
猪繁殖与呼吸综合征病毒RFLP 1-4-4 L1C株的分离鉴定及其遗传演化分析
2022-08-24 12:53田笑笑黄昕怡夏大松王海伟田志军蔡雪辉安同庆
中国预防兽医学报 2022年6期
田笑笑,黄昕怡,夏大松,王海伟,田志军,蔡雪辉,安同庆
(中国农业科学院哈尔滨兽医研究所兽医生物技术国家重点实验室,黑龙江 哈尔滨 150069)
猪繁殖与呼吸综合征(Porcine reproductive and re-spiratory syndrome,PRRS)是由PRRS 病毒(PRRS vi-rus,PRRSV)引起的,以妊娠母猪繁殖障碍和各生长阶段猪呼吸道症状为主的严重影响世界养猪业的传染病。PRRSV 属于套式病毒目、动脉炎病毒科成员,可分为两个属,Betaarterivirus suid 1(欧洲型PRRSV)和Betaarterivirus suid 2(北美洲型PRRSV)[1]。我国于1996 年首次发现PRRS[2],随后在我国各主要养猪省份均有该病的发生。2006 年以来,我国相继出现了高致病性PRRSV(HP-PRRSV)变异株[3-4]、类NADC30 株[5-6]、类NADC34 株[7-8],给我国养猪业造成了严重的经济损失。
PRRSV 遗传多样性复杂,病毒基因组中的ORF5和NSP2 是两个高变异区[9]。为了区分PRRSV 野毒株与VR-2332 株的传代致弱疫苗株,美国研究人员建立了PRRSV ORF5 基因的限制性片段多态性(Restric-tion fragment length polymorphism,RFLP)分类方法,该方法根据MluI、Hinc II 和SacII 酶切ORF5 基因后的DNA 片段长度和数量对PRRSV 分类和命名[10]。该方法是使用3 个数字分别代表ORF5 基因3 种酶切位点(MluI、Hinc II 和SacII)的数量和位置。例如:1-4-4 模式代表1(MluI=0)、4(Hinc II=nt88、nt219)和4(SacII=nt24、nt555)。RFLP 分类方法在美国养猪从业者使用较多,2007 年~2019 年美国有228 种RFLP模式的PRRSV,数量较多的RFLP 模式依次是RFLP 2-5-2(VR-2332 株传代致弱疫苗株为该模式)、1-7-4、1-4-4 和1-8-4,其中RFLP 1-7-4 是近年来的优势流行模式[12]。另外一种分类方法是根据ORF5 基因序列的系统发生树和序列之间的同源性,将北美洲型PRRSV 分为9 个分支谱系,L1~L9[11],其中,L1 可进一步划分为L1A~L1H。
