
基于生物信息学构建带状疱疹神经痛miRNA-mRNA 调控网络*
2022-09-16 06:04莫泳锋吴逸伦周增华何睿林蒋宗滨
中国疼痛医学杂志 2022年8期
带状疱疹 (herpes zoster, HZ) 是由潜伏在背根神经节、半月神经节或自主神经节中的水痘-带状疱疹病毒 (varicella zoster virus, VZV) 在机体疲劳、精神压力过大、全身性疾病、创伤等危险因素刺激下重新激活导致感觉神经受损,并伴有神经性疼痛的感染性皮肤病
。据报道,世界上有15%~30%的人一生中至少会患一次带状疱疹
,13.3%~42.2%的带状疱疹病人会出现带状疱疹后神经痛 (postherpetic neuralgia, PHN),表现为典型的神经病理性疼痛,且疼痛持续时间长,可从数月到数年,平均持续时间为3.3 年
。PHN 作为HZ 最常见和最严重的合并症
,往往由于长期的疼痛和伴随的焦虑、抑郁心理问题,严重降低病人的生活质量。虽然近些年对PHN 的分子机制研究有了较深的研究,其可能涉及背根神经节炎症、神经炎症、感觉神经根轴突和髓鞘缺失、纤维化和中枢敏化、外周敏化、神经去传导,但其具体发病机制尚未完全阐明,致使临床上有效或彻底根治PHN 的治疗方法较少。因此,探究带状疱疹神经痛发生发展的分子机制,将有利于我们对疾病本身的认识,同时给临床诊治提供指导。
基因测序和生物信息学分析已经被广泛应用于疾病的分子机制研究。微小RNA (microRNA,miRNA)作为一种内源性、低分子量、单链结构的非编码RNA,在人体液中含量丰富,通过结合转录mRNA 的特异性靶点而影响下游基因的表达,参与帕金森病、癌症、炎症性肠病、心血管疾病、糖尿病等多种疾病的病理生理过程和发病机制,并在其预防和治疗方面有着重要临床意义
。有研究表明
,miRNA 在神经系统的感觉器官中高度表达,如背根神经节和脊髓背角,且通过调节炎症因子、离子通道和调控免疫平衡参与神经病理性疼痛的发生发展,特别是水痘-带状疱疹病毒 (varicella zoster virus, VZV) 编码的多种miRNA 可能参与调节宿主细胞的感染,并与PHN 的发生过程密切相关。目前的研究大多关注于个别miRNA 对mRNA的调节作用,且大多数miRNA 与mRNA 之间的相互作用关系仍不清楚,特别是miRNAs 介导的带状疱疹神经痛miRNA-mRNA 调控网络的构建尚未见有研究报道。因此,有必要构建带状疱疹神经痛miRNA-mRNA 调控网络,探讨miRNA 与mRNA之间在带状疱疹神经痛方面的相互作用关系。本研究以miRNAs 为基础,利用生物信息学方法构建miRNA-mRNA 调控网络,筛选关键的miRNA-mRNA调控关系对,探讨miRNA 与其靶基因在带状疱疹相关神经痛分子调控机制中的作用,为靶向治疗带状疱疹相关神经痛提供理论参考和科学依据。
方 法
1.材料
采用美国国立生物技术信息中心(NCBI)的基因表达综合数据库(GEO 数据库https://www.ncbi.nlm.nih.gov/geo/)获取VZV 感染的人神经母细胞瘤细胞 (SH-SY5Y) 转录组测序数据(GSE141932)。其包含3 例未感染VZV 样本和6 例感染VZV 样本的mRNA 表达数据,其中3 例未感染VZV 样本纳入对照组,6 例感染VZV 样本纳入实验组,对两组测序数据通过平台文件转换为标准基因名称,构建mRNA 表达矩阵和归一化整理。
