
绵羊经济性状相关基因研究进展及其应用
2022-08-26 08:49李亭亭刘秋月李向臣王海涛
畜牧兽医学报 2022年8期
李亭亭,刘秋月,李向臣*,王海涛*
(1.浙江农林大学 动物科技学院·动物医学院,杭州 311300;2.中国科学院遗传与发育生物学研究所,北京 100101)
绵羊()起源于现亚洲中西部的新月沃地区,是世界上最早被人类驯化的家养动物之一。根据联合国粮农组织(Food and Agriculture Organization of United Nations,FAO)的统计数据,我国是世界上最大的羊肉生产国,其产量几乎占世界总产量的三分之一。随着人们对羊肉需求的增加,针对地方绵羊品种繁殖、生长以及抗病性状等方面的选育越来越受到重视,育种方式也越来越多样化。
绵羊在上万年的进化过程中经历了长期的人工和自然选择。在不同地区和环境中的绵羊群体为了适应自然环境或人类喜好,会对自身体型、毛色、繁殖、耐受等生理表型进行调整,还会在基因组上留下不同印记,通过对绵羊品种中不同印记进行检测,就能够对动物的环境适应性机制有更好的理解,同时如果能够找到影响这些表型的遗传变异,就能极大提升育种的效率。在绵羊上,目前研究者主要针对繁殖、生长、脂尾、抗病及抗逆等几个与经济效益关联紧密的性状,通过家系遗传定位、比较基因组学、候选基因关联分析以及全基因组关联分析等手段,先后鉴定获得绵羊中多个包括增加排卵数、提高生长速度、增加产肉量以及增强抗病能力的主效基因以及致因位点,如图1所示。其中部分基因已经大范围运用于绵羊育种实践,大部分正处于动物个体水平验证的阶段。
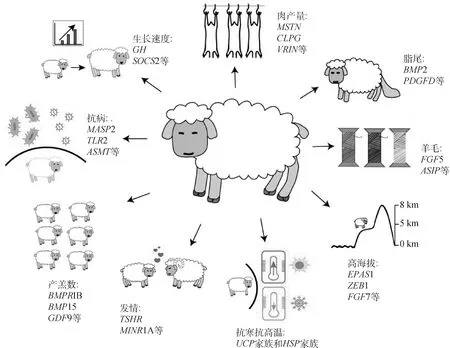
图1 绵羊重要经济性状及其相关基因Fig.1 Important economic traits and related genes of sheep
随着遗传修饰技术的进一步发展,一套新的育种策略和方法已经逐渐形成。研究者可以通过转基因或基因编辑等方式直接对获得的候选基因进行体内功能验证,极大的缩短了大动物中候选主效基因的验证周期。针对定位获得的候选基因,最普遍的做法就是通过CRISPR/Cas9系统将基因进行直接敲除,观察动物表型以确认基因功能;通过CRISPR/Cas9结合同源重组或供体DNA,可以在不完全破坏功能基因的前提下针对特定基因型进行定点编辑;更进一步,随着单碱基编辑系统的开发,目前研究者在无需依赖基因组产生双链断裂的情况下,可以进行高效定点的基因编辑,或者利用先导编辑(prime editor,PE)技术对基因进行插入、缺失等突变体的编辑以确认致因突变的价值。本文将目前已研究的相对清楚并影响绵羊重要经济性状的相关基因和应用情况进行了总结,为以后的绵羊分子育种工作提供参考。
1 绵羊重要经济性状及相关基因
1.1 繁殖性状
绵羊的繁殖性状是影响养羊业最重要的经济性状,可体现在产羔和季节性发情两个方面。目前在绵羊上鉴定出的重要多羔主效基因的多个突变主要集中在变异生长因子β(transforming growth factor-β,TGF-β)通路的相关基因上,主要有、15、9,另外还包括近年来鉴定出的基因,它是唯一一个不属于TGF-β信号通路的调控绵羊排卵数的基因。而对于影响绵羊季节性发情的研究,鉴定出的基因主要有、1A,可通过绵羊对光照的反应来调控体内相关生殖激素的合成,从而影响发情。
1.1.1 产羔数性状 产羔数性状是养羊业经济性状中最重要的生产性状,受母羊排卵数、子宫容量等因素影响,其中排卵数可直接决定母羊的产羔数和繁殖力。母绵羊产羔数与排卵数的相关性达到0.96,但产羔数性状的遗传力较低,约为0.11,而排卵数性状遗传力相对较高,自然群体中约为0.19左右,而在选择群体中可高达0.30。雌性绵羊在出生前,卵巢上便形成了大量原始卵泡。初情期前,卵泡虽能发育,但不能成熟排卵,当发育到一定程度时,便闭锁退化。初情期后,在每个发情周期中可发育的卵泡多达几十个,但单羔品种最终能发育到成熟排卵的卵泡一般只有1~2个,其余卵泡中途闭锁而死亡,而多羔品种则有多个卵泡能发育成熟并排卵。