2020 年10 月以来,美国明尼苏达州和衣阿华州突然出现大量的妊娠母猪流产、产木乃伊胎,伴有成年母猪的死亡;断奶仔猪高热、死亡率高达15%~30%;育肥猪生长缓慢并有呼吸道症状[13-14],该病迅速扩散周边多个州,现有疫苗对其保护效果不理想,经研究发现病原是PRRSV RFLP 1-4-4 lineage 1C(L1C)变异株[13]。我国尚无RFLP 1-4-4 L1C 变异株的相关报道。鉴于我国有从美国引种和进口冷冻猪肉的贸易,以及美国的NADC30 株、NADC34 株已经传入我国[5,15],因此在我国开展RFLP 1-4-4 L1C 的遗传演化分析非常必要。本研究对我国2016 年~2021 年从黑龙江、广东和山东等15 个省份多个猪场采集的共219 份临床发病猪样品采用PRRSV ORF5 基因RTPCR 方法检测,并对PRRSV 阳性样品ORF5 基因测序用于RFLP 模式分析,将PRRSV ORF5 基因RFLP 1-4-4 的阳性病料样品猪肺泡巨噬细胞(PAM)分离到一株PRRSV,采用PCR 扩增分离病毒的全基因组并测序拼接;分析其同源性、遗传演化及其NSP2 氨基酸序列特征和全基因组的重组事件。同时还分析了1996 年~2021 年我国PRRSV RFLP 1-4-4 株在时间和地理上的分布,比较我国与其他国家PRRSV 的RFLP 模式。为了解我国RFLP 1-4-4 模式PRRSV 的流行情况提供了重要参考数据。
1 材料与方法
1.1 主要实验材料参考郭宝清等的方法[2]制备PAM,用于制备PAM 的4 周龄SPF 猪购自中国农业科学院哈尔滨兽医研究所实验动物基地;Marc-145细胞、PRRSV HuN4 株由本实验室保存;PRRSV M蛋白单克隆抗体(MAb)3F7 由哈尔滨兽医研究所王倩博士惠赠;FITC 标记的羊抗鼠IgG 购自Sigma-Aldrich公司;病毒核酸提取试剂盒QIAamp Viral RNA Mini Kit 购 自QIAGEN 公 司;Recombinant RNase Inhibitor、Reverse Transcriptase M-MLV 和PremixTaq购自TaKa-Ra 公司。
1.2 临床样品的PRRSV 检测及其ORF5 基因的测序2016 年~2021 年间,从黑龙江、广东和山东等15 个省多个猪场共采集219 份临床发病猪的病料样品,取组织研磨后的上清液提取病毒RNA,反转录为cDNA 作为模板,按照本实验室建立的PRRSV ORF5 基因RT-PCR 方法检测,引物序列和反应条件均参考文献[16],扩增产物由吉林库美生物科技有限公司测序。
1.3 PRRSV ORF5 基因的RFLP 模式分类根据Wesley 等建立的PRRSV ORF5 基因RFLP 分类方法[10],对PRRSV 阳性样品的ORF5 基因序列进行分类。
