
家畜认知功能及其调控机制
2022-08-26 08:49迟长安彭思祺申长庆王世成涂静怡邱小燕
畜牧兽医学报 2022年8期
迟长安,彭思祺,申长庆,王世成,涂静怡,肖 雄,邱小燕*
(1.西南大学动物科学技术学院,重庆 400715; 2.西南大学动物医学院,重庆 400715)
1 家畜认知功能及认知障碍概述
认知(cognition)是个体认识客观世界的信息加工活动。广义的认知包括感知、学习、记忆和决策,简言之,就是个体通过一系列活动,如感觉、知觉、记忆、思维、想象和语言等获取知识的能力或过程,这个过程就是认知过程。对动物而言,动物认知是指动物受到环境变化的刺激或应激时,动物个体做出的关联性学习和记忆,并做出适当的决定与反应的能力。家畜的认知功能和认知学习行为也逐渐被研究,前期采用目前国际上公认并广泛使用的羊认知功能的行为学表型测试方法——“认知识别测试”以及“左右空间Y-型测试”发现澳洲美利奴羊具有良好的认知识别能力,随后研究发现,同群测试羊中大概有15%~20%的个体具有认知功能障碍,且认知障碍对其母性表达、繁殖性能和生产性能具有重要影响,近期在白绒山羊上开展的认知行为相关研究也获得了类似的结论。除了羊,目前国内外在猪、牛、马等多种动物个体上也进行了认知识别相关研究。
家畜认知功能的调控非常复杂。研究发现,与认知功能关系较密切的脑区为前额叶皮质和海马。Gueniot等和Farfara等研究表明,认知障碍个体中,脑的损害是多位点、多层次的,涉及血脑屏障、血管、神经元,即所谓的“神经血管单元(neurovascular unit, NVU)”。另外,最新研究表明,线粒体功能异常包括氧化应激、Ca紊乱、动力学损伤及自噬缺陷,以及表观遗传修饰包括DNA修饰、组蛋白修饰和mRNA翻译等均可能通过对大脑认知区域基因表达的改变等从而对认知功能的调控发挥重要作用。因此,本文就近年来国内外在牛、羊、马、猪等多个物种个体上进行的认知识别研究及其对畜牧生产和繁殖性能的影响进行综述,并从“神经血管单元”、表观遗传修饰和线粒体功能等方面对认知功能的调控机制进行了较为全面的阐述,为家畜认知行为和认知功能的深入研究提供重要的参考。
2 家畜认知功能及认知障碍的国内外研究进展
2.1 牛的认知及认知障碍
2.1.1 牛认知功能的表型测试装置 牛认知功能的测试装置如图1所示,走廊尽头是一扇胶合板门(A、B之间),门上有一扇23 cm×28 cm小窗户。“培训师”留在门后面,门向识别测试区域敞开。该竞技场远端胶合板屏障高出地面40 cm,在初始培训期间,喂料箱被放置在A,在识别测试阶段,喂料箱被放置在B。盖子是可操作目标,悬挂在与眼睛水平的位置。用斜线标记的区域为“训练区”。测试牛在多次训练后是否能建立辨别食物的认知。
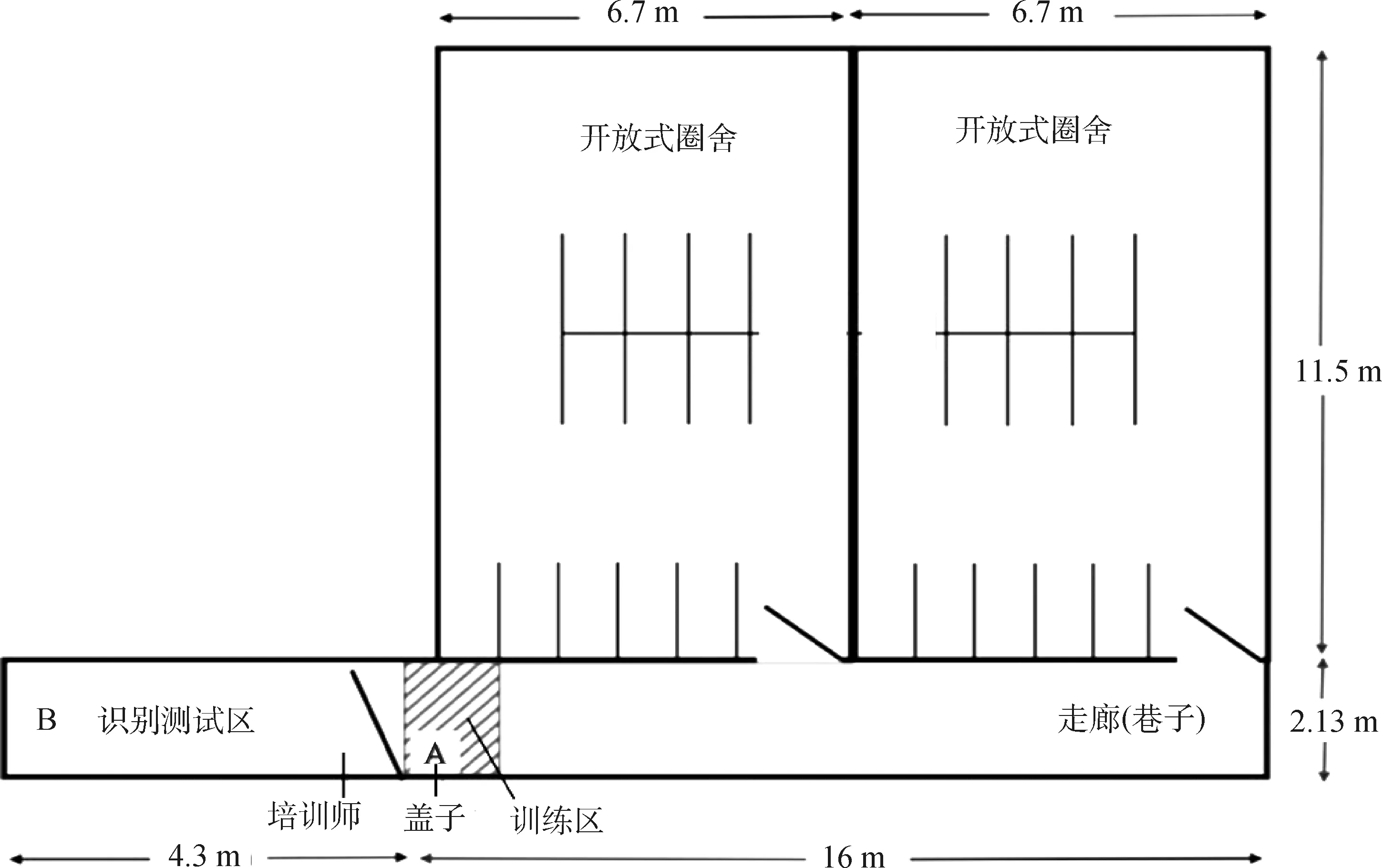
图1 牛认知识别测试装置的平面设计图[15]Fig.1 Schema of cow cognitive recognition test apparatus[15]
2.1.2 牛认知功能的表型测试方法 被测试牛首先需为最后的识别测试任务做准备,包括4个主要步骤:1)习惯化:通过让小母牛连续2 d进入训练小巷来完成,然后开始日常训练。在训练区内放置一个饲料箱(图1),并记录接近该区域的延迟时间,接近时间定义为小母牛头朝着该方向穿过距离大门约1 m的小巷一侧柱子的时间;2)点击器训练:当牛接近饲料箱时,培训师用响片发出信号,表示将提供食物奖励。在牛经历至少20对“咔嗒”与“谷物”的配对之后,开始学习操作反应;3)操作性反应和可变间隔训练:使用透明的蓝色塑料盖(8.25 cm×15.5 cm),悬挂在牛眼睛水平的绳子上,作为目标。牛用嘴触碰绳子或盖子会得到奖励,每次点击后会得到最多10次谷物奖励。一旦建立这一反应,就引入了可变间隔(variable interval, VI)训练,这样牛第一次触到盖子就会得到奖励,之后就有一个没有奖励的间隔。引入可变间隔随机产生,范围为20~40 s,平均30 s (VI-30);4)识别测试:一旦牛熟悉了VI-30训练,牛就被训练通过敞开的大门进入竞技场,在那里进行识别测试以获得奖励。