目前发现的多羔主效基因大部分都属于TGF-β信号通路成员。基因(fecundity booroola)是第一个被发现与绵羊产羔数有关的主效基因,1994年,Montgomery等利用微卫星标记和遗传连锁的方法将与Booroola Merino绵羊多产相关的基因定位到绵羊6号染色体上,之后发现该基因实际为绵羊骨形态发生蛋白受体1B型基因(bone morphogenetic protein receptor type 1B,1B)编码区发生了A746G突变,造成其编码的氨基酸由谷氨酰胺变为精氨酸(Q249R),该位点的突变减弱了BMPR1B在BMP信号通路中的配体功能,导致卵母细胞的加速成熟并排卵,使母羊排卵数增加。经过大量研究证实,突变在Booroola Merino绵羊、Javanese Indonesia、小尾寒羊、湖羊、Garole绵羊和Kalehkoohi等多产绵羊品种中均有发现。TGF-β家族中分别位于绵羊X和5号染色体上的骨形态发生蛋白15(bone morphogenetic protein 15,15)和生长分化因子9(growth differentiation factor 9,9)基因,是由欧洲科学家利用自然突变的高产羔数绵羊群体,构建遗传连锁图谱,利用微卫星扫描或者全基因组关联分析(genome-wide association study,GWAS)定位获得的影响绵羊产羔数的主效基因,二者有高度的氨基酸序列同源性,通常共同形成异源二聚体作用于BMPRⅡ型受体,并激活Smad2/3(sma- and mad-related,Smad)和Smad 1/5/8信号通路,促进颗粒细胞的增殖及分化、卵泡发育以及增加排卵数。迄今为止,在不同绵羊品种中已识别出15基因的10个与排卵率、繁殖力相关的点突变,分别具有不同的功能,即、(终止密码子提前使蛋白合成受损),、(通过改变阅读框),(导致15蛋白功能丧失),和(与9蛋白相互作用受损),和(细胞信号活性改变)以及(未知)。与其他突变不同的是,、和的纯合突变不仅不会导致母羊不育,反而使其具有更高的繁殖能力。在9中发现了11个突变,分别是G1~G8,以及、、,这些突变均可对排卵和产羔造成影响。目前,基因编辑技术已经运用到大动物模型上,研究者们利用CRISPR/Cas9技术获得了具有g.A746G点突变的绵羊,但有关这些突变羔羊繁殖能力的研究还未见报道。吴艳芳不仅获得了突变的滩羊,而且还证明具有突变的滩羊比野生型滩羊具有更高的产羔数,这些研究为获得突变的绵羊从而提高绵羊产羔数提供了一个新的方向。而对于15和9,可能是由于其效应位点较多难以选择或一些绵羊群体中存在自然突变,很少有关于其在绵羊上的报道,但是有研究人员在猪上引入15无义突变后,观察到纯合突变导致母猪不育,而杂合突变则可增加母猪排卵数,这一发现与存在自然突变的绵羊群体中观察到的现象一致。
上述3个与多羔相关的主效基因中,多个突变位点的效应已经在世界范围内多个绵羊品种中得到证实。近年来研究者还利用其它绵羊群体陆续在这3个基因中发现了新的突变,可为绵羊产羔性状研究提供新的参考。另外,研究者还通过转录组及代谢组等分析手段,发现在这3个基因发生突变的个体中,与胰岛素分泌、cAMP、MAPK、Smads、Hippo等相关的信号通路被显著富集,且突变个体卵泡液中的葡萄糖6-磷酸、天冬氨酸、半胱氨酸、谷胱甘肽等重要代谢物与排卵率的增加显著相关,这些研究结果将有助于深入阐明绵羊多羔性状的发生机制。
