
哈茨木霉对镉胁迫下草地早熟禾的促生和增抗效应及其生理机制
2022-09-08 01:31董斯琳姚桐桐谢琳淼孙广玉敖红
浙江农业科学 2022年9期
董斯琳,姚桐桐,谢琳淼,孙广玉,敖红
(东北林业大学 生命科学学院,黑龙江 哈尔滨 150040)
重金属镉(Cd)的生物毒性很强,是植物生长发育的非必需元素[1]。近年来,工业和农业生产中将镉不同程度地排放到土壤环境中,使镉污染成为危害最大、分布最广的农业污染之一[2]。据统计,中国已有2 000万hm2的土地面积被镉污染,并且污染面积正逐步扩大[3]。过量镉会对动植物产生极大危害,植物体内的镉过量时会产生毒害症状,主要表现为抑制根部增长,破坏某些生理生化反应,进而遏制根部从土壤吸收水分和营养物质[4]。植物的生长发育过程同样会受到镉的抑制,其结果会导致植物的生长迟缓,植株萎蔫,甚至导致死亡[5]。更重要的是,镉可以通过食物链进入人体,对人体造成巨大损伤[6]。对重金属土壤的修复有多种方式,其中植物修复受到人们的关注。而草坪草因其生命力旺盛、覆盖力和适应性强、分布广、抗逆性强、易收割等优点在植物修复重金属污染土壤中具有较好的应用价值[7]。
木霉菌是一类被广泛应用的植物生防菌,能抵御多种植物病原菌[8]。哈茨木霉菌可以防治及缓解黄瓜的根腐病和线虫病[9]、马铃薯黑痣病[10]、烟草青枯病[11]等植物病害。木霉菌还具有促进植物的生长发育,改善植物生理代谢等作用[12-15]。近年来发现,利用木霉菌与植物的互作关系可以增强植物对微生物胁迫的抗性[16]。李舒依等[17]研究发现,使用哈茨木霉菌可以提高盐胁迫下黄瓜种子的发芽率和胚芽长;付健等[18]研究发现,施用哈茨木霉菌可以提高盐碱土壤中玉米叶片光合色素含量,增强光合作用、抗氧化酶活性,从而缓解玉米盐碱抑制。关于哈茨木霉菌对植物重金属抗性作用相关研究较少,待进一步探索。
草地早熟禾(Poapratensis)是一种多年生草坪草,其生命力十分顽强,能适应各种不同的生长环境,无论潮湿还是干旱;气温偏高或是寒冷地区,只要水分供应充足就能生长[19]。因此,草地早熟禾常作为草坪草广泛运用在园林绿化中。本试验使用草地早熟禾作为材料,研究哈茨木霉菌对不同浓度镉处理下草地早熟禾种子萌发、生长状况及生理效应的影响,探讨哈茨木霉菌提高植物抗镉能力的生理机制,并为植物修复镉污染土地的应用提供理论支撑。
1 材料和方法
1.1 材料