从操作性反应开始,所有牛在进入小巷时最多有6 min的时间接近训练区。如果他们未能进入训练区,他们将被送回原地。当牛完成训练后,引入辨别性刺激。这些刺激是牛必须从饲料箱上推开盖子,由纯白色塑料制成或覆盖着一层红色泡沫。它们的颜色和质地都各不相同,红色的泡沫盖总是表示箱子里有谷物,因此是“正确的选择”,而普通的白色盖放在装有稻草的箱子上,是“不正确的选择”。以做出正确选择与否以及做出正确选择吃到食物的时间为统计指标,统计分析认知识别能力的差异。
2.1.3 牛认知障碍对繁殖和生产性能的影响 研究表明,牛的认知障碍对其产奶性能、寿命和生产性能都具有不良影响。Neja等提取了11 629头波兰荷尔斯坦-弗里斯牛的产奶性能和寿命等数据,研究发现,认知功能正常的牛在产乳性能方面更好,寿命也较认知障碍的牛要长。Choudhary等研究发现,穆拉水牛认知功能障碍导致卵巢循环性的恢复延迟,且体重、身体状况和产奶量都有所下降。Vann等研究发现,牛的认知障碍对后代的性情、体重、日增重和母性表达均有不良影响。
认知障碍除了影响牛的繁殖性能和泌乳性能外,对其生产性能和胴体性能也有很大的影响。Bruno等研究发现,牛的认知障碍影响饲养牛的日增重和日增重指数。Olson等研究发现,与认知障碍的肥育母牛相比,认知正常的肥育母牛生长效率更快、饲料效率更高和胴体品质更好。Cooke等研究发现,认知障碍的内洛尔肉牛生长速度减慢,血浆皮质醇浓度升高,青春期发育受阻。Braga等研究发现,认知障碍的内洛尔公牛在日常饲养中表现出更大的行为反应性,在面对压力时有更强烈的应激反应。随后Sant Anna等研究发现,与认知正常的内洛尔公牛相比,认知障碍的内洛尔公牛其胴体肉质更差。White等研究发现,认知障碍对婆罗门牛的骨骼肌线粒体功能有一定的影响,但其内在联系还需要进一步研究。Caroline等研究发现,在放牧系统中,认知障碍影响内洛尔牛的进食行为和生长性能。
2.2 羊的认知及认知障碍
2.2.1 羊认知功能的表型测试装置 采用目前国际上广泛使用的羊认知功能的行为学测试方法-“视觉认知识别测试(visual discrimination test)”和“空间认知识别及颠倒测试(spacial Y-test and reversal test)”。
2.2.1.1 视觉认知识别测试装置:羊的视觉认知识别测试装置(图2)由2个相邻的测试区(灰色和白色区域; 2.4 m×7.2 m)组成。测试装置四周均用遮光布与外界隔离。每个测试小区域中间有隔板(图2),隔板末端的门可以向左右关闭(图2虚线),每个测试小区域中分别有两种“刺激”(代表两种颜色“蓝色”和“黄色”装有食物豆子的桶和空桶;颜色不同,但形状一样,一个装有豆子,一个为空桶)。本测试目的在于测试羊在通过多次训练后是否能建立颜色与食物的联系认知。
2.2.1.2 空间认知识别及颠倒测试装置:空间识别测试装置(图3)由3个臂组成(A、B和C),测试装置四周均用遮光布与外界隔离。将装有食物的盆放在B或C臂末端。本测试目的在于测试羊在通过多次训练后是否能建立左右与食物的联系认知,从而记住哪个方向代表食物,进而在变化方向后即如原来左边有食物,变为右边有食物(颠倒识别,reversal learning)时,是否又能将右边与食物联系起来的这种左右空间识别认知能力。