除TGF-β信号通路之外的产羔数主效基因也已有报道。基因是法国农科院Bodin等从1975到1996年间通过5个世代的选育构建Lacaune绵羊的回交家系,又经过基因组扫描、高通量测序以及分子生物学验证,于2013年定位到了4GALNT2(glycosylation enzyme beta-1,4-N-acetyl galactosaminyltransferase 2)为的主效基因,可能的机制是突变导致了4GALNT2基因在卵泡中的异位表达,此蛋白在颗粒细胞中的高表达会使卵泡中特定靶蛋白的糖基化状态改变,进而引起了排卵数的改变。这一发现揭开了除TGF-β信号通路之外调控绵羊排卵数的全新机制。除了基因外,最新的研究也鉴定到一些新的与绵羊产羔相关的重要基因,并发现了这些基因上存在的影响产羔数的多个突变位点,例如、、、等基因,这些功能基因突变之后,会对繁殖相关的生物学过程产生影响,如可通过影响排卵过程而导致产羔数的变化,及其受体可参与卵泡的成熟和发育,影响胚胎附着和排卵过程,可能通过参与卵母细胞的发育而影响产羔数,则可影响绵羊卵巢和卵泡发育。另外,研究者还利用产羔数显著差异的绵羊个体进行比较生物学研究,从转录组和蛋白层面鉴定到了多个相关基因及信号通路,如4、1、2B、1等可分别参与昼夜节律、代谢和胰岛素分泌合成等相关激素调节过程;、和1能影响GnRH活性和生殖激素的释放;4可调节卵泡发育和卵母细胞成熟;3与能协同促进卵泡成熟和排卵;5可与卵泡发育相关的蛋白质相互作用,参与卵泡生成,这些结果使研究者对影响绵羊繁殖性状的机制多了新的认识。
1.1.2 发情性状 季节性发情是限制绵羊繁殖能力的瓶颈因素之一,经过研究者们几十年的探索,现在基本确定了哺乳动物中光照通过下丘脑-垂体-性腺轴(hypothalamic-pituitary-gonadal axis,HPGA)系统引起季节性发情,其机制为:眼球视网膜接受光照刺激产生视神经冲动,并由下视丘传至下丘脑视交叉上核(suprachiasmatic nucleus,SCN),SCN作为动物体内最主要的生物钟,将信号传至松果体,引起松果体中褪黑素(melatonin,MEL)分泌的改变,之后褪黑素通过HPGA轴进一步引起下丘脑中促性腺激素释放激素(gonadotropin releasing hormone,GnRH)以及相关生殖激素的合成、释放,最终调控动物的季节性发情活动。近年来,促甲状腺激素受体(thyroid-stimulating hormone receptor,)基因和褪黑素受体1A(melatonin receptor 1A,1A)基因被认为是影响绵羊季节性发情的重要调控基因,可作为改良绵羊季节性发情的重要分子标记。
最初在狗的甲状腺中被发现,之后通过对生活在不同环境下的野生和家养品种进行选择分析,发现在鸡和绵羊的驯化过程中均受到强烈选择,并且可通过HPGA轴通路反馈调节季节性繁殖活动。目前认为的调控方式是下丘脑中的促甲状腺激素β亚基(thyrotrophin β subunit,)可与结合,通过cAMP通路调控四碘甲状腺原氨酸(tetraiodothyronine,T)与三碘甲状腺原氨酸(triodothyronine,T)的转换,进而调节哺乳动物对光照周期的应答与季节性发情。研究发现,在Soay绵羊中光照长短能通过调节的表达从而影响绵羊的季节性繁殖活动。通过对国内多个地方绵羊品种的重测序以及分群验证,证实该基因的多态性与中国本土绵羊的季节性发情行为和繁殖活动显著相关。1A基因于1996年首先在西伯利亚仓鼠中被发现,参与介导褪黑素对哺乳动物生殖活动的影响,随后在Merinos d’Arles、Aragonesa、Sarda、小尾寒羊等绵羊品种中陆续发现1A基因与绵羊卵巢的排卵活动和产羔频率相关。目前,研究者发现绵羊1基因的不同突变不仅与绵羊的季节性发情和产羔有关,同时某些突变还对雄性绵羊的繁殖活动(如外阴嗅探、爬跨等)产生促进作用,被公认是调节哺乳动物繁殖活动的重要基因。
1.2 生长性状
畜禽生长性能包括生长速度、活体背膘厚及饲料转化率等,而其中生长速度可体现在肌细胞的增殖发育以及骨骼的生长速度两个方面。一方面,肌肉的形成和再生需通过一个高度精密的调控过程,使成肌细胞分化和融合成多核合胞体。在胚胎发育过程中,中胚层前体细胞受到周围组织信号的控制分化为肌源细胞,其中需要包括Pax7、MyoD和Myf5等因子的上调,增殖的前体细胞/成肌细胞停止细胞周期并诱导肌肉特异性基因表达,之后通过一系列高度有序的程序先发生肌细胞之间的融合形成新生的肌管,随后募集和融合额外的成肌细胞产生多核肌管,最终形成成熟的肌肉纤维。