本文提出基于LabVIEW的模拟调制系统设计。用LabVIEW实现模拟调制并将各系统进行程序管理,生成适合用户使用的界面。由于LabVIEW可以用来创建通用的程序,在测试、测量和自动化领域具有更大的优势,它提供了大量的工具与函数用于数据采集、分析、显示和存储。虚拟仪器与传统仪器相比,可以在数分钟内完成一套完整的从仪器连接、数据采集到分析、显示和存储的自动化测试测量系统。
当马老师给我描述李之恒家里多穷,他是多么努力,多么有出息的时候,我走神了。如果现在叫我去努力回忆李之恒是个什么样的人,我眼前浮现的形象却是我大学一个室友的模样。
2.差异基因的筛选
根据《环境空气质量标准(AQI)》(GB3095—2012)(环境保护部,2012)和相关研究(顾康康等,2018),将 PM2.5日平均浓度<35 µg·m-3、35~75µg·m-3和>75 µg·m-3分别定义为轻度、中度和重度污染浓度水平。同时,将O3-8 h-max(O3最大8 h平均浓度值)按照浓度水平进行分级,也分为轻度、中度和重度3个等级,分别对应100~160 µg·m-3、160~200 µg·m-3和>200 µg·m-3(杨书申等,2016)。污染程度划分等级后,可用于计算不同级别 PM2.5和O3各占季节总日数的比例,其公式为:
在雁北和吕梁山区,人们赋予吃莜面栲栳栳以“牢靠”“和睦”等美好象征。每逢老人寿诞、小孩满月或逢节待客,多以此进餐。山区有些人家婚配嫁娶时,新郎新娘也要吃,意谓夫妻白头到老。年终岁末时更要吃,以祈全家和睦、人运亨通。栲栳栳的制作方法并不复杂,但入口却筋道爽口,有一股浓浓的莜面清香,可沾酱料食用,也可以与其它蔬菜肉类同炒,风味独特。
3. miRNA-mRNA 网络构建
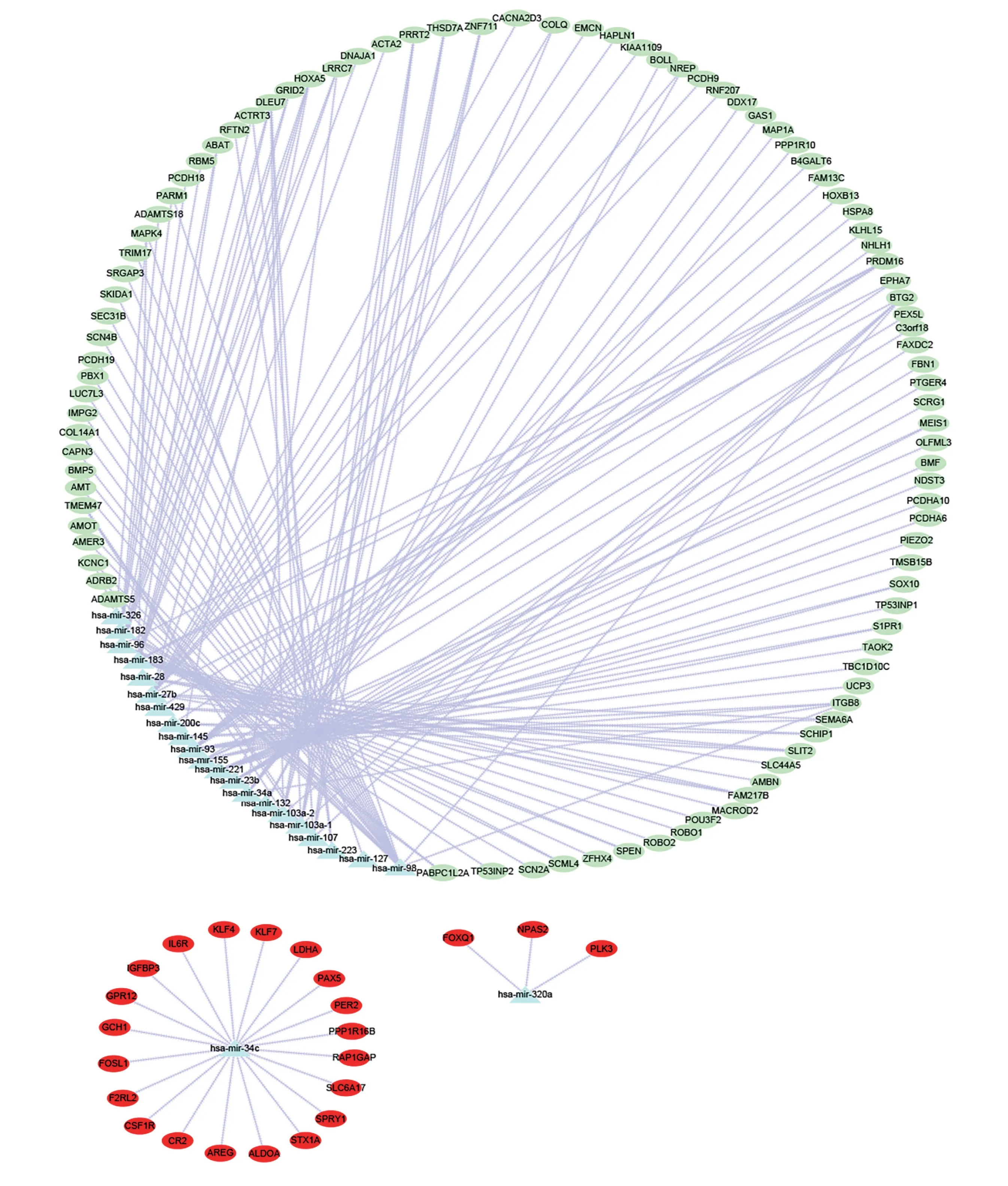
通过miRNet 在线数据库,共筛选出46 个miRNAs。预测miRNAs 下游靶基因,并与DEGs取交集得到123 个靶基因,构建miRNA-mRNA 可视化调控网络(见图3)。其中,miRNA-34c 和miRNA-320a 参与IL6R、FOSL1、ALDOA、PAX5、STX1A、LDHA、PLK3、NPAS2等靶基因的表达上调,miR-107、miR-183、miR-98、mir-182、mir-27b 参与PARM1、MAPK4、PBX1、SLIT2、SCHIP1、HOXA5、ACTRT3 等靶基因的表达下调。
4. miRNA 上游转录因子预测和GO 分析
FunRich 软件提供转录因子、途径、生物过程、细胞成分、分子功能和蛋白质相互作用等注释,利用该软件对miRNA-mRNA 网络中的miRNAs 进行上游转录因子(transcription factor, TF)预测和GO 注释分析,阈值
< 0.05。
利用R 软件limma_3.46.0 包对构建的mRNA表达矩阵整理分析,得到差异表达基因 (differentially expressed genes, DEGs),绘制火山图并使用pheatmap_1.0.12 包绘制DEGs 热图。DEGs 筛选条件:①取mRNA 差异表达变化倍数 (fold change, FC),|log
