
甜菜谷胱甘肽S-转移酶基因家族鉴定及在镉胁迫下的响应分析
2022-10-12 06:40钟鑫爱孟诗琪周婉婷刘大丽
植物研究 2022年5期
钟鑫爱 孟诗琪 周婉婷 姚 琦 张 琼 兴 旺 刘大丽
(1. 国家甜菜种质中期库,黑龙江大学,哈尔滨 150080;2. 黑龙江大学省高校甜菜遗传育种重点实验室/现代农业与生态环境学院,哈尔滨 150080;3. 黑龙江大学省高校生化与分子生物学重点实验室/生命科学学院,哈尔滨 150080)
谷胱甘肽S-转移酶()是一种由超基因家族编码的古老的多样性蛋白家族,在生物机体的免疫防御系统中发挥着解毒和抗氧化等作用。由于最初发现这种酶在植物中应对除草剂的过程中起解毒作用,因此也被称为解毒酶。植物中的主要功能为响应逆境条件胁迫,如化学药品胁迫、重金属逆境胁迫、高温、高盐、低温等。
在植物中GSTs 可分为Tau、Phi、Theta、Zeta、DHAR、Lambda、TCHQD 与EF18 个类型的亚家族,除了Lambda 与DHAR 亚家族,GSTs 在溶液中均以二聚体蛋白的形式存在,与谷胱甘肽(Glutathione,GSH)共价结合后形成不相同的三肽疏水亲电底物,在细胞中络合活性氧物质,调控细胞氧化还原状态,在次生代谢产物的解毒过程中发挥重要的作用。在各个GSTs 亚家族中,Phi 与Tau亚家族为植物所特有,并且数量最多。目前国内外对植物基因家族已有大量研究,研究表明在 拟南芥()、大 豆()、玉米()、水稻()、杨树(spp.)中分别存在25~81 个基因,并对其功能进行分析发现,植物的基因能够对不同的非生物胁迫做出响应,如除草剂、重金属胁迫、盐胁迫和干旱胁迫等。
重金属是植物生长过程中重要的微量元素,但当含量超过一定阙值时会导致生物活性分子的生理功能受到抑制,进而导致植物重金属中毒。镉是一种环境中常见的重金属,植物受到镉胁迫的影响主要表现为“低促高抑”,低浓度的镉可以适当促进幼苗生长,而高浓度的镉胁迫会导致植物生长停止。镉的长期积累,会转移到食物链,对粮食安全和人类健康构成重大威胁。而当遇到重金属如镉等胁迫时,生物体内的GSTs 可催化GSH 结合过氧化物或络合结合自由态Cd,也能够通过解除活性亲电子复合体来抵御一系列逆境胁迫的伤害,并在抵御活性氧物质对细胞的毒害中发挥重要作用。目前已有许多植物响应镉胁迫的报道,如番茄()、苎麻()、水稻、白菜()和拟南芥等。Kilili 等在番茄的研究中,鉴定出5 个Tau 家族参与了氧化应激反应所涉及到的催化与调节功能网络。