1.4 PRRSV 的分离与鉴定取PRRSV ORF5 基因为RFLP 1-4-4 的阳性病料样品组织研磨液以8 000 r/min离心5 min 后取上清,用0.22 μm 滤器过滤除菌后接种PAM,5% CO237 ℃培养72 h 后收集上清液,继续在PAM 中连续传3 代后,取病毒液接种Marc-145 细胞,72 h 后收集上清液。对PAM 连续传3 代后的上清液及Marc-145 细胞的上清液提取病毒RNA,反转录为cDNA 作为模板,按照1.2 RT-PCR 方法扩增分离病毒的ORF5 基因;将上述收获病毒上清液后的PAM 和Marc-145 细胞分别经无水乙醇固定,经PBST洗3 次后,以PRRSV M 蛋白MAb 3F7 为一抗,FITC标记羊抗鼠IgG(1:200)为二抗,室温孵育1 h,以不接种病毒的相应细胞作为阴性对照,以接种HuN4 病毒的细胞作为阳性对照,经间接免疫荧光试验(IFA)鉴定。
1.5 分离病毒的全基因组测序及序列分析提取分离病毒的RNA 反转录为cDNA 作为模板,按照Zhou等的引物和方法对其基因组分成8 个重叠的片段进行PCR 扩增[3],PCR 产物分别克隆至pMD18-T 载体中,并经PCR 鉴定为阳性的质粒由吉林库美生物科技有限公司测序。序列经DNAStar 软件包中的Seq-Man拼接。利用MegAlign 软件分析分离株的全基因组序列和各ORF 基因序列分别与PRRSV 代表株(CH-1a、JXA1、NADC30、NADC34、GM2 和VR-2332)及美国RFLP 1-4-4 L1C 变异株的相应基因序列的同源性;利用MEGA6.0 软件对分离株、美国新发1-4-4 L1C 变异株及PRRSV 各分支参考株(L1A~L1H、L3、L5 和L8)的ORF5 基因序列绘制系统发生树(ML 法),分析分离病毒的遗传进化关系;参考Yu 等的方法[17],利用MegAlign 对分离病毒NSP2 的氨基酸序列比对分析;利用RDP4 重组分析软件对分离株全基因组序列进行重组分析。
1.6 1996 年~2021 年我国PRRSV RFLP 1-4-4 的时间和地理分布为了确定PRRSV RFLP 1-4-4 L1C 在我国的流行情况,本研究将PRRSV 阳性临床样品中测序获得的41 条ORF5 基因序列,连同GenBank 中1996 年~2021 年我国3 125条北美洲型PRRSV ORF5基因序列,共计3 166条,参照上述的PRRSV RFLP 1-4-4模式和L1C 分支谱系划分依据分类[11]。根据分类结果、病毒株的采集时间和省份,对我国RFLP 1-4-4 L1C株进行了时间和地理分布的分析。
1.7 我国与其他国家PRRSV RFLP 模式的比较分析为了进一步了解近年来我国PRRSV 流行株的各种RFLP 模式,以及和同时期国外流行株RFLP 模式的差异,本研究下载了2016 年~2021 年GenBank 中所有北美洲型PRRSV ORF5 基因序列(共计1 918条),分别来自美国、中国、加拿大等7 个国家。按照PRRSV RFLP 分类方法分类[10],分析比较2016 年以来不同国家之间PRRSV RFLP 的模式特点。