草地早熟禾(Poapratensis),购于哈尔滨市和平绿化有限公司。哈茨木霉菌(T.harzianum)Th62菌株由东北林业大学园林学院张荣沭博士提供。Cd胁迫处理使用氯化镉试剂。
1.2 方法
1.2.1 种子萌发试验
哈茨木霉浓度筛选。选取颗粒饱满、大小均匀的草地早熟禾种子先用95%乙醇浸泡2 min,再用20%次氯酸钠溶液消毒20 min,蒸馏水冲洗3~4遍后浸种2 h。取5个小烧杯放入足够种子,将浓度分别为0(CK)、102、104、106和108mL-1的哈茨木霉菌悬浮液2 mL依次加入烧杯,浸种3 h。随后将种子转移到具有双层滤纸发芽床的直径9 cm的培养皿中,每皿50粒种子,加蒸馏水3 mL,设置3个重复。光照14 h、25 ℃,黑暗10 h、15 ℃。28 d为一个发芽期,每天记录种子萌发数量。本试验发芽结束时间以5 d内没有新种子发芽为准。结果显示,106mL-1处理后种子萌发率最高(表1),因此,选该浓度作为后续种子萌发试验的哈茨木霉处理浓度。

表1 不同浓度哈茨木霉菌处理对草地早熟禾的种子萌发率的影响
用106mL-1哈茨木霉菌悬浮液(T处理)和蒸馏水(C)分别浸种,处理方法同上。处理后的种子移入具双层滤纸发芽床的培养皿,皿中分别加入浓度为0(CK)、100、200、300和400 μmol·L-1的CdCl2溶液,每个处理3次重复,每个重复50粒种子。每日记录发芽及生长情况,10 d计算发芽势、28 d计算发芽率,并测量胚芽和胚根长度,计算出发芽指数及活力指数。
发芽势=(10 d内发芽种子数/供试种子数)×100%;
发芽率=(24 d内发芽种子数/供试种子数)×100%;
发芽指数=Σ(在不同时间的发芽数/发芽日数);
活力指数=发芽指数×胚芽长度。
1.2.2 幼苗生长、镉积累及生理特性试验
幼苗盆栽试验:将配置好的浓度为108mL-1(根据预试验结果确定)的哈茨木霉提取液混入无菌土中,每500 g无菌土加入200 mL哈茨木霉菌提取液,将草地早熟禾种子播种于土中(T处理),同时设置土壤中加200 mL蒸馏水为对照组(C)。28 d后补加一次哈茨木霉菌提取液,再培养28 d后进行Cd2+处理。Cd2+浓度为0(CK)、10、40、80和100 mg·L-1,每处理3个重复,每盆100 mL Cd溶液。处理7和14 d后分别取样,测定草地早熟禾幼苗的株高、生物量、各项生理指标和镉含量。脯氨酸含量参照高俊凤的测定方法;可溶性蛋白含量的测定参照G250法;丙二醛含量的测定参照硫代巴比妥酸法;超氧化物歧化酶(SOD)的测定采用氮蓝四唑法;过氧化物酶(POD)的测定采用愈创木酚法;过氧化氢(CAT)的测定采用过氧化氢法;镉含量的测定采用原子吸收分光光度计进行测定。