另一方面,骨骼系统包括硬骨、软骨、肌腱和韧带等,共同起源于中胚层和神经嵴产生的间充质干细胞,进一步分化为构成特定骨骼类型的细胞,这个过程受多种信号通路和转录因子的调节,如FGF、IGF1、MAPK、PI3K/Akt通路都可参与软骨细胞增殖和分化的调控,软骨形成后一部分作为永久性软骨,另一部分可作为生长板形成的模板,这些模板可形成钙化骨,成为纵向生长的骨骼,BMP信号通路在此过程中起着关键作用。在软骨中,肥大的软骨细胞产生Ihh(indian hedgehog),作用于生长板增厚带附近的软骨膜细胞,诱导成骨细胞分化,成骨细胞成熟后,产生骨基质蛋白,包括I型胶原蛋白和骨钙素,使骨骼有一定的弹性和硬度。
生长激素(growth hormone,)基因是研究较多的促进生长发育的基因,也是线性骨生长的主要调节剂,可通过全身和局部胰岛素样生长因子1(insulin-like growth factor 1,IGF1)的产生来刺激骨骼生长,使机体的生长速度更快。分子育种技术运用之初,就被通过转基因的方式导入绵羊体内获得生长更快的个体,但是近年来研究较少。
肌肉生长抑制素(myostatin,),也称为生长和分化因子8(growth and differentiation factor 8,8),位于绵羊2号染色体,主要表达于骨骼肌中,在肌肉分化和生长过程中起负调控作用,通过部分抑制成肌细胞的增殖来调节肌肉质量,不同畜种中的“双肌现象”已被证明均是基因突变的结果。研究表明,可降低肌肉发育相关基因7和的表达,阻断成肌细胞的细胞周期,使其停滞在G1期,抑制成肌细胞分化。因此,当基因敲除之后,蛋白功能受损,则会产生相反的效果,促进肌细胞的增殖和分化,导致双肌性状。迄今为止,已经在多个物种中发现的突变与肌肉质量增加相关,在不同绵羊品种中也发现了多个突变位点,其中大多数位于该基因的非编码区,这表明位于编码区之外的遗传变异在调节肌肉发育方面有重要作用。研究表明,绵羊基因3′UTR区的SNP可将其表达水平降低约三分之一,且在显著增加肌肉占比的同时不影响肉品质。近年来,随着CRISPR/Cas9技术的广泛应用,研究者已利用原核注射或体细胞核移植的方式获得成功敲除的绵羊个体,这些基因编辑个体与野生型绵羊相比,体重显著增加,骨骼肌纤维肥大,具有双肌性状。利用基因编辑的方法能够快速验证候选基因在绵羊中的效应并评价其运用效果,对绵羊产业的未来发展具有重要经济价值。
细胞因子信号转导抑制因子2(suppressors of cytokine signaling,2)是SOCS家族的一员,作为GH信号通路和骨骼生长的负调节因子,可通过SH2结构域与生长激素受体(growth hormone receptor,GHR)结合,对JAK/STAT5b通路起到抑制作用,影响机体的生长、骨骼发育以及新陈代谢等,表型上可对动物的骨骼线性增长和体重增加产生抑制作用。当2发生突变时,导致蛋白结构发生改变,功能丧失,与受体的结合也被破坏,从而调节下游的GH/IGF1通路,对动物的生长产生促进作用。研究表明,2小鼠体型可比野生型大30%~40%,而且这种体重增加是由于内脏器官、骨骼长度以及身体长度的显著增加引起的。Rupp等于2015年首次在Lacaune绵羊中定位到与绵羊体型大小、体重及产奶量显著相关的QTL,区段中包含2基因,当编码SOCS2蛋白SH2结构域的区段发生p.R96C点突变时,其编码的蛋白结构也发生改变,导致其与GHR的结合能力丧失,与野生型绵羊相比,p.R96C纯合突变的绵羊的体型、体重分别增加了24%和18%。之后,Zhou等对绵羊2基因进行单碱基编辑,将3 mRNA和sgRNA共同注射到绵羊受精卵中,实现了2中p.R96C的单碱基替换,获得具有点突变的绵羊,证明该基因突变对于绵羊体重和体型有促进作用。
除以上提到的基因以及、、基因都可以影响绵羊的生长性状,基因与有类似的效应,也是目前研究人员公认的与绵羊臀部肌肉发育有关的基因,是影响绵羊肋骨数的重要分子标记,而能够影响绵羊的眼肌宽度、深度及面积,这些分子标记都可对以后的绵羊育种提供重要的指导。
1.3 脂尾性状