2 结 果
2.1 临床样品中PRRSV 的检测结果对15 个省的猪场采集的219 份临床样品经PRRSV ORF5 基因RTPCR 检测,结果显示,检测出41份PRRSV 阳性样品,占比18.72%,对阳性样品扩增产物测序并采用RFLP分型结果显示,其中有1 份来自黑龙江省的PRRSV 阳性临床样品的ORF5基因为RFLP 1-4-4模式。
2.2 PRRSV 的分离与鉴定结果将ORF5 基因为RFLP 1-4-4 模式的PRRSV 阳性病料样品研磨液接种PAM,48 h 后部分细胞出现破碎和脱落。在PAM 中传至3 代后,接种Marc-145 细胞。72 h 后收获两种细胞的上清液分别经RT-PCR 检测,结果显示仅PAM 上清液中检测到PRRSV 目的基因(图1A);将上述PAM、Marc-145 细胞固定后,利用PRRSV M 蛋白MAb 进行IFA 鉴定。结果显示,仅PAM 中有特异性绿色荧光,接种病毒的Marc-145 细胞(结果未展示)和未接种病毒的对照细胞均无该绿色荧光(图1B)。上述结果表明,本研究从上述PRRSV 阳性病料样品中分离到了一株PRRSV RFLP 1-4-4 株,且该PRRSV能够在PAM 中增殖,不能在Marc-145 细胞中增殖,将该分离株命名为HLJ-80。
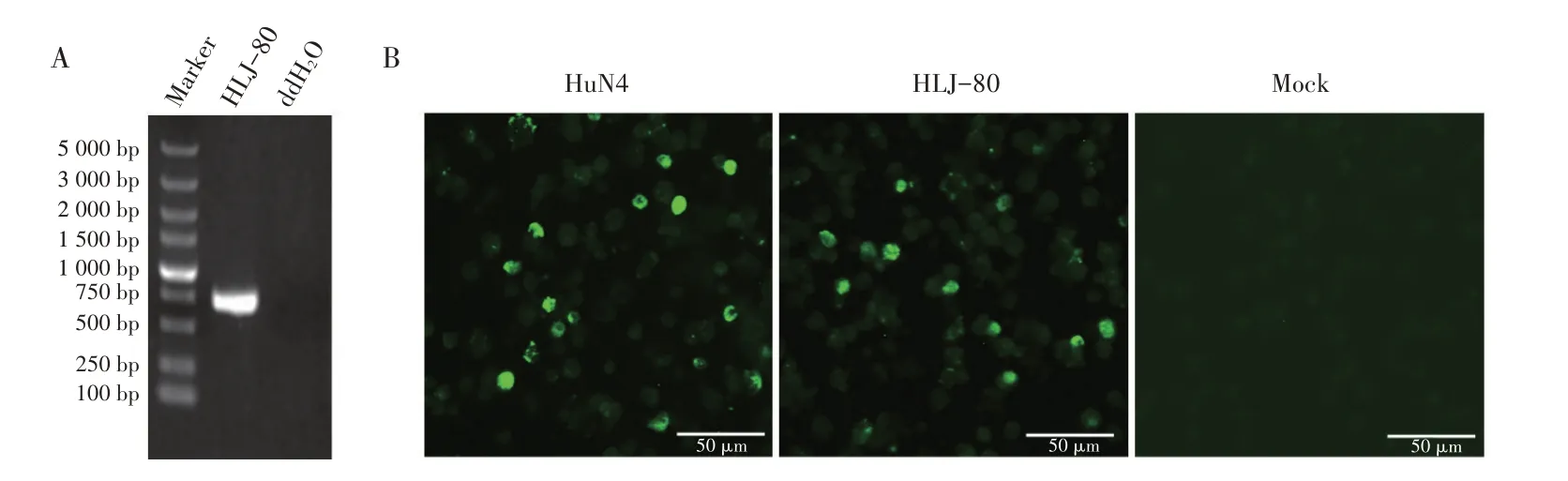
图1 分离病毒的RT-PCR(A)及IFA(B,在PAM中分离)的鉴定结果Fig.1 Identification results of RT-PCR(A)and IFA(B,in PAM cells)of isolated viruses
2.3 HLJ-80 株全基因组序列的同源性分析以分离病毒HLJ-80 株的cDNA 为模板,经PCR 分段扩增其基因组并测序,利用SeqMan 软件对测序结果拼接后获得了该病毒的全基因组序列。结果显示,HLJ-80株基因组全长为15 013 nt(不含3'末端的poly A),全基因组序列提交GenBank获得登录号为MN046222。HLJ-80 株 与PRRSV 代 表 株(CH-1a、 JXA1、 NADC30、NADC34、GM2 和VR-2332)的全基因组序列同源性分析结果显示,HLJ-80 株与NADC30 株的同源性最高,为93.3%,与其他代表株的同源性均低于90%。各ORF 基因序列的同源性分析结果显示,除ORF2 基因与VR-2332 株ORF2 基因同源性最高(97.5%)外,其余各基因均与NADC30 株同源性最高(90.6%~99.3%)。NSP2 氨基酸序列的比对分析结果显示,与VR-2332 株相比,HLJ-80 株NSP2 存在131 个氨基酸(111+1+19)的不连续缺失,缺失位置与NADC30 株一致。上述结果表明,HLJ-80 株与NADC30 株同源性较高,且其NSP2 氨基酸缺失特征与NADC30 株一致。