1.3 数据处理与分析
用Excel 2019整理试验数据、绘制图表。
2 结果与分析
2.1 哈茨木霉对镉胁迫下草地早熟禾种子萌发和幼苗生长的影响
由表2可知,随着Cd2+浓度增大,种子发芽率、发芽势降低,且发芽势受到抑制程度大于发芽率。哈茨木霉菌浸种(T)组相对于未浸种(C)组,发芽率和发芽势、发芽指数和活力指数在各浓度Cd2+处理下均升高。
由图1可知,随着Cd2+浓度的增加,草地早熟禾种子的胚芽长、胚根长均显著降低,且T组相对于C组,种子的胚芽长在各Cd2+处理下均升高,胚根长度除400 μmol·L-1浓度差异不明显外,其他组同样升高。
2.2 哈茨木霉对镉处理下草地早熟禾植株生长的影响
由表3可知,随着Cd2+浓度的升高,幼苗的株高呈降低趋势,且T组相对于C组,株高在不同浓度Cd2+处理下均升高。Cd2+处理7 d,各Cd2+浓度处理下T组株高比C组分别增长了5.29、7.87、9.22、8.14和8.41 cm;Cd2+处理14 d,T组比C组分别增长了5.84、8.28、8.79、8.75和7.32 cm。

表2 哈茨木霉对Cd2+胁迫下草地早熟禾种子萌发的影响

柱上无相同小写字母表示组间差异显著(P<0.05)。图2~4同。图1 哈茨木霉对镉胁迫草地早熟禾胚芽、胚根长度的影响

表3 哈茨木霉菌对Cd2+胁迫下草地早熟禾株高及生长量的影响
幼苗地上部分生物量的变化规律总体看,一定浓度的Cd2+对其有促进作用,且在低浓度下,T组数值高于C组。地下部分生物量随Cd2+浓度升高而降低。各Cd2+处理下T组比C组分别增长了6.7、10.8、19.4、19.9和38.3 mg;处理14 d时,各Cd2+处理下T组比C组分别增长了13.6、18.4、13.2、12.4和8.7 mg。
2.3 哈茨木霉菌对镉处理下草地早熟禾生理特性的影响