绵羊尾部脂肪是肥尾绵羊储存能量的重要组织,反映了绵羊对恶劣和极端环境的适应性,尾部堆积的大量脂肪不仅可以增强绵羊在极端环境下的耐寒能力,也可作为迁徙和食物缺乏期间维持机体所需的重要能量来源,对于人类来说,它还可以在干旱和饥荒期间为人类提供高能量的食物。脂肪细胞来源于间充质干细胞,尾部脂肪的沉积过程包括脂肪细胞的增殖、分化和成熟阶段。间充质干细胞首先分化为前体脂肪细胞,再转化为具有脂质运输和合成、分泌脂肪细胞特异性蛋白质等功能的成熟脂肪细胞,之后沉积在内脏器官、皮下或尾部,可为机体维持正常的生命活动提供能量。这些过程由复杂的基因网络之间相互协调,涉及PPAR、MAPK、Wnt/β-catenin、BMP/SMADs、胰岛素/IGF1等相关通路,多与脂质形成和代谢相关。然而,近几十年来,随生活水平的提高和饮食习惯等的改变,绵羊大脂肪尾巴作为能量储备的优势逐渐减少,生产者和消费者对低脂肉类更感兴趣,形成相同质量的尾部脂肪比生成等量的肌肉组织需要付出更大的饲养成本,且相同质量的肥尾的价格远低于瘦肉价格,肥尾还会对母羊繁殖能力产生负面影响,过多的尾部脂肪沉积极大的降低了养羊业的经济效益。因此,探索调节绵羊尾部脂肪沉积和发育的候选基因及遗传分子机制对培育尾脂较少的绵羊品种具有重要意义。
通过基因组学方法,已经定位到数个与绵羊尾部脂肪沉积相关的候选基因,研究较多的为血小板衍生生长因子D(platelet derived growth factor D,)和骨形态发生蛋白2(bone morphogenetic protein 2,2)基因,在不同的绵羊脂尾相关研究中被多次提到,证明2和基因是影响绵羊肥尾表型的主要因素。其中在全世界不同尾型的绵羊中选择信号最强,通过激活PDGFRβ来调节PPARγ2和C/EBPα的表达,抑制白色脂肪细胞分化,也可以通过促进成熟脂肪细胞的形成或增殖来影响脂质代谢(如脂质合成),造成绵羊尾部的脂肪沉积。2是脂肪发生的重要调节因子,不仅能诱导干细胞分化为脂肪细胞,还可能有助于将能量储存分配到内脏和皮下脂肪组织中。Zhu等的研究表明,2的错义突变(Ala>Val)与绵羊肥尾表型直接相关,并参与尾部脂肪沉积。最新研究也表明,2和基因的SNP可能与阿勒泰绵羊的尾脂沉积相关,但这些基因的具体作用机制还不清晰,后期可作为重要的候选基因继续研究和验证。
近年来有多项全基因组关联研究通过利用地方绵羊品种定位到了多个影响脂尾性状的候选区域,其中,1、、1CC、101117953等基因在绵羊尾型性状中受到强烈选择。1作为促脂肪因子,可能在胎儿发育过程中对中胚层细胞的分化起关键作用,并影响绵羊尾部的脂肪沉积;(也称为)参与脂肪的发育和功能,是控制脂肪生成、分化和发育的关键转录因子,在脂肪细胞分化的早期阶段就已开始发挥作用;1CC位于2附近,与2有很强的相关性,101117953为1的反转录本(-PPP1CC),但由于大多数反转录拷贝被认为是没有功能的,所以猜测其可能与1相似,通过影响上游的2来发挥作用。目前,对绵羊脂尾性状的研究仍主要集中在2和两个基因上,研究者们也逐渐发现了其他有效应的突变位点,但还需进一步对其进行功能验证和作用机制的探索。除此之外,一些新的相关基因和通路也逐渐被发现,也可作为研究其作用机制的参考。
1.4 抗病性状
在畜牧业养殖中,疫病常为养殖业带来巨大的经济损失。而在绵羊中,感染源通常包括细菌、病毒、寄生虫等,防治办法通常有添加抗生素、疫苗预防或治疗,但都会存在相应的问题,比如产生耐药株及疫苗失效等。不同品种的绵羊在对疾病的抵抗力方面存在差异,一旦通过现代基因组学方法找到与某些疾病抗性相关的主效基因以及确定相关突变,就可以更深入地了解与抗性相关的分子基础和遗传变异,结合现代基因操作技术就有可能获得稳定的绵羊抗病新品系。动物的抗病性状的是病原体在与宿主免疫系统对抗过程中被打败的结果,其抗性基因的定位大多也集中在病原体识别、免疫因子的分泌、抗感染及相关通路方面。