2.4 HLJ-80 株的重组特征分析以L1~L9 分支的PRRSV 代表株(分别为NADC30、XW008、MD001、EDRD-1、VR-2332、 P129、JXA1 和MN30100)为亲本株,利用RDP4 软件分析HLJ-80 株基因组中可能存在的重组事件,结果显示,HLJ-80 株是以NADC30 株为主要亲本、VR2332 株为次要亲本的重组病毒株。根据初步分析的重组断点,将HLJ-80 株基因组的主要亲本片段、次要亲本片段分别利用BLAST 检索与该株病素相应基因序列同源性较高的潜在亲本株。结果显示,HLJ-80 株主要亲本片段与PRRSV 2014-81 株同源性最高,次要亲本片段与PRRSV SD53-1603 株同源性最高,二者的分离时间均早于HLJ-80 株。随后以PRRSV 2014-81 株和SD53-1603 株为参考序列重新采用RDP4 进行重组分析,结果显示,重组断点有4 个,将HLJ-80 株分为两个明显的重组区域分别为nt1 601~nt3 101和nt9 001~nt14 921(图2)。表明HLJ-80株有可能与PRRSV 2014-81 株、SD53-1603 株的高度同源的病毒株重组而来。
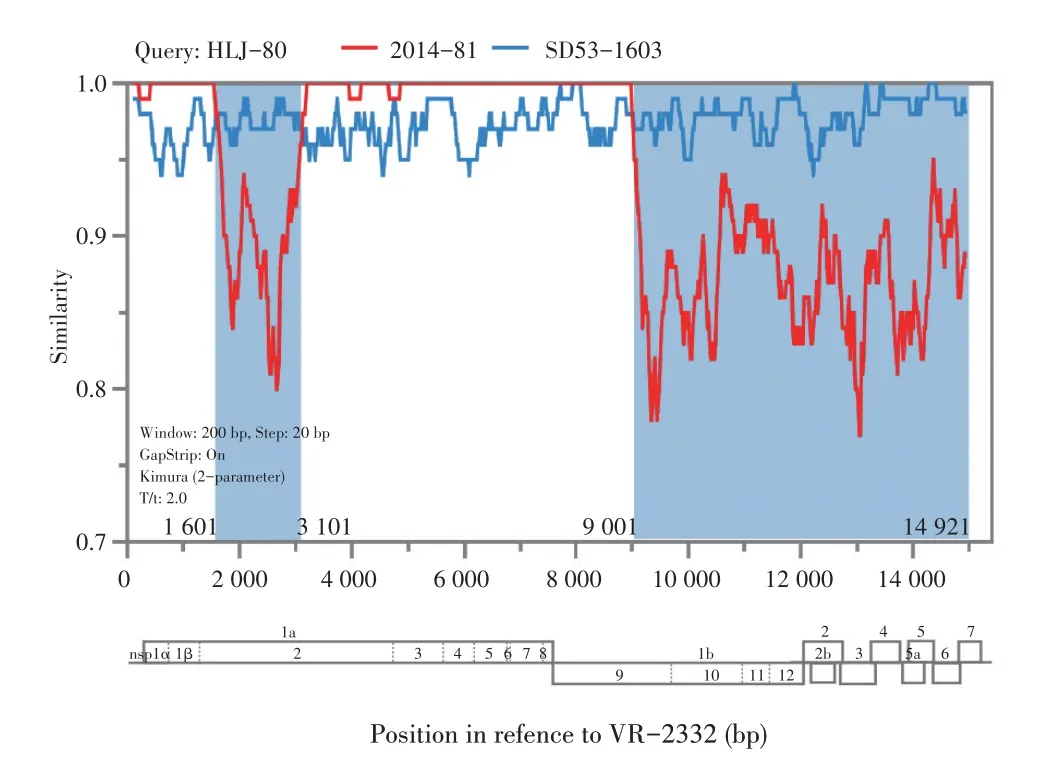
图2 PRRSV HLJ-80株的重组分析Fig.2 Recombination analysis of PRRSV HLJ-80 strain
2.5 HLJ-80 株的遗传演化分析利用MEGA6.0 软件对HLJ-80 株、美国新发1-4-4 L1C 变异株和各分支代表株(L1A~L1H、L3、L5 和L8)的ORF5 基因构建遗传进化树。结果显示,HLJ-80 株属于L1C 分支(图3),即HLJ-80 株属于RFLP 1-4-4 L1C 株。HLJ-80 株与美国RFLP 1-4-4 L1C 变异株虽然同属于L1C分支,但处于不同的分支(图3);全基因组同源性分析结果显示,HLJ-80 株与美国1-4-4 L1C 变异株同源性为93.4%,表明HLJ-80 株与美国1-4-4 L1C 变异株不是同一株病毒。
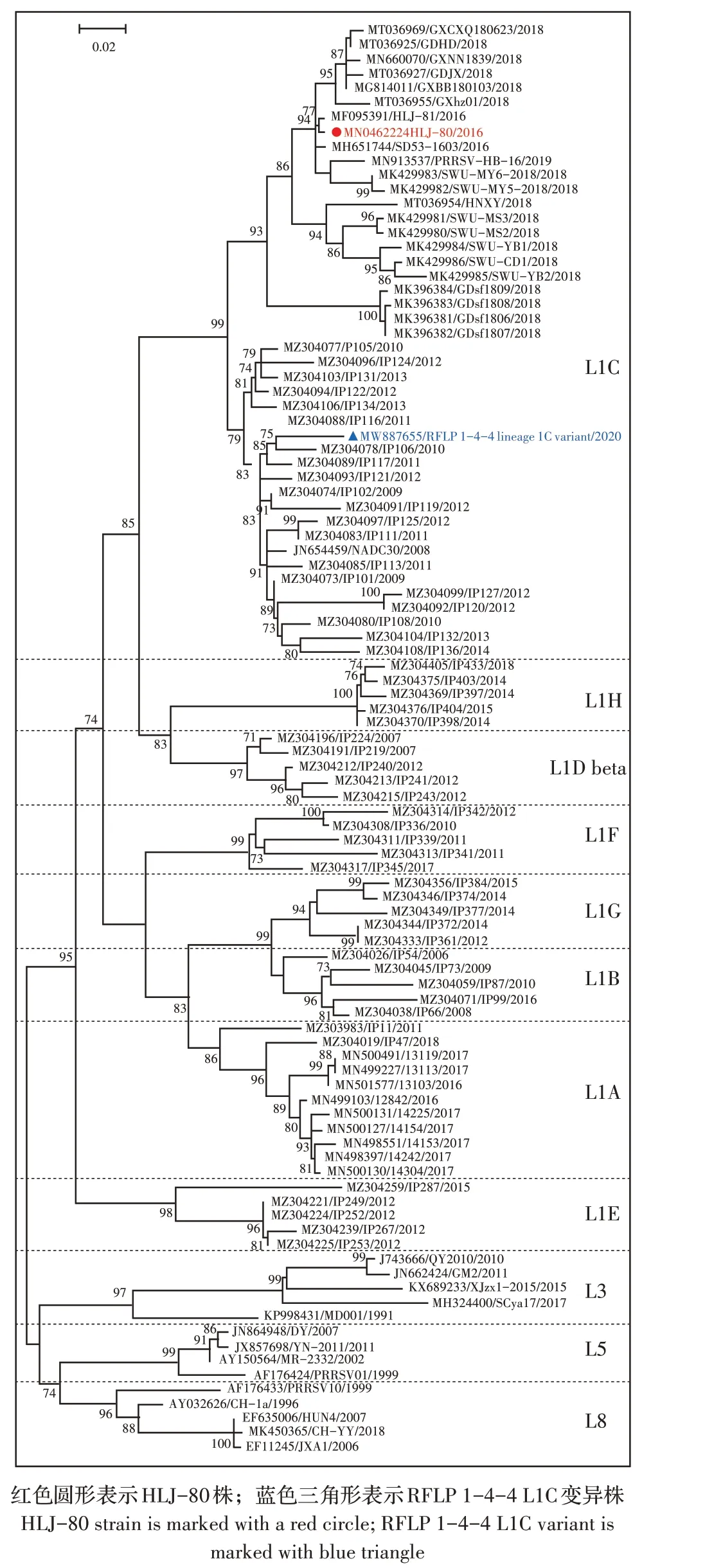
图3 基于PRRSV ORF5基因构建的系统进化树Fig.3 Phylogenetic tree of PRRSV based on ORF5 gene sequences