由表4可知,随着Cd2+浓度的升高,叶绿素a、叶绿素b和总叶绿素含量均呈降低趋势。Cd2+处理7 d,各Cd2+浓度处理下T组叶绿素a比C组分别增加了0.04、0.07、0.11、0.19和0.20 mg·g-1;叶绿素b含量增加了0.11、0.31、0.34、0.32和0.33 mg·g-1;叶绿素总含量分别增加了0.15、0.39、0.44、0.50和0.53 mg·g-1。Cd2+处理14 d,各浓度Cd2+处理下,T组相较于C组叶绿素a含量增加,但叶绿素b和总叶绿素含量减少。
脯氨酸和可溶性蛋白是植物在逆境下产生的主要渗透调节物质。由图2可知,随着Cd2+浓度的升高,脯氨酸含量增加,且T组脯氨酸含量高于C组。Cd2+处理7 d,各Cd2+浓度处理下T组脯氨酸含量比C组分别增加了79.91、104.37、119.95、115.71和119.87 μg·g-1;Cd2+处理 14 d,T组较C组分别增加了47.15、39.00、41.39、89.02和92.54 μg·g-1。
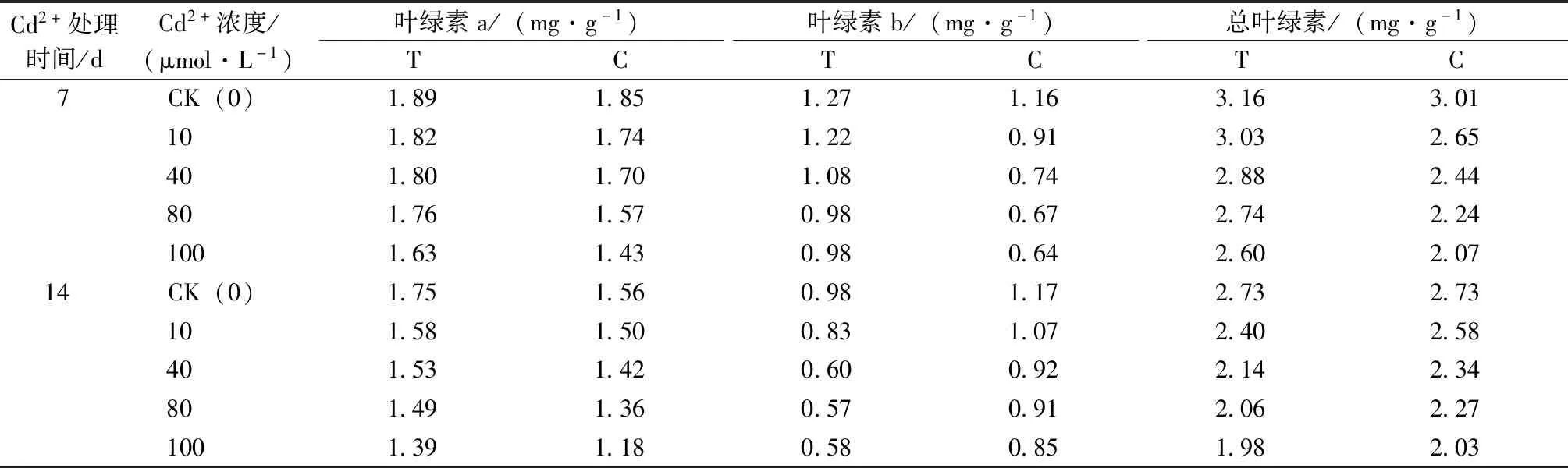
表4 哈茨木霉菌对镉胁迫下草地早熟禾叶绿素含量的影响
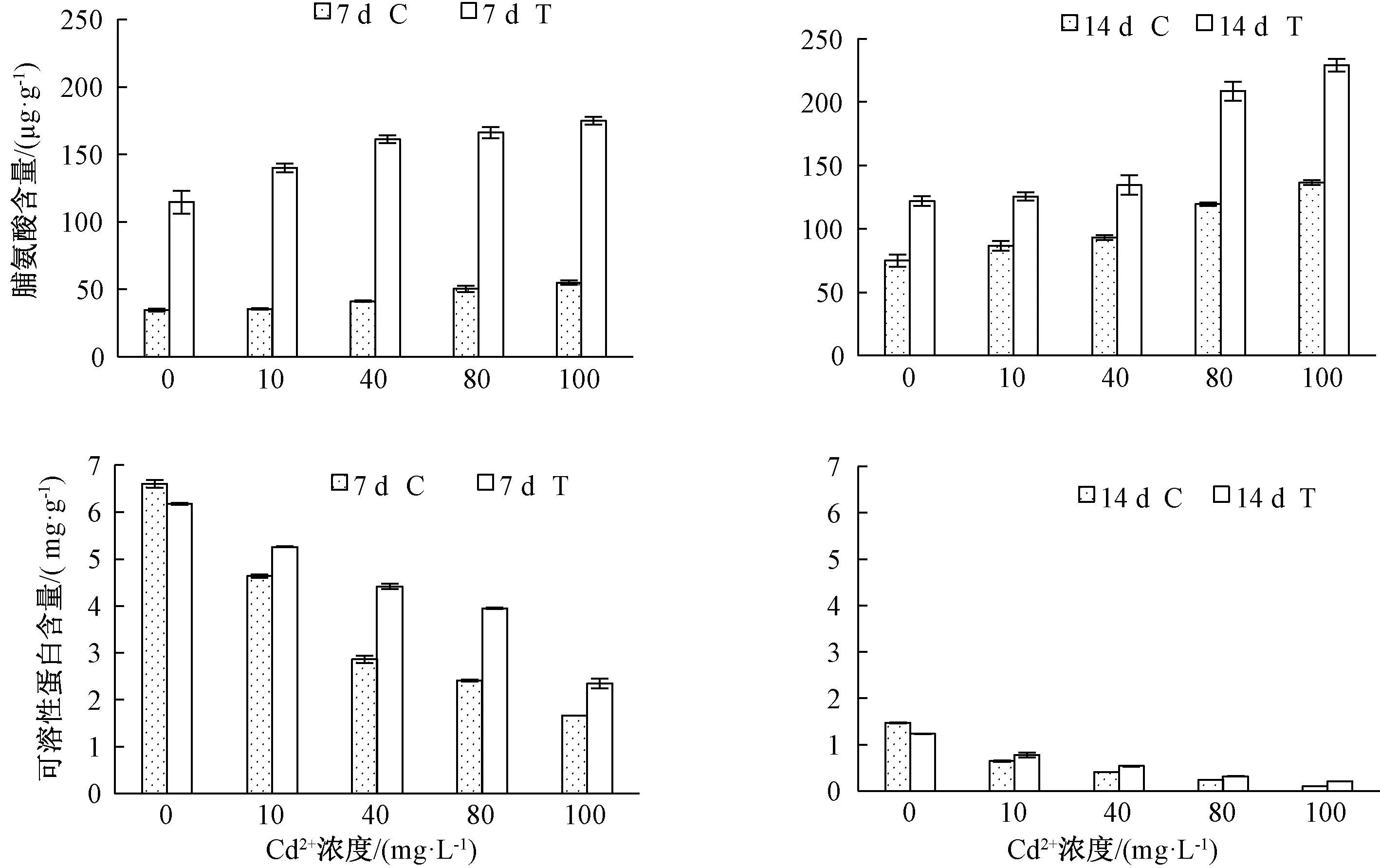
图2 哈茨木霉对镉胁迫下草地早熟禾脯氨酸、可溶性蛋白含量的影响
随着Cd2+浓度的升高,可溶性蛋白含量减少。在无Cd2+处理时,T组低于C组,在各组Cd2+处理时,T组高于C组。Cd2+处理7 d,各Cd2+浓度处理下T组可溶性蛋白含量比C组分别增加了0.62、1.56、1.54和0.69 mg·g-1;Cd2+处理14 d,T组较C组分别增加了0.13、0.12、0.08和0.11 mg·g-1。
丙二醛(MDA)为膜脂过氧化产生的有毒物质。图3结果显示,Cd2+浓度越大,叶片MDA含量越大,且T组均低于C组。Cd2+处理7 d,各Cd2+浓度处理下T组MDA含量比C组分别减少了0.45、0.11、0.43、0.65和0.83 mg·g-1;Cd2+处理14 d,T组较C组分别减少了0.81、0.90、0.64、1.58和1.43 mg·g-1。
在逆境下,植物体内可产生大量的活性氧,而保护酶系统可以起到清除活性氧的作用。由图4可知,随着Cd2+浓度的增加,SOD和CAT活性先升高后降低,POD活性呈逐渐升高。在Cd2+处理7 d后,除Cd2+浓度为80 mg·L-1外,其他各Cd2+浓度下T组SOD活性较C组升高;Cd2+处理14 d后,比较T与C处理组SOD活性在低Cd2+浓度下升高,高Cd2+浓度下差异不明显。Cd2+处理7 d和14 d后,在各Cd2+浓度下,T组POD活性较C组POD活性均降低。Cd2+处理7 d后,40 mg·L-1Cd2+处理下CAT活性最大,且Cd2+浓度为80和100 mg·L-1时,T组较C组分别减少1.67、3.93 U·g-1;Cd2+处理14 d,T组较C组CAT活性增加。