线虫是寄生虫的一种,而胃肠道线虫(gastrointestinal nematode,GIN)是对绵羊产业危害最大的一种线虫,胃肠道线虫病是影响全球畜牧生产系统的主要健康问题,动物感染之后可在死亡率、生产力和治疗成本方面给全世界造成重大经济损失,澳大利亚绵羊产业中每年约有4.3亿美元用于与线虫感染相关的生产损失和治疗成本,是世界各地绵羊和其他放牧反刍动物的最重要的健康疾病之一。在过去的几十年中,养羊业越来越依赖驱虫药物作为一种控制寄生虫病的方法,然而,这种方式不仅花费巨大又通常不是很有效,还会导致耐药株的出现。该病对反刍动物的影响主要反映在体重减轻、腹泻、厌食、贫血和死亡等方面,而幼龄羔羊又特别容易受到线虫感染,这是影响绵羊生产的主要经济负担,因此,大多数关于GIN抗性的QTL逐渐被定位出来。Al Kalaldeh等使用GWAS发现位于绵羊2号染色体上的、6基因与寄生虫抗性之间存在很强的关联,2、32也被发现与内部胃肠道线虫感染的抗性相关。另外,绵羊对线虫的抗性及恢复能力也可能与介导病原体识别(、和基因家族)、先天免疫反应(、、、20R)以及对感染的适应性反应(、2R、-)方面的基因有关,这些结果都可为研究绵羊抗胃肠道线虫感染提供参考。
布鲁氏菌病由布鲁氏菌()感染所致,它是对反刍动物养殖和人类健康危害极大的人畜共患病,该病主要对动物的生殖系统造成危害,不仅能造成母羊流产、不孕不育,也会造成雄性动物生殖障碍,严重影响牲畜生产性能。科研人员对于布鲁氏菌病的抗性基因也进行了很多的挖掘,目前也有一些收获。Li等通过构建褪黑素合成酶(acetylserotonin O-methyltransferase)基因过表达载体,成功获得具有抗布病的转基因绵羊,并证明基因过表达绵羊可能通过影响免疫相关的信号通路和肠道微生物群来增强对布鲁氏菌病的抵抗力。另一团队通过全基因组重测序分析,确定了4种与绵羊布鲁氏菌病易感性相关的重要途径以及相关的候选基因,即粘附小带(3、3和)、细胞粘附分子(1、、1和)、唾液分泌(101102109、1和2)和Hippo信号通路(3、1、3),这些基因与细胞粘附过程密切相关,可能有助于机体发挥免疫功能的第一道屏障。
目前,病原菌疾病相关的研究很少是在建立动物模型的基础上来开展的,大多数仍然是在群体中通过抗病个体来寻找可能的相关基因,或直接在发病个体或细胞水平研究其致病机制,从而来研制治疗方法,原因可能是目前大家对这些疾病深入的致病机制仍不清晰,导致即使想要构建动物疾病模型却无从下手。
1.5 羊毛性状
绵羊毛作为一种天然的蛋白纤维在绵羊养殖业和纺织业中都占有重要的经济地位,也是毛用绵羊的重要经济性状,其品质和质量可体现在羊毛的长度、细度、强度等方面。毛发生长发生在毛囊发育的生长期,始于真皮乳头部周围的基底膜干细胞,随着细胞系分化的开始而停止分裂,之后细胞的形状和位置停止变化,纤维细胞系开始合成角蛋白(keratin,K),皮层逐渐角质化,非角蛋白细胞成分逐渐分解,在上层的毛囊中,被皮脂腺和汗腺包裹的富含油脂的纤维从体表长出,形成毛发。羊毛纤维主要是由角蛋白组成的角质层,羊毛角蛋白包括含量较稳定的角蛋白中间丝蛋白(intermediate filament proteins,IFPs)和影响羊毛特性的角蛋白联合蛋白(keratin associate proteins,KAPs)两种,而角蛋白基因的表达可影响羊毛的弯曲、粗细、重量等重要经济特征。
绵羊的羊毛性状是受遗传、环境和营养等因素影响的复杂生理生化特性,许多信号通路和相关因素都参与到这些经济特征的调节中。成纤维细胞生长因子是一系列在促进和调节成纤维细胞生长中起重要作用的因子,其中成纤维细胞生长因子5(fibroblast growth factor 5,5)基因已被证实是毛发生长的抑制剂,沉默5基因的表达会延长毛发周期的生长期VI期,导致极长毛发的表型,而通过靶向使5功能丧失可抑制真皮细胞的活化和毛发纤维的合成,促进羊毛的生长和增加羊毛产量。除了5基因,影响绵羊毛色的刺鼠信号蛋白(agouti signaling protein,)基因也是影响羊毛质量的重要基因,在哺乳动物中可抑制黑色素皮质素受体1(melanocortin 1 receptor,1R)信号转导和黑色素的产生,调控毛发的颜色,Zhang等利用基因编辑技术破坏其功能,使绵羊的毛色发生改变。另外,角蛋白相关蛋白基因的表达及其多态性对绵羊羊毛直径、韧性、弯曲程度的影响也已在多项研究中被报道。