2.6 1996 年~2021 年我国PRRSV RFLP 1-4-4 株的时间和地理分布1996 年~2021 年,GenBank 中共有3 125 条PRRSV ORF5 基因序列来自我国,加上本研究测序获得的41 条,共计3 166 条ORF5 基因序列。根据PRRSV RFLP 分类依据分类和RFLP 1-4-4 株的统计分析。结果显示,我国共有260 条ORF5 基因属于(8.2%)RFLP 1-4-4 模式株。将该260 条RFLP 1-4-4 模式株ORF5 基因序列进行分支谱系划分,结果显示其中217 条属于L1C 分支(217/260,83.46%),其余为L8和L1A分支(图4A)。在时间上,PRRSV RFLP 1-4-4 模 式 在2003 年 就 已 存 在(HZ-X 株,GenBank AY450301),属于L8 分支;在2006 年~2009 年间该模式的PRRSV均属于L8分支,2010年~2013年间则为L8和L1A分支共存;2014年以后RFLP 1-4-4株的数量迅速增多,且绝大多数PRRSV 属于L1C 分支。在地理分布上,RFLP 1-4-4 L1C 分支主要分布在河南、广东和山东,分别占总数的31.15%、10%和5.38%,各省该类病毒株数量依次为河南(n=81)、广东(n=26)、山东(n=14)、福建(n=14)、江苏(n=13)、四川(n=11)、黑龙江(n=10)、新疆(n=7)、河北(n=6)、江西(n=6)、安徽(n=5)、广西(n=5)、山西(n=5)、浙江(n=5)、北京(n=3)、湖北(n=2)、贵州(n=1)、吉林(n=1)、上海(n=1)和中国台湾(n=1)(图4B)。表明PRRSV RFLP 1-4-4 L1C分支早已出现在我国多个省份。
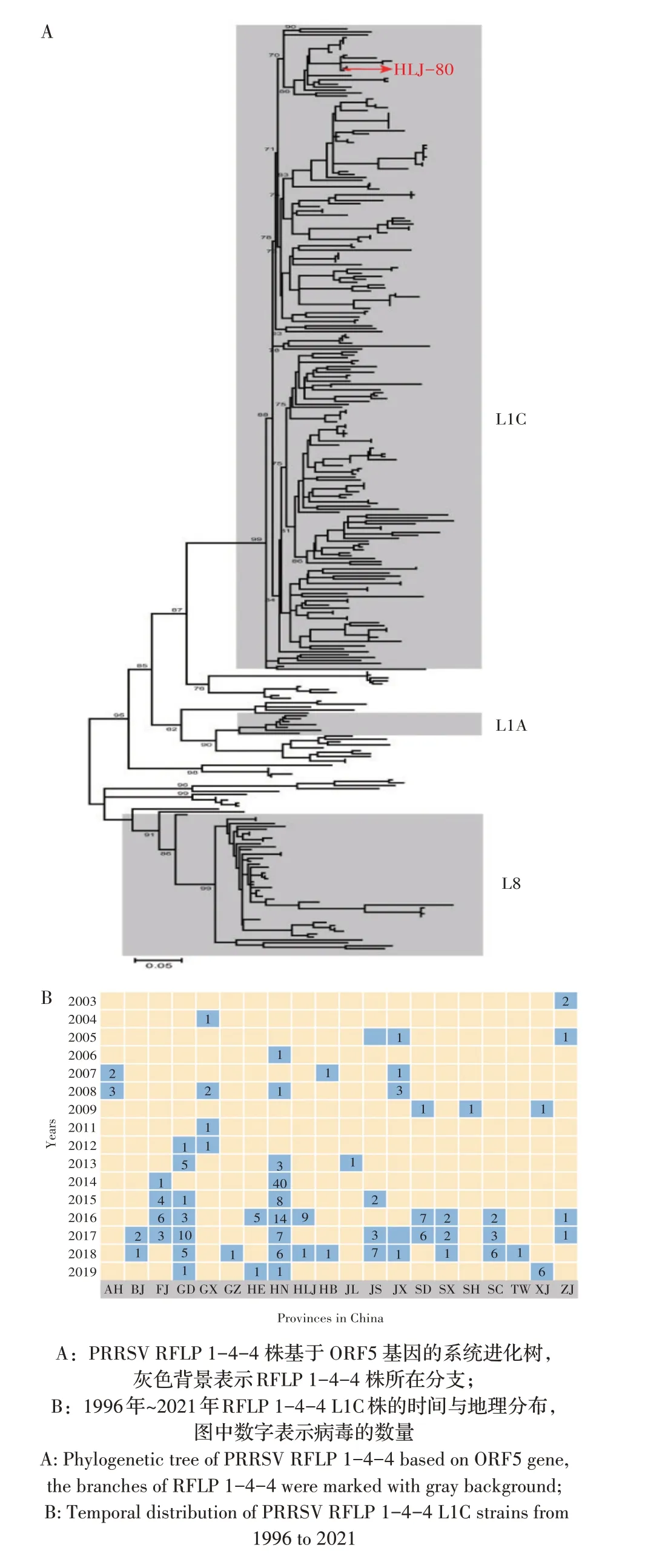
图4 1996年~2021 年中国PRRSV RFLP 1-4-4株的系统进化树及时间与地理分布Fig.4 Phylogenetic and spatiotemporal distribution of PRRSV RFLP 1-4-4 strains in China
2.7 我国与国外PRRSV 流行株RFLP 模式的比较分析统计2016 年~2021 年GenBank 中所有北美洲型PRRSV ORF5 基因序列的数量,从多到少依次为美国(n=1 167)、中国(n=635)、加拿大(n=65)、印度(n=29)、秘鲁(n=12)、韩国(n=7)和越南(n=3),共7 个国家1 918 条ORF5 基因序列。RFLP 分类结果显示,这些病毒株共有36 种RFLP 模式(图5A)。我国PRRSV 共有23 种RFLP 模式,美国28 种,加拿大11种,印度等国家RFLP 模式种类较少(图5B)。从分离年份上,2016 年~2018 年RFLP 模式复杂多样,2019 年~2021 年,各 国 上 传GenBank 的PRRSV 序 列较少(其中加拿大、印度和秘鲁等国家未上传PRRSV序列),该年间RFLP 模式较为单一,主要为1-4-4、1-4-2、1-4-3、1-2-4、1-7-4、2-5-2、1-3-4 和1-8-4(图5C)。