2.4 哈茨木霉菌对镉处理下草地早熟禾镉转运的影响
由表5可知,草地早熟禾地上部所吸收的Cd含量随着Cd2+处理浓度的增加和处理时间的延长而增加,且T组含量均高于C组。Cd2+处理7 d,各Cd2+浓度处理下T组Cd含量比C组分别增加了1.24、0.78、2.71、2.10和4.26 mg·kg-1;Cd2+处理14 d,除40 mg·L-1Cd2+处理T组与C组无明显差异外,其他组处理下T组比C组分别增加了0.32、1.62、1.37和2.46 mg·kg-1。

图3 哈茨木霉菌对镉胁迫下草地早熟禾MDA含量的影响

图4 哈茨木霉菌对镉胁迫下草地早熟禾抗氧化系统活性的影响
C组草地早熟禾地下部所吸收的Cd含量随着Cd2+处理浓度的增加和处理时间的延长而增加;T组地下部所吸收的Cd含量随着Cd2+处理浓度的增加先增加后减少;随处理时间的延长而增加。100 mg·L-1Cd2+处理时,T组地下部Cd含量低于C组。镉胁迫会降低早熟禾Cd转移系数,施加哈茨木霉菌会增加早熟禾Cd转移系数。处理7 d时,0、10、40、80和100 mg·L-1Cd2+处理时T组比C组增加了0.25、0.04、0.09、0.02和0.09;处理14 d时,各组Cd2+处理时T组比C组增加了0.19、0.05、0.01、0.01和0.05。
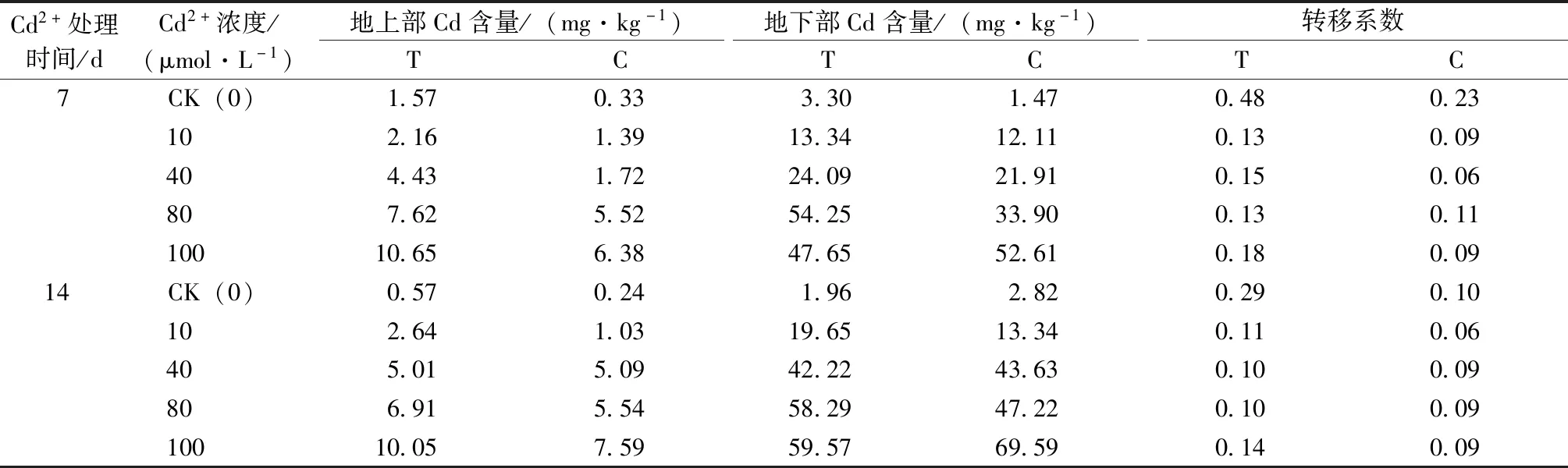
表5 哈茨木霉对镉胁迫下草地早熟禾Cd含量的影响
3 小结与讨论
哈茨木霉作为一种重要的生防菌,对多种植物具有促生效果[20]。众多研究结果显示,经低浓度哈茨木霉处理后种子的发芽率有不同程度的提高,浓度升高时促进作用变弱[21-22]。木霉菌可以产生一些生物调节类物质,如赤霉素、生长激素等,赤霉素可以增加种子萌发过程中可溶性糖和可溶性蛋白的含量,从而促进生长[23]。有研究还发现,使用康宁霉素浸种可减少种子内部营养物质外渗,保留种子内部营养充足,供种子萌发过程使用[24]。