在不同的苎麻品种中,部分苎麻基因表现出对镉胁迫的积极响应。水稻植株在镉胁迫下,通过蛋白质互作提高了还原性GSH 的含量,GSTs 蛋白质复合体会产生特异性的表达,有效降低了镉对水稻植株的毒害作用。在白菜中,的大量表达使部分白菜品种的Cd积累量和Cd耐受性升高。研究表明,拟南芥 中 的、、基因参与有毒物质的代谢过程,其中、参与了镉离子的胁迫应答。此外,在烟草()、苜蓿()和 芦 苇()等植物中过表达基因,均证明基因与镉耐受相关。
在研究土壤重金属污染修复的方法中,传统的修复方法如化学修复法、物理修复法都具有成本高、耗能大的问题。而利用生物代谢治理重金属污染的生物修复技术则具有经济有效,并且能够防止二次污染的特点,是具有较大前景的修复方法。甜菜()是我国乃至世界重要的糖料作物之一,同时由于其体内含有大量的糖类碳水化合物,因此可以生产工业乙醇,作为一类新兴的能源作物,其具有较大的经济效益。在面对土壤中重金属污染时,甜菜适应非生物胁迫的能力很强,能通过根际富集重金属,促进重金属的分解,进而可以降低土壤中重金属的污染程度。但目前关于甜菜耐受重金属胁迫的分子机制的研究较为少见。实验室前期研究发现,基因(LOC104898671)与镉逆境胁迫存在着一定的应答关系。将其转入大肠杆菌()中,该基因可以有效的直接或间接地降低Cd 对细胞的毒害,进而提高了大肠杆菌的重金属耐受性。在对基因进行不同浓度的Cd胁迫,均能诱导甜菜基因表达上调,在Cd浓度为0.5 mmol·L时该基因的相对表达量达到最高值。这证实基因可以提高甜菜在镉胁迫下的耐受性,但是其在镉胁迫下精确的作用机制还有待研究,针对其家族成员的全基因组鉴定尚且鲜见报道。因此,本研究利用同源比对法,在甜菜基因组中挖掘基因家族成员,并对基因家族成员的保守基序、基因结构、进化关系与顺式作用元件进行深入的分析,结合对镉胁迫应答的转录组数据,筛选与镉胁迫相关的,并分析它们的转录表达模式。进而为研究基因家族的生物学功能提供基础信息,为提高甜菜对重金属逆境的耐受性和种质创新提供新途径。
1 材料与方法
1.1 植物材料
甜菜材料取自国家甜菜种质资源中期库。将大小均一的甜菜种子,用体积分数75%的乙醇消毒后,取蒸馏水反复冲洗,再用2%福美双浸泡过夜,用蒸馏水冲洗干净。将消毒后的种子均匀排列在含有蛭石的培养槽中,放在含有光照的温室条件下培养。至甜菜种子萌发后,取2片子叶期的甜菜幼苗培养于Hoagland 营养液中,培养至6 片真叶期。用浓度为0.5 mmol·L的CdCl溶液处理甜菜幼苗6 h,以正常条件下生长的甜菜幼苗为对照,每个处理3 次重复,分别取甜菜地上部及地下部组织进行转录组测序。