FC| > 1;②adjusted
< 0.05。
5.靶基因功能富集分析
对构建miRNA-mRNA 网络的靶基因ID 转换后,通过clusterProfiler 包和pathview 包进行GO 和KEGG 通路富集分析,阈值
< 0.05。
6.蛋白质互作网络建立与分析
虽然MicroRNA 已经被证实与PHN 的发生机制及预后密切相关,并在PHN 病人的皮肤病变组织和血清中发现了几种异常表达的miRNAs
,但目前大多数miRNAs 与mRNA 的相互作用关系尚未完全阐明,特别是带状疱疹神经痛相关的miRNA-mRNA调控网络的构建未见有研究报道,构建带状疱疹相关神经痛miRNA-mRNA 调控网络可为深入研究其分子发病机制提供重要的理论参考和科学依据。
结 果
1.差异基因的筛选
共筛选到差异表达基因 (differentially expressed genes, DEGs) 1250 个,其中719 个基因表达上调,531个基因表达下调,绘制火山图和DEGs的热图(见图1、2),可见对照组和实验组之间有明显差异。


2. miRNA-mRNA 网络构建
miRNet 数据库是一个高质量的数据预测平台(https://www.mirnet.ca),它同时收录miRTarBase、TarBase、miRanda 等多个数据库信息
。本研究通过miRNet 在线数据库筛选神经病理性疼痛、带状疱疹后神经痛、疼痛、慢性疼痛、慢性炎性疼痛、神经炎症相关的miRNAs。通过FunRich_3.1.3软件进行miRNAs 下游靶基因预测。预测得到的miRNAs 下游靶基因与DEGs 取交集,获得miRNA-mRNA 关系对,并利用Cytoscape_3.8.1 软件构建miRNA-mRNA 可视化调控网络。
3. miRNA 上游转录因子预测和GO 分析
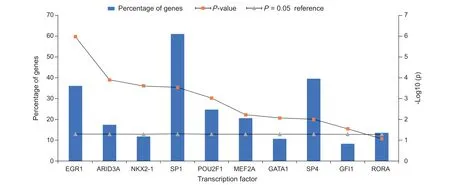
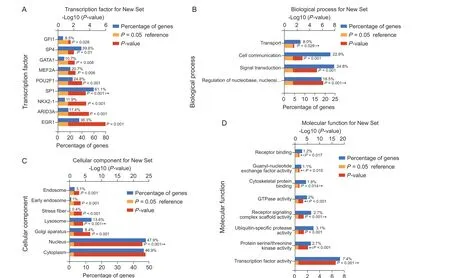
共得到195 个上游转录因子,其中前9 个最具有分析意义(见图4),包括EGR1、ARID3A、NKX2-1、SP1、POU2F1、MEF2A、GATA1、SP4、GFI1。对195个上游转录因子GO分析发现(见图5),其主要富集在核苷酸代谢调节、信号传导、丝氨酸/苏氨酸蛋白激酶活性、泛素特异性蛋白酶活性、信号转导复合体活性、GTP 酶活性、鸟苷酸交换因子活性等生物功能和分子功能。
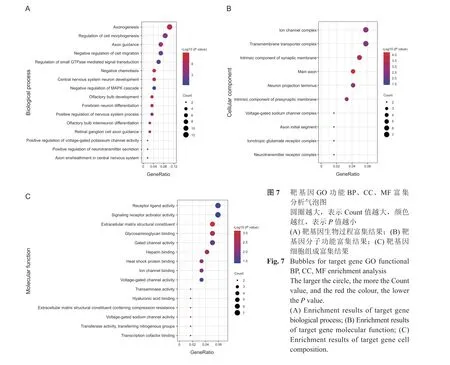
4.靶基因GO 功能注释和KEGG 分析
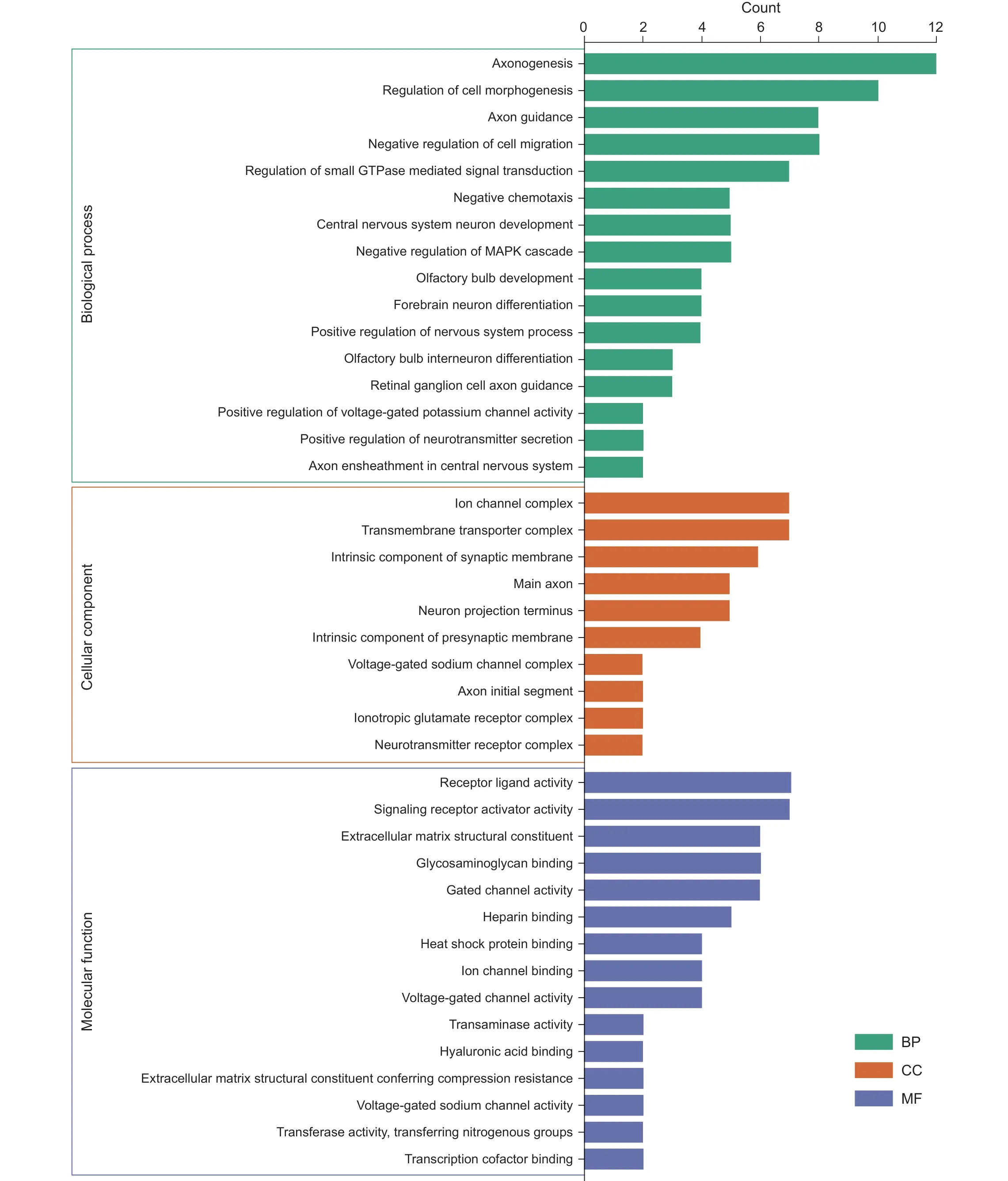
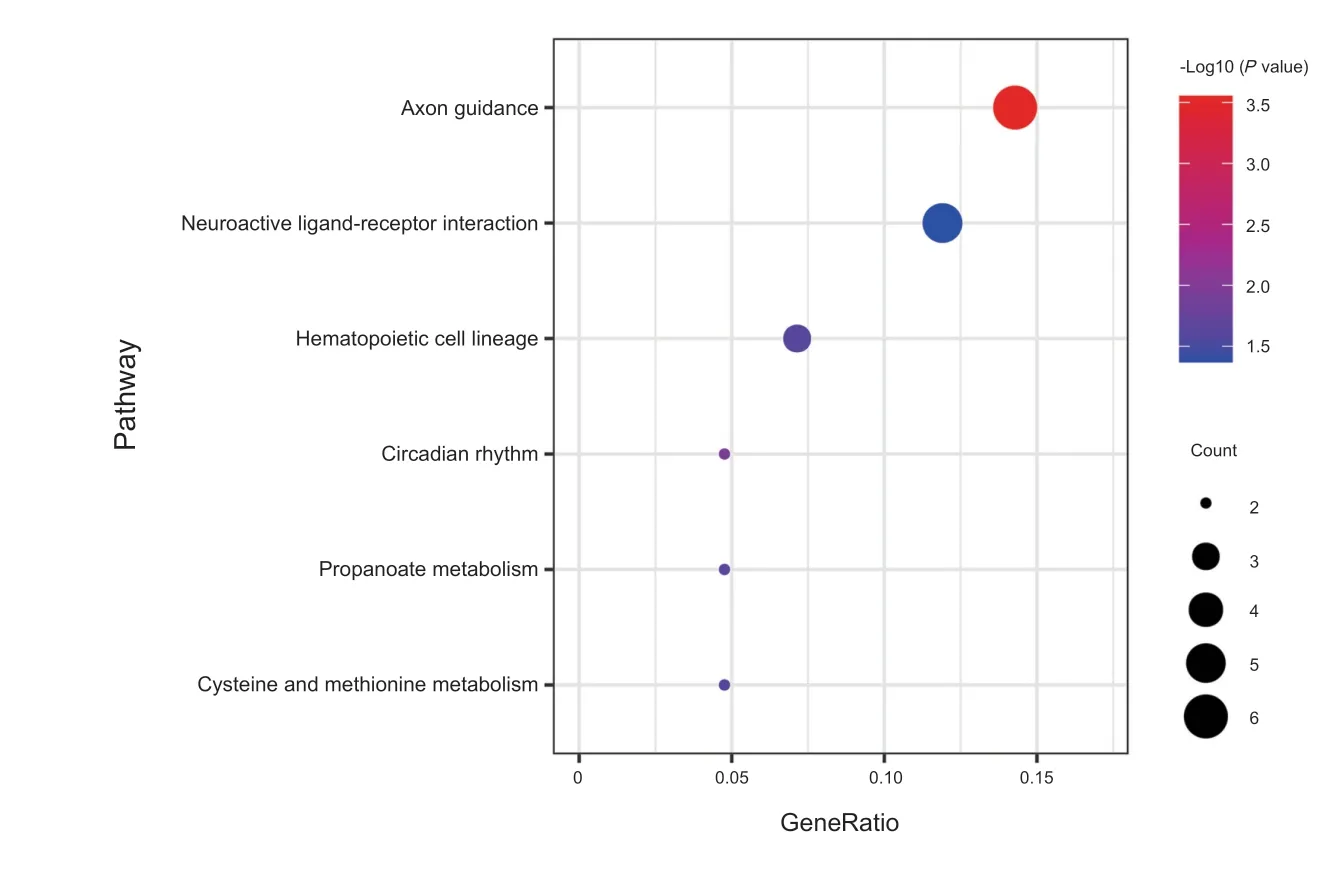