2.2.2 羊认知功能的表型测试方法
2.2.2.1 视觉认知识别测试方法:被测试羊从“入口门”(图2)进入。开始的训练阶段是:先将“蓝色”与食物相联系,即如图2第一个测试小区域(灰色区域)左边“刺激”即蓝色装豆子桶,右边“刺激”即黄色空桶,被测试羊在进入后,会面临选择,如果羊跑向左边蓝色桶,则立即将中间的铁栅栏关闭,使羊在正确区域待上1 min,保证羊能吃到蓝色桶的豆子;如果羊在进入后,错跑向了右边黄色空桶,同样立即将中间的铁栅栏向右关闭,给予1 min,让羊发现右边黄色桶中无豆子。羊完成第一个识别后,进入第二个测试小区域(图2,白色区域)完成下一个识别,识别原理和过程同上。以整个一套(8次)测试中,被测试羊能6次直接跑向正确一边,即要达到75%正确率视为羊学会了这种识别,具有简单认知识别能力。以做出错误选择的次数以及做出正确选择吃到食物的时间为统计指标,统计分析认知识别能力的差异。
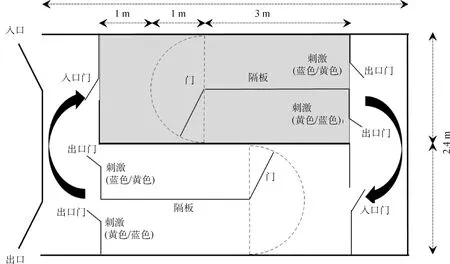
图2 视觉认知识别测试装置的平面设计图[1](根据Morton和Avanzo研究[2]改造而成)Fig.2 Schema of visual discrimination test apparatus[1] (based on Morton and Avanzo[2])
上述颜色识别完成后进行颠倒测试,即将原先“蓝色桶装豆子”颠倒改变成“黄色桶装有豆子”,测试羊在一定数量训练后能不能将原先建立起来的蓝色与食物的认知联系转变成黄色与食物的认知联系,学会标准和统计指标同上。
2.2.2.2 空间认知识别及颠倒测试方法:被测试羊从“入口门”(图3)进入,在通道(图3中A臂)末端将会面临左右(图3中B臂和C臂)的选择。开始的训练阶段是:先将装有食物的盆放在左边B臂末端(C臂末端为空桶)。被测试羊在进入后,会看见B臂和C臂末端的桶,面临选择,允许被测试羊在3个臂之间来回走动(以每个臂的虚线为准判断进入哪一个臂)。被测试羊如能连续4次跑向正确一边(如装食物盆在左边,即左边B臂为正确选择),视为羊学会了这种识别,具有简单的左右认知记忆能力;反之,被测试羊学不会,其左右认知记忆功能障碍。以正确选择前走错臂的次数和做出正确选择吃到食物的时间为统计指标,分析认知能力的差异。
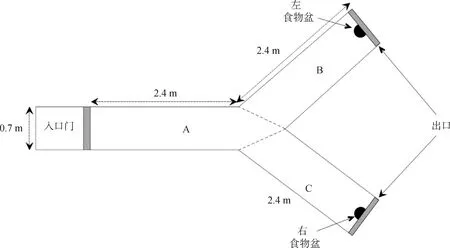
图3 左右空间的识别记忆测试(Y-test)平面图[1]Fig.3 Schema of the Y-maze apparatus[1]
上述左右识别记忆测试完成后进行颠倒测试,即将原先“左边B臂放食物”颠倒改变成“右边C臂放食物”,测试羊在一定数量训练后能否将原先建立起来的左边与食物的认知联系转变成右边与食物的认知联系,经过一定数量的反复训练(次数会比之前的稍多,因为要有一个发现原来的认知联系错误,纠正,然后建立相反方向的认知过程),学会标准和统计指标同上。