1.2 不同物种基因组数据库的获得及甜菜BvGSTs基因家族成员的鉴定
在Ensemble 网站(http:plants.ensemble.org/index.html)下载甜菜、拟南芥全基因组序列、蛋白质组数据及基因组注释文件。借助隐马尔科夫模型(PF00043,PF02798)搜索甜菜蛋白质组中的GSTs蛋白序列,将冗余去除后,在Pfam(http//pfam.xfam.org)数据库及CDD(http:www.ncbi.nlm.nih.gov/Structure/cdd/wrpsb.cgi)数据库中将所有候选基因进行结构域的确认。
1.3 BvGSTs蛋白的理化性质分析
利用Expasy-ProtParam tool 在线分析工具(http://web.expasy.org/protparam/)获得氨基酸数、等电点(pI)、相对分子质量、不稳定系数等数据,整理获得相关的理化性质。利用在线软件Cell-PLoc 2.0(http:\www.csbio.sjtu.edu.cn/bioinf/Cell-PLoc-2)对基因进行亚细胞定位预测。
1.4 BvGSTs基因家族系统发育树的构建
利用MEGA X 软件中的Clustal W 功能对甜菜基因家族进化分类,根据52个甜菜和53 个拟南芥的氨基酸序列进行比对,采用邻近法(neighbour-joining method,NJ)构建系统进化树,其中Bootstrap method 值设定为1 000,其他参数设为默认值。
1.5 BvGSTs 基因家族的基因结构和和保守基序分析及染色体定位
从甜菜基因组的GFF3 注释文件中获取基因家族成员的外显子、内含子结构信息及染色体的位置信息,利用在线网站MEME(http:memesuite.org/tools/meme)对蛋白质保守基序进行分析。通过TBtools 软件对基因结构,保守基序、染色体位置进行可视化,保守基序数目设置为9,其他参数设为默认值。
1.6 BvGSTs基因家族顺势作用元件分析
利用TBtools软件提取与重金属镉胁迫相关的52 个成员起始密码子上游2 000 bp 的基因序列,通过PlantCARE(http://bioinformatics.psb.ugent.be/webtools/plantcare/html/)在线软件进行顺式作用元件预测,再用TBtools 软件与R 软件将顺势作用元件预测结果可视化。