具体而言,RFLP 1-7-4 的数量最多(n=498),占25.96%,主要分布在美国、中国和秘鲁。RFLP 1-4-3 株数量其次,占16.53%,主要分布在美国、中国、印度和越南,也是我国的优势RFLP 模式。RFLP 1-4-4分布最广泛,除越南和印度外,该模式在上述5个国家均有分布;其中我国的数量最多(n=159,含中国台湾1条),美国其次(n=129)。上述结果表明,中美两国PRRSV RFLP 模式较多,且PRRSV RFLP 1-4-4 模式早已存在于多个国家。
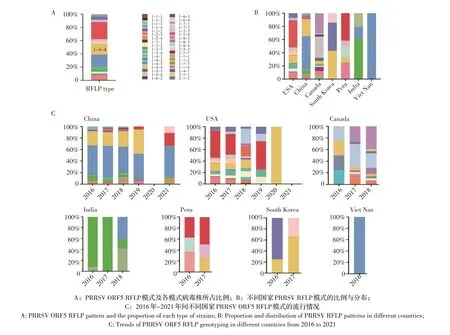
图5 2016年~2021年各国PRRSV RFLP模式及分布特征Fig.5 Classification and distribution of PRRSV ORF5 RFLP patterns from 2016 to 2021
3 讨 论
2020 年10 月,美国出现RFLP 1-4-4 L1C 变异株,该变异株ORF5 基因发生了重组[18],其大部分序列来自L1C 分支,所以在进化树中该株病毒被划分为L1C 分支,且在L1C 进化树中形成单独的一个分支。因有别于其他RFLP 1-4-4 L1C 株,被称为RFLP 1-4-4 L1C 变异株。目前我国尚无RFLP 1-4-4 L1C 变异株的相关报道。本研究从219 份疑似PRRSV感染的临床样品中分离出一株PRRSV RFLP 1-4-4 L1C 株,命名为HLJ-80 株。不同于美国1-4-4 L1C变异株NSP2 蛋白存在100 个氨基酸的连续缺失[18],HLJ-80 株与NADC30 株同源性较高且NSP2 蛋白均为131 个氨基酸(111+1+19)的不连续缺失。遗传演化分析结果显示,HLJ-80 株虽与美国1-4-4 L1C 变异株同属于L1C 分支,但两者全基因组序列的同源性仅为93.4%。综上,本研究分离的HLJ-80 株与美国1-4-4 L1C 变异株并不是同一株病毒。
为了解我国PRRSV RFLP 1-4-4 株的流行情况,本研究分析了1996 年~2021 年我国3 166 条PRRSV ORF5 基因,发现我国共有260 条(8.2%)ORF5 基因属于RFLP 1-4-4 模式,其中217 条属于L1C 分支(217/260,83.46%),其余为L8 和L1A 分支。由此可见,即使RFLP 模式相同的病毒株,它们之间的基因序列也可能存在很大的差异,这是由于PRRSV 的RFLP 分类是依据某些限制性内切酶酶切位点的数量和位置来命名的[10],在酶切位点中个别碱基的突变,即可改变RFLP 的模式。例如,美国RFLP1-4-4 大部分属于L1C 分支,也有L1A 分支、L5 分支以及L7~L9 分支[12],因此RFLP 模式不能反映ORF5 基因更多的遗传信息,更不能作为判定是否为美国变异株的唯一标准。美国将这两种分类方法结合起来,对新出现的病毒命名为RFLP 1-4-4 L1C 变异株,更多地体现出了该株病毒的遗传进化位置。