2.2.3 羊认知障碍对繁殖和生产性能的影响 研究表明,羊的认知障碍对其繁殖性能包括同期发情率、怀多胎比率以及后代存活率等方面都具有不良影响。前期研究发现,认知障碍的中国美利奴羊的排卵率和怀多胎比率显著低于认知正常羊;认知障碍羊的母性表达包括对出生后羔羊的舔舐时间、允许羔羊的吮吸时间等均显著低于认知正常羊,且认知障碍羊的后代死亡率显著高于认知正常羊。Hart等研究发现,认知正常的母羊具有较高的同期发情排卵率和孪生率,在产后最初6 h内,其初乳更黏稠,且免疫球蛋白G浓度更高、乳糖比例更大、乳脂浓度更低。Pajor等研究发现,认知障碍的母羊在产奶期产奶量较低,且断奶前增重和断奶体重较认知正常的母羊更低,且认知正常母羊的羔羊比认知障碍母羊的羔羊更平静,具有更好的育肥性能。随后又发现在育肥结束时,认知正常的羔羊皮质醇浓度低于认知功能障碍的羔羊,且胆固醇和葡萄糖较低,日增重显著增高。Gavojdian等研究发现,认知正常的杜泊羊母羊比认知障碍母羊更多产,活体重更高,母羊的小崽体型更大,出生至断奶羔羊生长速度更快。
认知障碍除了影响羊繁殖性能和母性表达外,对羊的生产性能也有很大的影响。Zhang等研究发现,认知正常的湖羊,其羔羊的干物质消化率较高,血清TNF-α水平较低, 总抗氧化能力、总超氧化物歧化酶活性、屠宰率、腰横截面积、肌纤维密度、肉的最终pH以及羔羊育肥结束时体重和日增重均高于认知障碍的湖羊
2.3 马的认知及认知障碍
2.3.1 马认知功能的表型测试装置 马的认知识别测试装置(图4)由两个面板(140 cm×90 cm×3 cm)组成,被放置在一个小围栏中。在面板表面有两个洞:右边是一个黄色圆圈,左边是一个蓝色三角形(从动物的角度来看),操作者(图4中“人1”)手持有两个胡萝卜片,作为奖励插入其中,被测试动物识别选择。另一个操作员(图4中“人2”,总是在动物的左边)带领马至面板前面1.5 m,马可以自由地从两只手中选择一只。
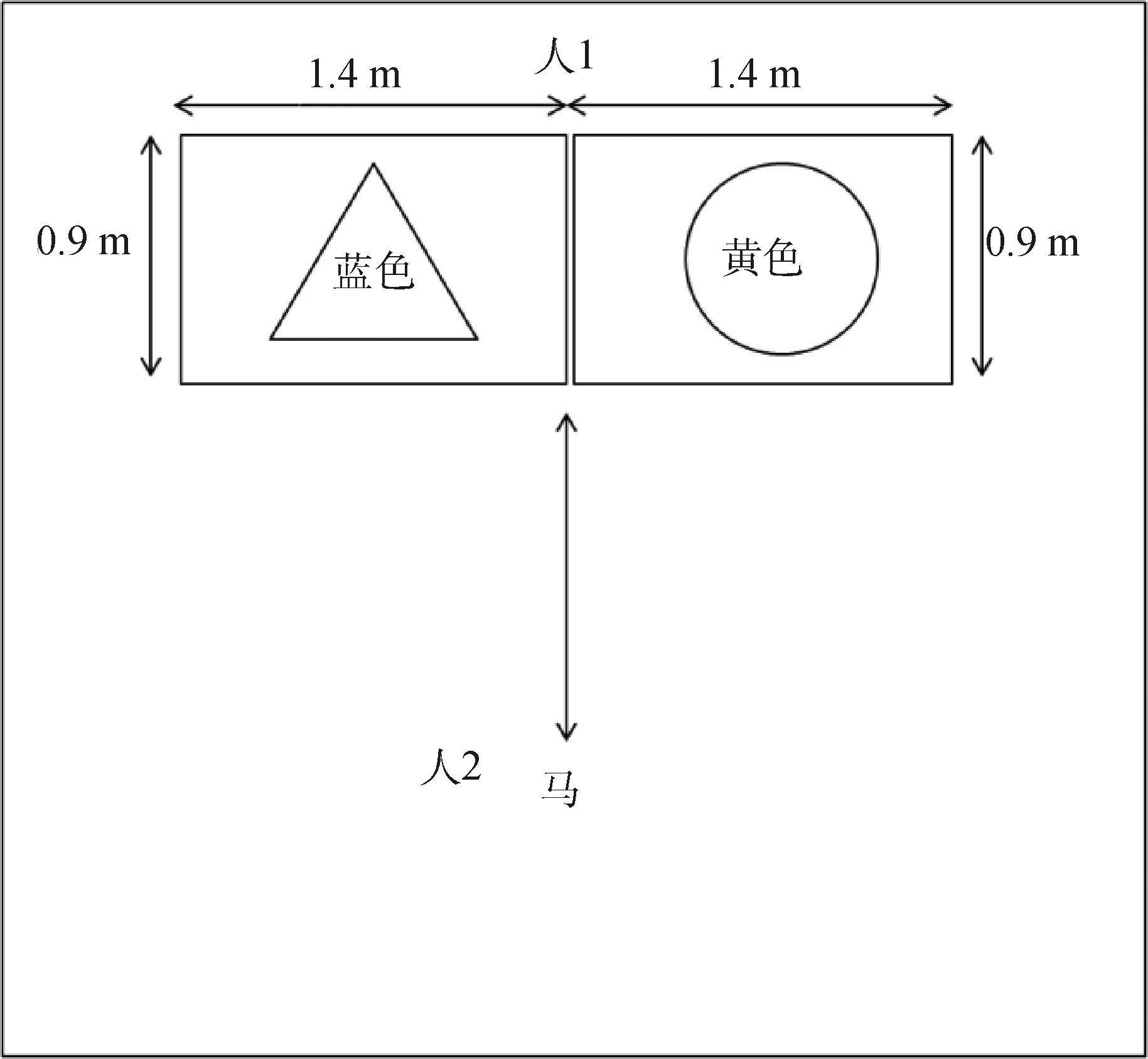
图4 马的认知测试模型[32]Fig.4 Schema of horse cognitive recognition test apparatus[32]
2.3.2 马认知功能的表型测试方法 马在两个面板前(图4),在蓝色三角形和黄色圆圈之间进行选择。所有操作员停止任何互动,尽可能减少对马做出选择的影响。每匹马都习惯于两个面板,有一个预定的奖励设置(当动物对设备或操作者表现出积极反应时,就会给予奖励)。在第一次(学习)和第二次(回忆)阶段,训练马面临区分两个几何图形并选择黄色圆圈(通常在马的右边)。在第三次中,马必须选择相反的颜色图形(颠倒;当位于马的左边的蓝色三角形成为新的正确选择)。第二次和第三次在同一天连续进行,从第一次后的2~3周连续进行。如果马在2 min内成功连续做出4个正确选择,则被认为通过测试,具有这种认知识别能力,以做出正确选择与否以及做出正确选择吃到食物的时间为统计指标,统计分析认知识别能力的差异。
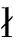
2.4 猪的认知及认知障碍
2.4.1 猪认知功能的表型测试装置 猪食物奖励与颠倒学习测验装置(图5)采用T型迷宫。奖励的食物碗(直径18 cm)只含有压碎的香蕉或含有压碎的香蕉和草莓酱。目的在于测试猪是否具有学习认知能力。
图5 食物奖励与颠倒学习测验装置平面设计图[39]Fig.5 Schema of food reward and reverse learning test apparatus[39]
2.4.2 猪认知功能的表型测试方法 被测试猪在5 d的时间内进行12次食物奖励试验,包括训练个体将随机选择的T型迷宫手臂(图5)与食物奖励(即用压碎的香蕉作为诱饵的熟悉食物碗)相关联。食物奖励的位置在所有试验中保持一致。为了尽量减少视觉和嗅觉的影响,在T型迷宫的对侧臂(即错误臂)放置一个先前含有食物奖励的非诱饵碗。在每次试验中,猪通过位于中心的门进入迷宫,并记录找到诱饵碗所需时间。如果猪在进入迷宫180 s内找到食物奖励,而没有进入不正确的手臂,则认为猪“成功”识别了T型迷宫。在找到食物之前进入迷宫错误臂的个体被记录为“不成功”。如果食物奖赏在进入迷宫后180 s内未找到,则该个体被视为“未通过”试验,给予180 s的最大潜伏期,并由一名研究人员轻轻引导其走向诱饵碗。为了确保猪持续测试的动力,从第六次试验开始,在猪已经习惯于围栏中的新奖励后,在食物碗中添加草莓酱(除了压碎的香蕉)。