1.6 抗逆性状
极端气候地区的动物品种通过长期的自然选择及人工选择,在生理、生化、形态等方面获得了稳定的遗传特征,以适应高海拔、低氧、寒冷、干旱或高温等恶劣环境。因此,极端环境中具有特殊环境适应性的物种是挖掘抗逆机制以及候选基因的资源宝库,如果能在全基因组水平上了解其遗传基础并阐明这些分子标记,解析动物不同适应性表型的遗传基础,可为未来气候环境变化背景下制定适当的育种计划提供方向。
1.6.1 高海拔适应性状 生活在高海拔地区的动物为了适应低氧带来的挑战,在生理、细胞水平都发生了适应性变化,比如机体通过降低代谢活性、增加呼吸速率和红细胞总量来维持氧气运输,以满足动物维持其正常生长、发育和繁殖所需的组织氧气水平。此外,高原动物还会启动线粒体氧化代谢和信号转导通路,引起一系列的生理变化,使动物能够在缺氧条件下生存,而缺氧诱导因子(hypoxia-inducible factor,HIF)通路及相关因子在这其中发挥了重要的作用。藏羊是一种耐低氧物种,生活在极其荒凉的高海拔环境中,与低海拔地区相比,该地区紫外线辐射强、温度低、氧分压低。藏羊除了提供优质的羊毛和肉类外,还具有抗粗饲和适应高海拔缺氧等优点,可作为一种对极端环境具有较强适应性的模型,从中发掘一些有助于指导育种实践的关键基因。目前已有大量关于高海拔适应性的研究报道,例如藏獒(1、1)、牦牛(17、2和3)、藏羚羊(、12和3)和藏野猪(1、42和7)等,这些基因功能涉及缺氧、能量代谢、血管生成、Ca代谢、促进红细胞生成等方面。
利用高原上的低氧来完成正常的生理功能,这是机体在低氧的恶劣环境中生存的一种有效方式。近些年的研究报道了许多与低氧适应性相关的候选基因,其中研究最为透彻的基因是1和1,这两个候选基因都是在HIF通路上游起作用的关键基因。1(endothelial PAS domain protein 1,1),又称为2α,通过对不同海拔地区的山羊进行外显子组测序和基因分型分析,发现1多态性在藏绒山羊中显示出强烈的选择信号,而Q579L突变对藏绒山羊适应高海拔环境起了关键作用。在绵羊上,Wei等发现,1基因3′UTR的一个SNP与藏绵羊适应高海拔、促进红细胞生成有关,另外,该研究也发现了一些与代谢相关的基因,如4能维持葡萄糖稳定性,参与糖酵解过程;与磷脂代谢相关;1可激活ATP生成并减少线粒体ROS的产生。绵羊的靶基因1以及位于7号染色体的7基因在高海拔地区的绵羊群体中受到显著的选择,可能参与高海拔条件下细胞环境中可用氧的调节或其他调节。对于1基因,在绵羊中还未见到有关其与高海拔适应性相关的报道。
高海拔地区的生物除了要面对高海拔带来的缺氧和寒冷问题之外,还要应对紫外线强度对身体带来的损伤,因此紫外线信号通路扮演着重要的角色。在哺乳动物中,高强度的紫外线辐射可诱导大量ROS的产生,导致DNA损伤、细胞凋亡、组织损伤和癌症,破坏细胞内环境。Zhang等对藏鸡的研究发现,与炎症和免疫反应以及与紫外辐射导致的DNA损伤刺激和DNA修复过程相关的基因受到选择。在对绵羊的研究中,发现藏羊的和基因、印度肥尾羊的3和3基因受到紫外线照射方面的选择。从以上研究可以看出,目前参与高强度紫外线辐射适应性的研究较少,发现的相关基因也有限,并且相互之间的相似性很低。
除了以上提到的相关基因外,高海拔适应相关的5、、和3基因也受到强烈选择,但还需要进一步的研究和功能验证。
1.6.2 抗寒冷性状 由于中国北方高海拔地区有着漫长而严酷的高寒冬季,绵羊经常出现体温过低的情况,因此,抵御寒冷的能力对其生存至关重要。为了在处于低温环境时维持体温,绵羊必须增加机体的产热(生热),这包括颤抖生热(shivering thermogenesis,ST)和非颤抖生热(non-shivering thermogenesis,NST),一是通过骨骼肌的快速收缩水解ATP释放热量,二是通过中枢神经系统释放去甲肾上腺素,激活棕色脂肪细胞表面的受体或募集富含解偶联蛋白1(uncoupling protein,UCP1)的产热脂肪细胞分解甘油三酯产生热量。据报道,同一品种内不同品系的绵羊抗寒能力存在差异,而绵羊抗寒能力增强的遗传力约为0.27。