靶基因GO 功能分析发现(见图6、7),在生物过程方面,主要富集在负趋化作用、嗅球发育、轴突生成、嗅球中间神经元分化、中枢神经系统神经元发育、视网膜神经节细胞轴突引导等。在细胞组成方面,主要富集在跨膜转运蛋白复合体、轴突起始部、突触前膜的组成、神经元映射末端、离子通道复合体、电压门控钠通道复合物等。在分子功能方面,主要富集在细胞外基质结构组成、热休克蛋白结合、电压门控钠通道活性、转移酶活性、离子通道结合、转录辅助因子结合等。KEGG 分析发现(见图8),靶基因主要参与轴突引导、昼夜节律、神经活性配体-受体相互作用等信号通路。本研究发现,靶基因SLIT2、ROBO2、ROBO1 共同参与轴突引导信号通路,其中ROBO2、ROBO1 与miR-183,SLIT2 与miR-27b、miR-429、miR-200c关系密切。靶基因PER2、NPAS2 共同参与昼夜节律信号通路,其中PER2 与miR-34c,NPAS2 与miR-320a关系密切。靶基因F2RL2、GRID2、ADRB2、S1PR1、PTGER4 共同参与神经活性配体-受体相互作用信号通路,其中F2RL2 与miR-34c,GRID2 与miR-96 和miR-183,ADRB2 与miR-98,S1PR1 与miR-93 和miR-155,PTGER4 与miR-23b 关系密切。
对于选择终止妊娠的HIV感染孕妇,应给予安全的人工终止妊娠服务,应尽早手术,以减少并发症的发生。对于选择继续妊娠的孕妇,应给予优质的孕期保健、产后母乳喂养等问题的咨询,并采取相应的干预措施。
5. PPI 网络建立与分析
本研究利用生物信息学分析方法,对从GEO 数据库获取的VZV 感染神经母细胞瘤细胞 (SH-SY5Y)转录组测序数据成功构建了miRNA-mRNA 可视化调控网络。对该网络进一步分析发现,hsa-mir-34c参与上调靶基因较多,hsa-mir-98、hsa-mir-221、hsa-mir-93、hsa-mir-183、hsa-mir-107、hsa-mir-182、hsa-mir-27b 参与下调靶基因较多。
hsa-mir-34c 已被发现与癌症、炎症性肠病等疾病的发病机制有关,且有研究表明,miR-34c 在大脑中的表达下调可能参与帕金森氏病的发病机制
。与急性期带状疱疹病人相比,带状疱疹后神经痛病人血清中miR-34c-5p、miR-107 表达水平明显升高,特别是miR-107 在调节分化神经元,维持特定miRNA 稳态中起着重要作用,且miRNA 可能参与神经性疼痛有关的免疫应答
。有研究表明
,miR-183-5p 和miR-221-3p 可以通过外泌体输送调节靶基因,促进血管生成。miR-183 的过表达可引起电压门控钠通道Na
1.3 和脑源性神经营养因子(brain-derived neurotrophic factor, BDNF)的下调,进而减轻脊神经结扎(spinal nerve ligation, SNL)诱导的机械性痛觉过敏
。miR-98 可靶向作用于caspase-3,抑制内皮细胞凋亡,调节Aβ 的产生,进而治疗阿尔茨海默病。同时,其可通过抑制Fas和caspase-3,抑制心肌梗死相关的细胞凋亡
。miR-98 过 表 达 通 过 减 少IL-6、IL-1β、TNF-α 和COX-2 产生,抑制神经炎症反应,调节HMGA2和STAT3 表达而减轻坐骨神经慢性压迫损伤模型(chronic constriction injury, CCI)大鼠的神经病理性疼痛