之后进入颠倒学习试验,诱饵碗被重新定位到之前不正确的迷宫臂,而非诱饵碗被放置在之前正确的位置。同样,记录猪只发现带诱饵碗所需的时间,进入第一只手臂的时间,进入每个手臂的次数。以做出正确选择次数以及做出正确选择吃到食物的时间为统计指标,统计分析学习认知识别能力的差异。
2.4.3 猪认知障碍对繁殖和生产性能的影响 研究表明,猪的认知障碍对其繁殖性能及生产性能都具有不良影响。D’Eath等研究发现,与认知正常母猪相比,认知障碍的母猪在遇到陌生个体后表现出的攻击性行为增加,导致皮肤损伤及生产性能的下降,严重影响猪的福利和经济效益。Yoder等研究了4 774头白切斯特公猪、杜洛克公猪、长白猪、约克郡公猪和小母猪的核心群体来估计认知的品种差异及其与生产性能的关系,结果表明,不同品种猪的认知水平不同,认知障碍的猪体肥胖,腰深宽大,对瘦肉率有不利的影响,且生长较慢。认知障碍对猪与人类的互动行为方面也有所影响。Rault研究发现,认知正常的猪,脑脊液中含有更高浓度的催产素(OXT),且与人类有更积极的互动,猪的福利方面较认知障碍猪也更好。
3 家畜认知功能的调控机制
3.1 “神经血管单元(neurovascular unit, NVU)对认知功能的调控
认知障碍个体中,脑的损害是多位点、多层次的,研究发现“神经血管单元”(包括血脑屏障、血管、神经元)对认知功能具有非常重要的调节作用。
3.1.1 认知障碍与血脑屏障的损伤 研究表明,若血脑屏障的屏障功能和转运功能受损,个体对葡萄糖、钠、钾、胆碱和胰岛素等重要物质的通透性增加,引起脑微循环的血流动力学、微血管生化物质以及神经递质活性改变,甚至微血管闭塞,进而影响中枢神经系统内环境的稳定性,最终影响认知功能。Takechi等在小鼠上的研究发现,血脑屏障的破坏或损伤导致小鼠认知功能障碍,Geng等也得出类似结论。血脑屏障的特异性结构是血管内皮细胞及其紧密连接,紧密连接由细胞骨架蛋白、跨膜蛋白和胞质附着蛋白共同组成。跨膜蛋白包括3种膜蛋白:闭锁蛋白(occludin)、封闭蛋白(claudin)和黏着分子(junctional adhesive molecule, JAM);胞质附着蛋白有闭合小环蛋白ZO-1、ZO-2和ZO-3三种。研究显示,血脑屏障被破坏时,紧密连接蛋白Occludin、Claudin-5、ZO-1等的表达均降低。因此,增加Occudin、Claudin-5、ZO-1等的表达,修复血脑屏障紧密连接有可能成为认知障碍治疗或改善的有效途径。
3.1.2 认知障碍与血管功能的损伤 研究表明,认知障碍的脑损伤修复过程中,神经血管单元的多种细胞之间相互作用共同参与神经血管的重塑。在外周血中存在一种细胞,是血管新生的重要物质,称为内皮祖细胞(endothelial progenitor cells, EPCs),可进一步分化为内皮细胞而参与受损害血管内皮的修复与血管新生。研究表明,认知障碍个体中存在EPCs数量的减少以及增殖能力、黏附能力和血管形成能力的下降。Zhang等研究发现,增加EPCs的数量可提高小鼠的认知功能。低氧诱导因子-1ɑ(hypoxia-inducible factor 1α, HIF-1ɑ)和血管内皮生长因子(vascular endothelial growth factor, VEGF)被认为是EPCs迁移增殖、血管新生重构过程中起重要作用的2个因子。HIF-1ɑ是哺乳动物组织细胞在缺氧/缺血条件下产生的一种核转录因子,VEGF是HIF-1ɑ的下游靶基因。Zhu等研究发现,HIF-1ɑ和VEGF能特异性促进内皮细胞有丝分裂,增加血管通透性,促进EPCs增殖和新生血管形成。因此,增加EPCs数量及VEGF的表达来促进血管新生也有可能成为认知障碍治疗或改善的有效途径。