在与抗寒相关的研究中,UCP家族通常受到强烈选择。该家族主要包括1、2和3,三者之间有高度同源性,1主要存在于线粒体内膜中,是棕色脂肪的标志物,2在全身各个组织中均有表达,3主要分布于骨骼肌和心组织中。先前的报道称,该家族基因与不同家畜的寒冷适应性有关,存在于绵羊15号染色体上的1基因参与寒冷诱导的非颤抖产热,2和3基因在绵羊骨骼肌中的表达对于其适应寒冷发挥着重要作用。通过将印度的一种高海拔绵羊品种Changthangi与分布在平原地区的绵羊品种相比较,15号染色体基因家族存在最显著信号,说明该基因可能是参与绵羊适应寒冷气候环境的重要分子标记。
1.6.3 耐高温性状 从春季到夏季,气温和环境湿度均升高的综合影响会导致农业动物的热应激。热应激对动物的饲料消耗、生长速度和繁殖效率影响比较明显。据统计,热应激对全球畜牧生产的经济影响超过12亿美元。研究认为,热应激会诱导ROS的产生和脂质过氧化,可造成DNA和蛋白质的损伤,而ROS的产生被认为是诱导热休克蛋白基因(heat shock protein,)表达的关键因素。热应激对动物健康有严重影响,它可能导致牲畜的多器官衰竭,降低其生产力和繁殖效率,严重时可造成死亡,但是在沙漠地区的炎热气候下,仍有保持正常生长的绵羊品种,这表明在绵羊上存在耐热性的遗传基础。
目前发现,与绵羊热应激相关的基因大多都是属于家族或相关成员。热休克蛋白,也被称为分子伴侣,可由各种应激诱导表达,被认为是细胞应激的保护剂,可以介导降解受损细胞内的蛋白折叠,这种保护机制使细胞能够在高温条件下生存。此外,也能促进细胞的生长和分化,调节细胞凋亡,而且还和免疫功能相关。的激活主要是通过热休克转录因子(heat shock factor 1,HSF1)实现的,HSF1是一种响应热应激而被激活的转录因子。白绵羊中发现,90AA1(HSP90伴侣)基因的多态性可有效克服热应激并保持正常的生产性能,8基因中的T177P多态性也与Arabi 绵羊对炎热气候的高度耐热适应性有关。另外,多浪羊与中国本土其他绵羊品种遗传距离较大,具有良好的抗逆性和耐热性,Wang等发现5(dnaJ heat shock protein family (Hsp40) member B5)基因多态性可能参与多浪羊的热应激,5为DNAJ热休克蛋白40家族伴侣蛋白的一个成员,在免疫细胞的热应激耐受性中起着至关重要的作用。
2 总结与展望
很久以前人们就观察到某一性状在不同个体中的差别,但是由于当时技术的局限性,只能通过传统的杂交育种方式获得所想要的性状,而对其背后隐藏的生物学机制发掘进展缓慢。随着测序技术的飞速发展,基于基因组重测序以及GWAS等遗传定位研究方法的普遍运用,针对绵羊经济性状主效基因的研究也较之前有较大进展。目前,鉴定出的部分绵羊经济性状主效基因及效应影响如表1所示,其中部分基因的作用机制已比较明晰,也成功构建了动物模型对其功能进行验证。另外通过遗传定位的方式获得了一些候选基因,但还需要进一步阐明机制和验证其功能。再加上近年来基因编辑技术的迅猛发展,这为快速建立基于基因编辑的动物模型,验证候选基因在个体水平的效应提供了解决方案,也为未来建立具有优良性状的绵羊新品系提供参考。

猜你喜欢
中国现代医生(2022年19期)2022-11-04
水产养殖(2022年8期)2022-09-21
中国现代医生(2022年19期)2022-08-25
中国典型病例大全(2022年11期)2022-05-13
情感读本·道德篇(2022年2期)2022-04-12
儿童时代·幸福宝宝(2019年11期)2019-12-13
农民致富之友(2017年19期)2017-10-21
数学大王·低年级(2016年8期)2016-05-14
中学生物学(2008年11期)2008-12-02
作文与考试·高中版(2008年11期)2008-11-21