。miR-182 与SORT1 共同调控体内外血管平滑肌细胞的钙化,参与肺动脉高压血管的重塑,miR-27b 通过调节血管内皮生长因子 C (VEGF-C)的表达促进血管生成和皮肤修复
。综上所述,miR-34c、miR-107、miR-183、miR-98、mir-182、mir-27b 在神经病理性疼痛方面的部分研究结果与本研究的分析结果一致。因此,我们推测调控网络中的mir-221、mir-93 等其他miRNAs 在带状疱疹相关神经痛的发病机制中可能同样起着重要作用,特别是miR-183、mir-221、miR-182 和miR-27b 可能与带状疱疹神经痛相关的血管病变机制关系密切。
讨 论
PHN 是带状疱疹病人最常见的并发症之一,不少病人要经受多年的剧烈疼痛,严重的慢性疼痛给社会和个人带来沉重的负担,包括经济、社交、日常活动和生活质量下降,并逐渐成为一个公共卫生问题。虽然有研究认为PHN 的发生与外周敏化、中枢敏化、炎症和伤害性感受器的异位放电有关
,但其具体发病机制尚不清楚,致使临床上有效或彻底根治这个疾病的治疗方法较少。基因和蛋白表达的改变被不少学者认为是神经病理性疼痛发展和维持的重要特征
,同时,认识到miRNAs 在基因表达调控中起着关键作用,可通过抑制疼痛同源靶基因的表达而参与神经病理性疼痛的发生和发展
,且miRNAs 已被作为研究多种疾病预后的潜在标志物。
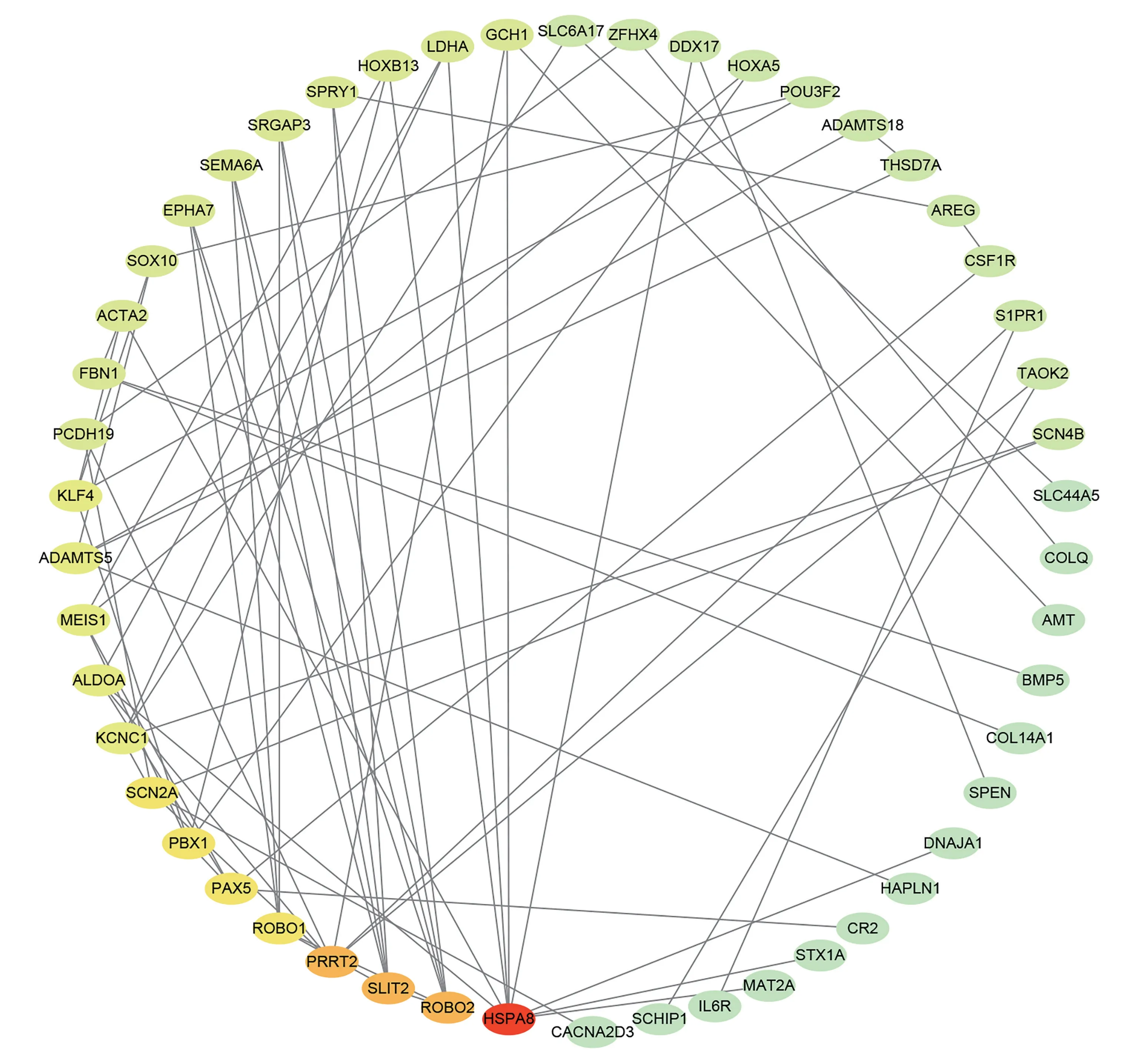
为了更好地了解靶基因所表达蛋白质之间的关系,使用String (https://string-db.org/) 数据库,在综合得分≥0.4 条件下建立靶基因蛋白质互作网络,并利用Cytoscape_3.8.1 软件对蛋白质互作网络图进一步调整,同时利用 “Cytohubba” 插件,通过最大团中心性MCC 算法筛选出前10 位的Hub 基因。
将靶基因通过String 数据库导出,并经Cytoscape 软件处理得到蛋白质互作网络(见图9),同时使用Cytohubba 插件,通过最大团中心性MCC算法筛选出前 10 位的Hub 基因,包括HSPA8、PRRT2、SLIT2、ROBO2、SCN2A、PAX5、PBX1、ROBO1、ALDOA、ADAMTS5。
研修班主要特色:主承办单位高度重视,组织落实到位;以行业需求为导向,完善课程体系;不断创新研修机制,优化研修环境;提升研修水平,培育领军人才。主要成效:聚集创新资源,推广科研新成果;强化责任意识,激发组织新活力;发挥辐射作用,提升品牌新高度。中国BIM高级研修班多期的成功举办促进了行业知识型、技术型、创新型高素质人才队伍建设,为产业持续创新发展提供了重要人才支撑,对探索有效的行业人才培养机制和模式、引领开创人才教育培训新格局起到积极的示范效果。
miRNA 的表达受转录因子调控,miRNA 亦可反过来影响转录因子的功能,两者共同参与mRNA转录和翻译,是细胞代谢的重要组成部分
。在本研究中,通过FunRich 软件预测miRNA-mRNA网络中的miRNA 上游转录因子,得到调控miRNA有显著意义的转录因子EGR-1、SP1、SP4、MEF2A等。EGR-1 参与细胞生长、突触传递和认知活动等不同类型的神经元反应,背根神经节和脊髓背角中的EGR-1 参与神经损伤诱导的神经病理性疼痛过程,同时,EGR-1 表达受miR-124-3p 调控