3.1.3 认知障碍与神经元的损伤 神经血管单元(neurovascular unit, NVU)除了血脑屏障、血管,还有一个非常重要的构成就是神经元。众多关于认知障碍的研究都表明,认知功能损伤的同时往往伴随前额叶皮质和海马神经元的损伤。在小鼠/大鼠上的研究发现,神经炎性因子(Fas-L、IL-6、iNOS、TNF-ɑ、MMP-9、MCP-1)和神经营养因子(GDNF、BDNF)参与神经元的损伤和修复。那么神经炎性因子或营养因子是如何调节神经元的可塑性和数量从而影响神经元功能的呢?研究表明,神经元突触的可塑性与认知功能密切相关。研究发现,环磷酸腺苷反应元件结合蛋白(cAMP response element-binding protein, CREB)参与认知和记忆相关蛋白的表达及新突触生成,在神经元再生和突触形成等方面具有重要的调节作用。PI3K/PKB/CREB是CREB激活通路中非常重要的一条信号通路,PI3K(磷脂酰肌醇-3-羟激酶)活化后可激活PKB(蛋白激酶B),使CREB磷酸化,增加突触可塑性基因及相关蛋白的表达。研究表明,神经营养因子通过PI3K/PKB信号通路参与神经元细胞凋亡的调控,从而影响认知和学习记忆等相关功能。
3.2 表观遗传修饰对认知功能的调控
表观遗传失调可能导致大脑认知区域基因表达的改变,是认知功能障碍疾病的关键病理生理基础之一。与认知功能调控相关的表观遗传修饰包括DNA修饰、组蛋白修饰和mRNA翻译(micro-RNAs)等。
因此,DNA甲基化和羟甲基化在神经元分化和功能过程中协调基因组结构和基因表达水平,从而在大脑发育和记忆形成中发挥基础性作用,它们的损伤与神经发育密切相关,对认知障碍疾病的改善和治疗有着至关重要的作用。
3.2.2 组蛋白修饰 组蛋白修饰作为基因表达的表观遗传调控的一个主要机制,已经成为探索治疗认知功能障碍神经退行性疾病病理基础的研究热点。乙酰化是组蛋白共价修饰的重要调节方式之一。研究表明,组蛋白去乙酰化酶(histone deacetylase, HDACs)能使组蛋白的赖氨酸残基去乙酰化造成组蛋白的低乙酰化,抑制核小体结构的充分打开,从而抑制基因的转录。前期研究发现,HDACs引起神经细胞组蛋白去乙酰化,造成组蛋白的低乙酰化,抑制了神经细胞存活相关基因或突触可塑性相关基因的表达,进而导致了记忆和认知功能缺损等一系列的病理变化。I类HDACs,尤其是HDAC1、HDAC2和HDAC8是抑制神经炎症,预防和治疗认知功能障碍的潜在治疗靶点。HDACs抑制剂(histone deacetylase inhibitors, HDACis) 由于能抑制组蛋白去乙酰化从而恢复或增加组蛋白乙酰化水平,因此有望用于认知障碍的神经疾病治疗。近年来,几种 HDACis包括丙戊酸(VPA)、丁酸(SB)、曲古菌素A(TSA)、伏立诺他(SAHA)、苯基丁酸钠(NaPB)在国内外已经被应用于认知损害性疾病模型中,且发现对认知损伤具有一定的恢复和改善效应。Okado研究发现,类锌指转录抑制物(POK家族)的许多重要分子,包括RP58(也称为Znf238、Znp238和Zbtb18),通过染色质重塑和组蛋白去乙酰化调节神经元产生、迁移和成熟的各种生理过程。