1.7 镉胁迫下甜菜BvGSTs基因家族的差异表达分析
对提取的甜菜总RNA 进行高通量测序,对获取的家族基因在镉胁迫下的表达量进行差异表达分析。根据基因在地上部和地下部中的log(FoldChange)值,利用TBtools 软件进行聚类分析。
1.8 镉胁迫下BvGSTs基因表达GO分析
利用转录组测序结果获得镉胁迫下响应的甜菜基因的GO 条目,并运用在线网站Hiplot(https://hiplot.com.cn)对其GO 功能富集分析结果进行可视化。
1.9 qRT-PCR荧光定量分析
利用Trizol试剂盒提取样品总RNA,反转录成cDNA。根据目的基因序列,分别设计定量PCR 引物(见表1),使用SuperReal PreMix Plus(SYBR Green)荧光定量试剂盒进行qRT-PCR,检测每份样品的目的基因和内参基因Ct值,3次重复。利用2相对表达量进行分析。
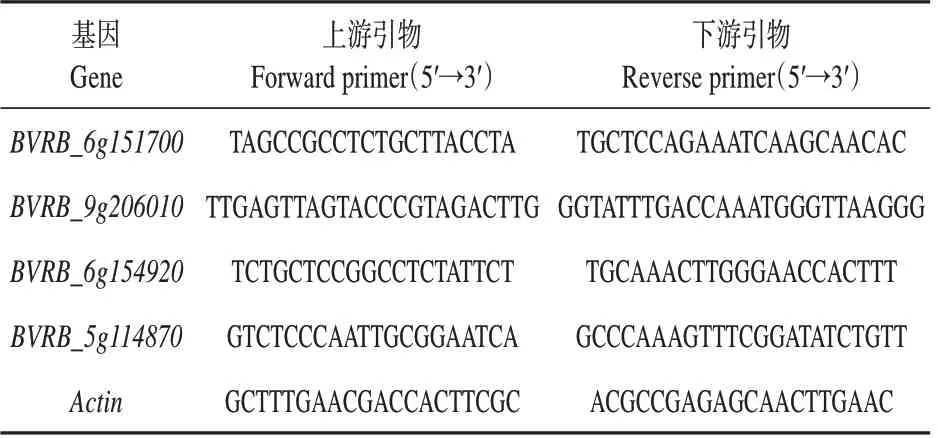
表1 荧光定量引物及序列Table 1 The primers and sequences for qRT-PCR
2 结果与分析
2.1 甜菜BvGSTs基因家族的筛选与理化性质
在Pfam 网站中利用序列号PF00043 和PF02798(GST 保守域)鉴定基因。在Ensemble 网站中下载甜菜全基因组蛋白序列。通过隐马尔科夫模型(PF00043,PF02798)搜索甜菜蛋白质组中的GSTs 蛋白序列。利用Pfam 和CDD 验证甜菜的保守结构域,最终在甜菜基因组中鉴定出52个家族成员。
如表2所示,52个基因编码的氨基酸在212aa()~240aa(),推测相对分子质量为17~28 kDa;pI 为4.72~9.65,其中47 个成员的pI 小于7,偏酸性;其余5 个成员的pI 大于7,偏碱性。在甜菜基因家族成员中41%的蛋白不稳定系数小于40,为稳定蛋白,其余的为不稳定蛋白;且大部分的定位于细胞质,仅基因位于细胞核。
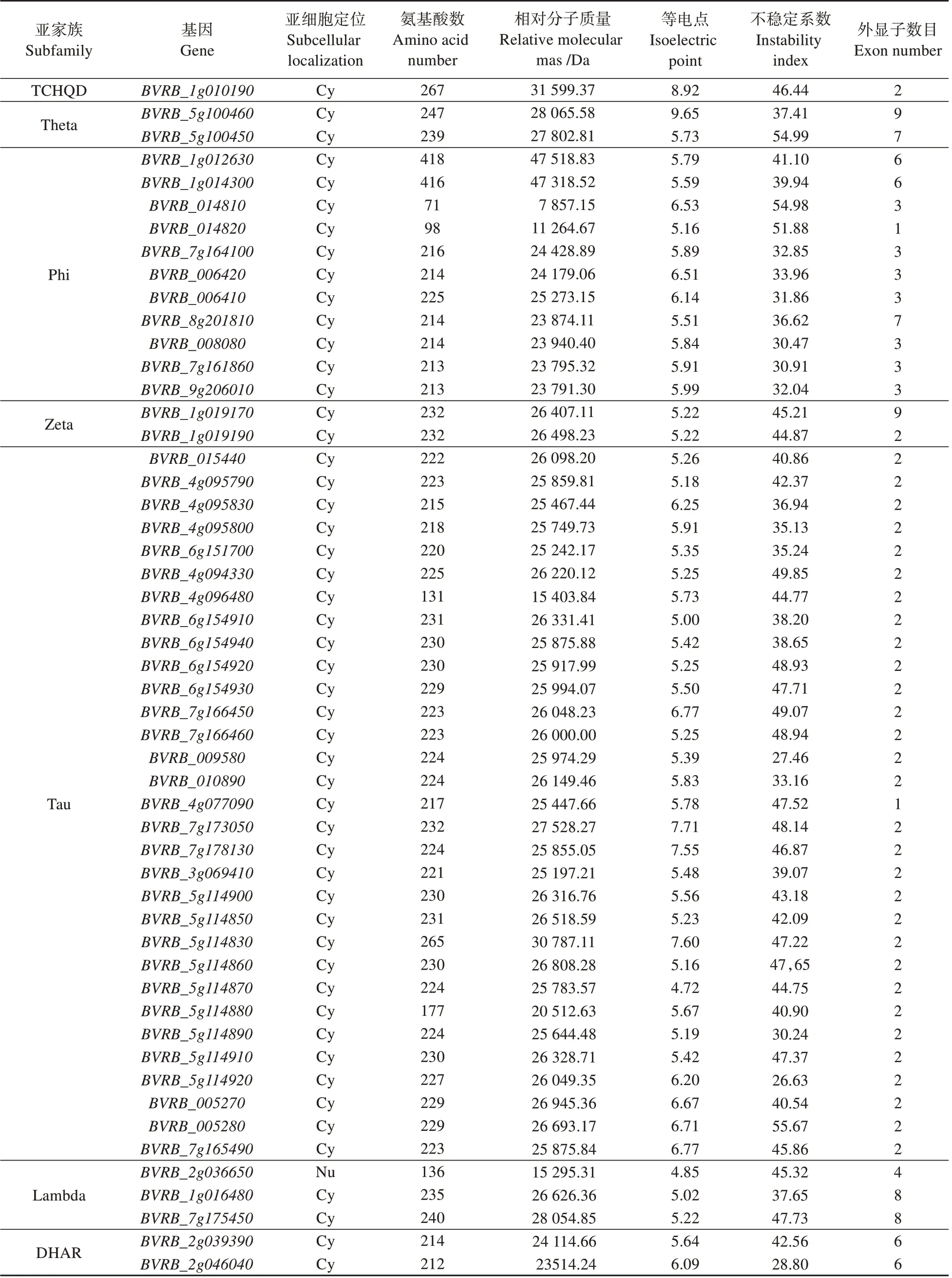
表2 甜菜BvGSTs基因家族的基本信息Table 2 The basic information of BvGSTs gene family in Beta vulgaris L.