。转录因子SP4 主要在神经元中表达,参与信号传导和能量产生相关的生物过程,SP1 参与调控神经性疼痛钙通道α2δ-1 亚基的表达,诱导机械性痛觉过敏
。转录因子EGR-1、SP1、SP4、MEF2A可能通过干扰细胞生长、突触传递、细胞凋亡及钙通道等途径参与带状疱疹相关神经痛的病理生理过程,同时这些转录因子也可能成为PHN 的潜在治疗靶点。
靶基因GO 和KEGG 富集分析发现,这些靶基因主要富集于轴突生成、中枢神经系统神经元发育、热休克蛋白结合、转移酶活性、电压门控通道活性、转录辅助因子结合等生物过程和分子功能,并参与轴突引导、昼夜节律、神经活性配体-受体相互作用等信号通路。Slit2 作为一种分泌蛋白,与其受体 Robo1 结合,可激活细胞自噬,调节细胞分化,参与炎症过程
。有证据表明,miRNA 可能参与神经病理性疼痛相关生物钟基因的转录后调控,miR-183/96/182 在感觉神经系统中有重要作用,且miR-182 和miR-96 的表达水平具有明显的昼夜节律
。各种慢性疾病伴随的疼痛都存在昼夜节律,且神经病理性疼痛的昼夜变化节律有多种机制参与,如μ 阿片受体、β-内啡肽和纳洛酮之间的日常变化、与疼痛有关的钙通道及炎症调节因子
。
本研究构建的调控网络表明,靶基因SLIT2、ROBO2、ROBO1 共同参与轴突引导,PER2、NPAS2共同参与昼夜节律,F2RL2、GRID2、ADRB2、S1PR1、PTGER4 共同参与神经活性配体-受体相互作用信号通路。鉴于筛选的多种基因参与多种疾病的发生机制,因此,我们推测这些靶基因可能参与带状疱疹相关神经痛有关的轴突引导、昼夜节律、神经活性配体-受体相互作用信号通路的调控,特别是筛选得到的Hub 基因SLIT2、ROBO2、ROBO1,它们可能在带状疱疹相关神经痛的轴突引导信号通路机制中有重要作用。
尽管该研究成功构建了带状疱疹神经痛miRNA-mRNA 调控网络,筛选得到一些关键基因,并发现了miRNA 和mRNA 是通过多途径、多靶点参与调控带状疱疹相关神经痛的发病机制,但该研究尚有许多不足之处,如缺乏适当的实验来进一步支持该研究的结果,我们将在今后的研究中进行有针对性的实验验证,以更好地解释其作用机制。
综上所述,该研究构建的带状疱疹神经痛miRNA-mRNA 调控网络对阐明带状疱疹相关神经痛的分子发病机制提供了理论支持,为未来构建更有意义的带状疱疹相关神经痛临床预测模型,研发带状疱疹相关神经痛的基因靶向药物提供了新的生物学标识和重要参考。
利益冲突声明:作者声明本文无利益冲突。
[1] Harbecke R, Jensen NJ, Depledge DP,
. Recurrent herpes zoster in the Shingles Prevention Study: are second episodes caused by the same varicella-zoster virus strain?[J]. Vaccine, 2020, 38(2):150-157.