因此,组蛋白修饰尤其是乙酰化修饰在认知功能调控中发挥重要作用,组蛋白去乙酰化酶抑制剂HDACi为认知障碍疾病的治疗提供了新的表观遗传机制和潜在的治疗策略。
3.2.3 mRNA翻译 研究表明,mRNA的降解或翻译是通过与microRNAs相互作用而实现的。microRNAs(miRNAs)是一类新的短(18~25个核苷酸)单链非编码RNA,参与基因表达的转录后调控。研究表明,miRNA对细胞生长、细胞凋亡、癌症和与衰老相关的认知功能障碍具有调控作用。Liu等对认知障碍鼠侧脑室注射miR-134-5p拮抗剂,发现其可以阻止突触蛋白的丢失和认知功能障碍表型的形成。Wu等研究发现,认知障碍小鼠的miR-181a表达下降,过表达miR-181a可改善认知障碍。Sun等研究了miR-144在体内外对认知功能的影响,发现在体内抑制miR-144可使神经功能得到改善,尤其是认知功能障碍的改善;在体外,miR-144敲除通过增强细胞活力、抑制caspase-3活性以及降低认知相关蛋白水平来保护神经元损伤。Tang等研究表明,miR-192-5p表达水平的上调改善认知障碍,增强认知功能。因此,miRNAs调控基因表达是治疗或改善认知障碍的另一个有效途径。
3.3 线粒体对认知功能的调控
3.3.1 线粒体氧化应激和ATP缺失 研究表明,认知障碍疾病中的线粒体功能障碍来自于神经元ATP水平的下降,与过量的自由基ROS产生有关,因此ROS的异常导致线粒体功能障碍是认知障碍疾病发病机制的重要因素。线粒体ATP合成酶是控制ATP产生、细胞信号和命运等细胞功能的关键枢纽。Cunnane等发现,线粒体ATP合成酶由ATP酶抑制因子1(IF1)调节,Esparza-Moltó等通过在小鼠神经元中切除或过度表达IF1来控制线粒体功能和线粒体活性氧(mtROS)的产生,发现IF1剂量可以调节线粒体代谢、突触功能和认知,IF1的消融会损害记忆,而IF1的过度表达会增强突触传递和学习能力。因此,阻断ROS恢复ATP合成酶的活性,可以成为改善线粒体功能从而治疗认知功能障碍性疾病的新策略。
3.3.2 线粒体Ca紊乱 研究表明,Ca在重要的神经元过程如分泌、代谢调节、突触可塑性、增殖和凋亡中发挥重要的调节作用。Diaz-Juarez等研究发现,游离线粒体钙浓度是激活线粒体呼吸链复合物和ATP生成的基础,且可调节关键线粒体脱氢酶的活性,线粒体Ca除了可调节参与三羧酸循环(TCA)的Ca敏感脱氢酶的活性从而直接控制ATP合成的作用之外,线粒体基质中过多的Ca浓度会升高线粒体活性氧水平,导致线粒体通透性转换孔(mPTP)的开放,释放细胞色素c到细胞质中,最终触发内源性凋亡,因此细胞Ca稳态对认知功能至关重要。Kostic和Sekler发现,线粒体Ca单转运蛋白(MCU)及关键调节因子线粒体Ca摄取蛋白(MICU1、MICU2和MICU3)、MCU调节亚单位b(MCUb)、必需MCU调节因子(EMRE)和线粒体Ca单转运蛋白调节因子1(MCUR1)在介导线粒体Ca稳态中起着关键作用。
Zhang等研究表明,下调MCU相关信号通路,抑制神经元的凋亡,可以实现脑保护作用。Calvo-Rodriguez等以神经元线粒体为靶点研究发现,在脑β-淀粉样变性转基因小鼠模型中线粒体Ca水平升高与斑块沉积和神经元死亡相关,Aβ积累通过线粒体Ca单转运体(MCU)复合物诱导体内线粒体Ca过载,导致神经元死亡。Fross等研究表明,抑制钙蛋白酶激活可防止神经元细胞骨架蛋白断裂和神经元细胞死亡,表明钙蛋白酶激活是外周神经退行性病变的关键执行者。因此,线粒体钙(Ca)动力学作为校准细胞能量学和生存功能的关键决定因素之一,是研究认知功能障碍疾病的重要机制,而MCU复合物抑制和阻断mPTP的激活可能成为认知功能障碍疾病新的治疗方法。