本试验结果表明,浓度为106mL-1的哈茨木霉菌对促进草地早熟禾种子的萌发和生长最为有效,浓度超过106mL-1对发芽率的促进作用减弱,这与前人研究结果一致。在镉胁迫下,随着Cd2+浓度的升高,草地早熟禾种子的发芽率呈显著降低的趋势,且T组较C组的发芽率、发芽势、发芽指数、活力指数均增加,说明哈茨木霉菌浸种可以在一定程度上减轻Cd抑制的种子萌发过程。试验还表明,草地早熟禾种子的胚芽、胚根受镉胁迫的抑制,随着Cd2+浓度的升高抑制作用增强,且对胚根的抑制作用大于胚芽。而哈茨木霉菌可以起到缓解镉胁迫对草地早熟禾的毒害作用。
镉对植物形态和生理代谢具有严重的毒害作用[25]。诸多研究表明,镉对植物的影响呈“低促高抑”的现象,低浓度可以促进植物的生长,但当镉含量过高时,会抑制植物生长,严重时还会出现植株矮小萎黄、根系死亡、生物量减少等症状[26-27]。本研究结果显示,随着镉胁迫强度及胁迫时间增加,草地早熟禾植株株高呈下降趋势,相较于C组,T组可以缓解镉胁迫的抑制作用,使株高增加。镉胁迫对草地早熟禾植株地上生物量、地下生物量的影响有差异,T组在低浓度Cd2+处理时地上生物量高于C组。地下部分生物量随着Cd2+浓度增加而降低,且T组较C组增高。从上述结果可以看出,哈茨木霉菌对镉胁迫下草地早熟禾的生长有一定的促进作用。
光合作用是植物转化光能为化学能的基础生理活动,叶绿素具有捕捉和传递光能的作用[28],是光合作用的关键色素,其含量多少在一定程度上标志着植物生长能力的强弱[29]。本研究结果显示,处理7 d后,随着Cd2+浓度升高草地早熟禾植株的叶绿素各指标降低,T组含量高于C组;随着处理时间增长到14 d时,T组的叶绿素b含量降低并低于C组,说明哈茨木霉菌可以缓解镉胁迫短时间内造成的草地早熟禾植株的光合色素的下降。
细胞渗透调节物质是一类具有维持细胞平衡作用的物质,主要包括游离脯氨酸、可溶性蛋白、可溶性糖等[30]。逆境条件会使植物中脯氨酸含量显著增加,当逆境持续加重时,会致使植株内脯氨酸含量减少,严重时还会致使植株死亡,所以通过植物体内游离脯氨酸含量的变化情况观察植物的抗逆性强弱[31]。可溶性蛋白的积累可以防止植物细胞水分的流失,进而增强植物抗逆性;可溶性蛋白还可以充当各种生理代谢过程所需的酶类,植物体在逆境条件下,也会消耗大量的可溶性蛋白来激活抗氧化系统,使植物抵御伤害[32]。本试验显示,草地早熟禾植株脯氨酸含量随着Cd2+浓度的增加和胁迫时间的增长而增加,且T组高于C组;可溶性蛋白含量则随Cd2+浓度增加和胁迫时间增长而减少,且T组同样高于C组。说明哈茨木霉菌可以增强镉胁迫下草地早熟禾的渗透调节能力。