为进一步比较近年来国内外流行株RFLP 模式的差异,本研究分析了2016 年~2021 年国内外1 918 条PRRSV ORF5 基因,RFLP 分类结果显示,这些病毒共有36 种RFLP 模式,中美两国PRRSV RFLP 模式最多,这是PRRSV 在猪群中不断遗传演化的结果,印度等国家RFLP 模式种类较少,可能与这些国家上传到GenBank 中的ORF5 基因序列数量较少有关。此外,在上传序列至GenBank 的国家中(除越南和印度)均发现了RFLP 1-4-4 株,但无类似美国RFLP 1-4-4 L1C 变异株导致大规模发病的报道。其原因可能包括以下几方面,RFLP 分类和分支谱系分类均是基于ORF5 基因,该基因序列长度仅占PRRSV 全基因组的4%,仅能反映PRRSV 的部分遗传信息;重组赋予了病毒更复杂的生物学特性,RFLP 1-4-4 L1C 变异株[13]、中国农业大学研究团队从山东某猪场分离的L1 分支的RFLP 1-4-4 株[14]与本研究中分离的RFLP 1-4-4 L1C 株均属于重组病毒,且重组的亲本株和位置不一;另外,研究表明PRRSV 对猪的致病性更多受PRRSV 在猪体内复制效率的影响[19]。
我国和美国的养猪规模居世界前列,PRRSV 流行株也较其他国家更为复杂多样。本研究发现PRRSV RFLP 1-4-4 模式早已在多个国家存在,这些病毒的全基因组序列与美国新出现的RFLP 1-4-4 L1C 变异株全基因组序列有很大的差异。目前国内尚未发现与美国RFLP 1-4-4 L1C 变异株基因组序列高度同源的PRRSV。鉴于我国有从美国、加拿大引种和进口冷冻猪肉制品的贸易,以及有美国PRRSV传入我国的先例,因此有必要加强引种和进口冷冻猪肉制品的出入境检验检疫、加强国内猪群的流行病学监测、及与美国相关疫控部门或研究人员的信息沟通,共同抵御RFLP 1-4-4 L1C变异株的传播。本研究为了解我国RFLP 1-4-4模式PRRSV的流行情况提供了重要参考数据,为我国PRRSV的防控提供数据参考。
猜你喜欢
黑龙江大学自然科学学报(2022年1期)2022-03-29
计算机系统应用(2021年10期)2022-01-06
支部建设(2020年15期)2020-07-08
学生天地(2019年28期)2019-08-25
中国报道(2018年2期)2018-04-20
百科知识(2015年18期)2015-09-10
小星星·阅读100分(高年级)(2015年4期)2015-05-26
疯狂英语·口语版(2013年1期)2013-01-31
雕塑(1996年4期)1996-07-12