[2] Sullivan NL, Eberhardt CS, Wieland A,
. Understanding the immunology of the zostavax shingles vaccine[J]. Curr Opin Immunol, 2019, 59:25-30.
[3] 陈杨, 蒲勋, 肖智,等.带状疱疹及带状疱疹后神经痛病人受累神经分布特点[J]. 中国疼痛医学杂志,2022, 28(4):295-298.
[4] Leung J, Harpaz R, Molinari NA,
. Herpes zoster incidence among insured persons in the united states, 1993-2006: evaluation of impact of varicella vaccination[J]. Clin Infect Dis, 2011, 52(3):332-340.
[5] Oster G, Harding G, Dukes E,
. Pain, medication use, and health-related quality of life in older persons with postherpetic neuralgia: results from a populationbased survey[J]. J Pain, 2005, 6(6):356-363.
[6] Texakalidis P, Tora MS, Boulis NM. Neurosurgeons'armamentarium for the management of refractory postherpetic neuralgia: a systematic literature review[J].Stereotact Funct Neurosurg, 2019, 97(1):55-65.
[7] Schuster NM, Hsia-Kiung ME. Glossopharyngeal postherpetic neuralgia palliated with fluoroscopicguided nerve block: a case report[J]. Headache, 2018,58(1):154-156.
[8] 吴雨菲, 邹天浩, 杨东. 脊髓电刺激治疗带状疱疹神经痛的应用进展[J]. 中国疼痛医学杂志, 2022,28(2):134-138.
[9] Plotnikova OM, Skoblov MY. Efficiency of the miRNA-mRNA interaction prediction programs[J]. Mol Biol (Mosk), 2018, 52(3):543-554.
[10] Irmak MK, Erdem U, Kubar A. Antiviral activity of salivary microRNAs for ophthalmic herpes zoster[J].Theor Biol Med Model, 2012, 9:21.
[11] Chen T, Xue H, Lin R,
. MiR-34c and PlncRNA1 mediated the function of intestinal epithelial barrier by regulating tight junction proteins in inflammatory bowel disease[J]. Biochem Biophys Res Commun,2017, 486(1):6-13.
[12] Kabaria S, Choi DC, Chaudhuri AD,
. Inhibition of miR-34b and miR-34c enhances alpha-synuclein expression in Parkinson's disease[J]. FEBS Lett, 2015,589(3):319-325.
[13] Markus A, Golani L, Ojha NK,
. Varicella-zoster virus expresses multiple small noncoding RNAs[J]. J Virol, 2017, 91(24):e01710-17.
[14] Huang Y, Li X, Tao G,
. Comparing serum microRNA levels of acute herpes zoster patients with those of postherpetic neuralgia patients[J]. Medicine (Baltimore),2017, 96(8):e5997.
[15] 彭丹, 阎雪彬. microRNA 在神经病理性疼痛中的研究与进展[J]. 中国疼痛医学杂志, 2020, 26(1):58-60.
[16] Chang L, Zhou G, Soufan O,
. miRNet 2.0:network-based visual analytics for miRNA functional analysis and systems biology[J]. Nucleic Acids Res,2020, 48(W1):W244-W251.
[17] Lin CS, Lin YC, Lao HC,
. Interventional treatments for postherpetic neuralgia: a systematic review[J]. Pain Physician, 2019, 22(3):209-228.
[18] Wang J, Ma SH, Tao R,
. Gene expression profile changes in rat dorsal horn after sciatic nerve injury[J].Neurol Res, 2017, 39(2):176-182.
[19] 郭佳宝, 陈炳霖, 朱毅等. MicroRNA 在外周神经损伤所致神经病理性疼痛中的作用[J]. 中国疼痛医学杂志, 2019, 25(4):243-250.
[20] Cao S, Zhang D, Yuan J,
. Microrna and circular rna expression in affected skin of patients with postherpetic neuralgia[J]. J Pain Res, 2019, 12: 2905-2913.
[21] 徐海峰. 带状疱疹后遗神经痛与血清miRNA 表达的关系研究[D]. 沈阳:中国医科大学, 2018.
[22] Wang C, Ding M, Xia M,
. Afive-miRNA panel identified from a multicentric case-control study serves as a novel diagnostic tool for ethnically diverse nonsmall-cell lung cancer patients[J]. EBio Medicine,2015, 2(10):1377-1385.