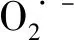
植物对镉的吸收能力是有限度的,当外界镉浓度超过植物本身能承受的浓度时,植物将减少对镉的吸收[37]。卜和申等[38]发现,镉污染土壤中分离的哈茨木霉菌可以通过细胞壁和细胞质吸附Cd2+,产生IAA类物质,促进植物生长,并可提高植物吸收Cd2+的能力,使Cd2+向地上部分转移,从而降低土壤中的镉含量。本试验结果显示,草地早熟禾地上部分Cd2+含量随着Cd2+浓度的增加而增加,且处理时间越长积累的越多,T组积累量比C组增多。地下部分Cd2+含量随着Cd2+浓度增加大致呈增加趋势,且0~80 mg·L-1时,T组Cd含量较C组增高。转移系数可以反映植物根部从土壤中吸收重金属后从地下向上转运能力的强弱,转移系数越大,转运能力越强[39]。施加哈茨木霉菌可以增加Cd转移系数,处理7 d时,Cd2+浓度为40 mg·L-1时转移系数增长最多,处理14 d时,Cd2+浓度为10 mg·L-1时转移系数增长最多。这说明哈茨木霉在较低Cd2+浓度范围内可以增强草地早熟禾对镉的积累和转运,并且通过多种生理效应保证植株正常生长,这对于增强镉污染土壤的植物修复效果具有重要的意义。
本试验结果表明,哈茨木霉浸种能够减轻镉胁迫对草地早熟禾种子萌发及幼苗生长的抑制作用;哈茨木霉也可以通过提高Cd转运能力,减少Cd对植株根系的直接伤害,缓解镉胁迫对草地早熟禾生长的抑制作用,并可以通过降低叶绿素破坏、减少MDA积累,增强抗氧化酶(SOD、POD、CAT)活性,增加渗透调节物质来提高草地早熟禾对Cd的抗性。
猜你喜欢
中老年保健(2022年3期)2022-11-21
天津农业科学(2022年6期)2022-07-17
热带农业科学(2022年4期)2022-05-18
当代水产(2022年1期)2022-04-26
干旱地区农业研究(2021年4期)2021-08-11
湖南农业大学学报(自然科学版)(2021年3期)2021-07-02
安徽化工(2021年3期)2021-05-29
棉花学报(2020年3期)2020-08-08
科学与财富(2019年15期)2019-10-21
中国钱币(2015年6期)2015-11-18
