁殖力、胴体、肉质等重要经济性状表型趋同的重要因素。
猜你喜欢
水产养殖(2022年8期)2022-09-21
军事文摘(2022年16期)2022-08-24
中学生数理化·高一版(2022年4期)2022-05-09
新高考·高一数学(2022年3期)2022-04-28
中国典型病例大全(2022年7期)2022-04-22
思维与智慧·上半月(2022年4期)2022-04-08
新高考·高一物理(2017年7期)2018-03-06
农民致富之友(2017年19期)2017-10-21
中学生物学(2008年11期)2008-12-02
作文与考试·高中版(2008年11期)2008-11-21