3.3.3 线粒体动力学损伤及自噬缺陷 线粒体在形状、大小、数量和位置上是处于动态变化中的。线粒体的质量和稳态通过裂变(分裂)和融合维持,裂变与融合是线粒体生物学和质量控制的基本方式。Stanga等研究表明,线粒体动力学缺陷(如裂变和/或融合过程的紊乱)可导致神经退行性疾病。在线粒体动力学的中心阶段,动力相关GTP酶(GTPases)驱动线粒体分裂和融合,维持线粒体分裂和融合之间的平衡。裂变由GTPases动力相关蛋白-1(Drp1)和动力蛋白-2(Dyn2)执行,而GTPase蛋白丝裂分裂蛋白-1(Mfn1)、丝裂分裂蛋白-2(Mfn2)和视神经萎缩1型(OPA1)蛋白促进聚变。Duboff等研究发现,Drp1向线粒体的募集是裂变的关键步骤,而Drp1向线粒体的定位募集受到肌球蛋白II调控。因此,调控线粒体分裂和融合可以成为改善线粒体功能,治疗认知功能障碍的新策略。
同时,线粒体的不断更新对于维持健康线粒体的数量至关重要。Cai和Jeong研究发现,线粒体数量和大小增加的过程是通过一种线粒体自噬机制完成的,线粒体自噬是一种选择性的自噬,是线粒体质量数量控制的关键途径,有缺陷的线粒体被隔离成自噬体进行随后的溶酶体降解。线粒体自噬功能障碍影响细胞的蛋白质稳定,从而对记忆和认知造成损伤,与衰老和许多认知功能障碍类疾病密切相关。PTEN (磷酸酶-张力蛋白)诱导的激酶蛋白1(PINK1)-Parkin(E3泛素连接酶)介导的线粒体自噬是研究最广泛的线粒体吞噬途径,当线粒体因细胞应激而受损时,其内膜发生持续去极化,导致线粒体膜电位的丢失,稳定线粒体外膜(OMM)中的PINK1。PINK1磷酸化Mfn2(丝裂分裂蛋白-2),然后刺激泛素-蛋白酶体系统(UPS),反过来,将Parkin招募到线粒体外膜(OMM),进一步促进了受损的线粒体被吞噬,形成线粒体吞噬体,最后通过溶酶体去除,维持线粒体质量控制。因此,稳定线粒体中的PINK1,促进受损线粒体降解,维持线粒体健康与更新有望成为认知功能障碍治疗的新方向。
4 总结与展望
国内外越来越多的研究开始关注家畜认知行为和认知功能。目前,已在猪、马、牛、羊上开展了认知相关的研究,虽然表型测试装置和方法有细微区别,但测试原理大致一样,均是通过食物诱导,训练并测试动物将食物与左右空间、物体颜色、形状等的联系识别和认知记忆能力,且家畜的认知障碍均影响其繁殖性能、生产性能和动物福利。
家畜认知功能的调控非常复杂,本文从“血管神经单元”、表观遗传修饰和线粒体三方面较为全面地阐述了认知功能的调控机制(图6)。但认知障碍致病因素及发病机制非常复杂,还有待更深入的研究来揭开复杂的认知功能调控过程。
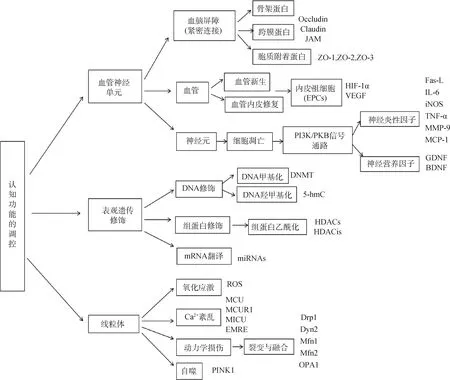
图6 认知功能的调控机制Fig.6 Regulation mechanisms of cognitive function
猜你喜欢
中国现代医生(2022年20期)2022-11-04
电子产品世界(2021年8期)2021-01-16
健康之家(2020年7期)2020-11-02
祝您健康(2019年8期)2019-08-09
环球时报(2019-06-21)2019-06-21
中国医药导报(2019年13期)2019-06-20
中国计算机报(2019年49期)2019-02-07
养生大世界(2018年10期)2018-10-18
中国新闻周刊(2017年36期)2017-10-21
创新时代(2016年8期)2016-10-21