[23] Shang A, Wang X, Gu C,
. Exosomal miR-183-5p promotes angiogenesis in colorectal cancer by regulation of FOXO1[J]. Aging (Albany NY), 2020,12(9):8352-8371.
[24] Wu XG, Zhou CF, Zhang YM,
. Cancer-derived exosomal miR-221-3p promotes angiogenesis by targeting THBS2 in cervical squamous cell carcinoma[J].Angiogenesis, 2019, 22(3):397-410.
[25] Lin CR, Chen KH, Yang CH,
. Intrathecal miR-183 delivery suppresses mechanical allodynia in mononeuropathic rats[J]. Eur J Neurosci, 2014, 39(10):1682-1689.
[26] Sun C, Liu H, Guo J,
. MicroRNA-98 negatively regulates myocardial infarction-induced apoptosis by down-regulating Fas and caspase-3[J]. Sci Rep, 2017,7(1):7460.
[27] Li Q, Li X, Wang L,
. miR-98-5p acts as a target for alzheimer's disease by regulating abeta production through modulating SNX6 expression[J]. J Mol Neurosci, 2016, 60(4):413-420.
[28] Li HW, Meng Y, Xie Q,
. miR-98 protects endothelial cells against hypoxia/reoxygenation induced-apoptosis by targeting caspase-3[J]. Biochem Biophys Res Commun, 2015, 467(3):595-601.
[29] Zhang Y, Su Z, An LJ,
. miR-98 acts as an inhibitor in chronic constriction injury-induced neuropathic pain via downregulation of high-mobility group AT-hook 2[J]. J Cell Biochem, 2019, 120(6):10363-10369.
[30] Zhong L, Fu K, Xiao WM,
. Overexpression of miR-98 attenuates neuropathic pain development via targeting STAT3 in CCI rat models[J]. J Cell Biochem,2018. doi:10.1002/jcb.28076.
[31] Sun L, Lin P, Chen Y,
. miR-182-3p/Myadm contribute to pulmonary artery hypertension vascular remodeling via a KLF4/p21-dependent mechanism[J].Theranostics, 2020, 10(12):5581-5599.
[32] Liu J, Sun F, Wang X,
. miR-27b promotes angiogenesis and skin repair in scalded rats through regulating VEGF-C expression[J]. Lasers Med Sci,2020, 35(7):1577-1588.
[33] Mullany LE, Herrick JS, Wolff RK,
. MicroRNAtranscription factor interactions and their combined effect on target gene expression in colon cancer cases[J]. Genes Chromosomes Cancer, 2018, 57(4):192-202.
[34] Jiang M, Zhang X, Wang X,
. MicroRNA-124-3p attenuates the development of nerve injury-induced neuropathic pain by targeting early growth response 1 in the dorsal root ganglia and spinal dorsal horn[J]. J Neurochem, 2021, 158(4):928-942.
[35] Duclot F, Kabbaj M. The role of early growth response 1 (EGR1) in brain plasticity and neuropsychiatric disorders[J]. Front Behav Neurosci, 2017, 11:35.
[36] Gomez K, Sandoval A, Barragan-Iglesias P,
.Transcription factor Sp1 regulates the expression of calcium channel alpha2delta-1 subunit in neuropathic pain[J]. Neuroscience, 2019, 412:207-215.
[37] Martinez-Hernandez E, Gonzalez-Ramirez R, Sandoval A,
. Isolation and characterization of the 5' upstream region of the human voltage-gated Ca(2+) channel channel α 2δ-1 auxiliary subunit gene: promoter analysis and regulation by transcription factor Sp1[J].Pflugers Arch, 2013, 465(6):819-828.
[38] Nair B, Johar K, Priya A,
. Specificity protein 4(Sp4) transcriptionally regulates inhibitory GABAergic receptors in neurons[J]. Biochim Biophys Acta, 2016,1863(1):1-9.
[39] Priya A, Johar K, Nair B,
. Specificity protein 4(Sp4) regulates the transcription of AMPA receptor subunit GluA2 (Gria2)[J]. Biochim Biophys Acta,2014, 1843(6):1196-206.
[40] Xie J, Li L, Deng S,
. Slit2/Robo1 mitigates dssinduced ulcerative colitis by activating autophagy in intestinal stem cell[J]. Int J Biol Sci, 2020, 16(11):1876-1887.
[41] Morioka N, Saeki M, Sugimoto T,
. Downregulation of the spinal dorsal horn clock gene Per1 expression leads to mechanical hypersensitivity via c-jun Nterminal kinase and CCL2 production in mice[J]. Mol Cell Neurosci, 2016, 72:72-83.
[42] Shende VR, Neuendorff N, Earnest DJ. Role of miR-142-3p in the post-transcriptional regulation of the clock gene Bmal1 in the mouse SCN[J]. PLoS One,2013, 8(6):e65300.
[43] Kim HK, Lee SY, Koike N,
. Circadian regulation of chemotherapy-induced peripheral neuropathic pain and the underlying transcriptomic landscape[J]. Sci Rep, 2020, 10(1):13844.
[44] Takada T, Yamashita A, Date A,
. Changes in the circadian rhythm of mRNA expression for micro-opioid receptors in the periaqueductal gray under a neuropathic pain-like state[J]. Synapse, 2013, 67(5):216-223.
猜你喜欢
医学概论(2022年3期)2022-04-24
中国典型病例大全(2022年7期)2022-04-22
保健与生活(2021年23期)2021-12-12
新传奇(2020年44期)2020-11-23
红领巾·探索(2020年10期)2020-10-26
科普童话·神秘大侦探(2017年4期)2017-04-06
人生十六七(2016年9期)2016-09-22
妇女生活(2015年12期)2015-09-10
第二课堂(课外活动版)(2009年12期)2009-03-18
中国美容医学(2004年3期)2004-09-17