2.2 甜菜BvGSTs基因家族的系统发育
利用52个甜菜BvGSTs蛋白序列和53个拟南芥AtGSTs蛋白序列构建系统发育树,利用MEGA X软件的Clustal W 功能构建系统发育树(见图1)。根据系统发育树的分枝和保守序列可看出,甜菜与拟南芥的GSTs 聚类为一个大分枝。甜菜GSTs可分为7 类:DHAR、Lambda、Phi、Tau、TCHQD、Theta 和Zeta。各个亚家族中,Tau 类亚家族是甜菜中较大的亚家族,数量有31 个;Phi 类为11 个、DHAR 类2 个、Lambda 亚家族有3 个、TCHQD 类1个、Zeta类2个以及Theta类2个。
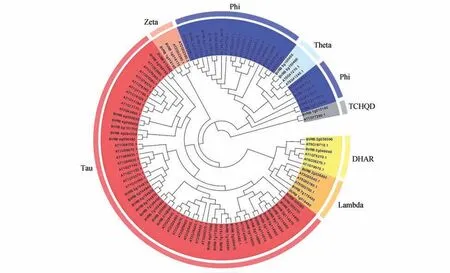
图1 甜菜(BV)、拟南芥(AT)家族进化树分析Fig.1 Phylogenetic tree analysis of Beta vulgaris(BV)and Arabidopsis thaliana(AT)
2.3 甜菜BvGSTs家族基因结构和保守基序及顺式作用元件
利用MEME 在线工具对甜菜52 个成员的保守域进行分析,并根据甜菜全基因组注释文件GFF 和甜菜家族基因CDS 序列信息进行基因结构分析,通过TBtools进行可视化(见图2)。结果表明,Tau 亚家族成员大都为2 个外显子;Phi亚家族外显子大都为3个,极个别基因的外显子为6~7个;Theta家族的和Zeta 家族的外显子均为9 个;DHAR、Lambda亚家族外显子为6个左右。在甜菜基因家族成员中,大部分都有5′和3′端翻译 区(untranslated region,UTR),其 中的3′端非翻译区长度较长,将近2 500 bp;而在这些成员中,没有5′和3′端UTR。
如图2 所示,在基因家族成员中,共鉴定出9个保守基序,并且不同的家族成员含有的基序为1~7个。大部分基因家族成员都包含motif 2,说明motif 2 在基因家族中较为保守。在52 个成员中可看出各个亚家族中的motif 排列相对相似,而不同亚家族之间的motif排列不同。Tau亚家族成员均含有motif 8、motif 9、motif 4、motif 3,且排列顺序相同;Phi 亚家族大部分含有motif 7、motif 4、motif 8,其中motif 8为Phi家族特有。

图2 甜菜BvGSTs基因家族系统进化树、基因结构和保守基序分析A.进化树;B.基因结构;C.保守基序;刻度标尺表示基因长度(bp)和蛋白序列长度(aa)Fig.2 Analysis of phylogenetic tree,gene structure and conserved motif elements of BvGSTs in Beta vulgarisA.Phylogentic;B.Gene structure;C.Conservative motif;Scale markers repersent gene length(bp)and protein sequence length(aa)
利用PlantCARE 在线软件预测顺式作用元件结果(见图3),可发现所有成员都有光反应、脱落酸反应、生长发育元干旱响应元件及MYB结合位点。可以推测,甜菜基因家族可能响应激素调节和非生物胁迫,并且MYB 可能是调控转录表达调控的转录因子之一。

图3 甜菜BvGSTs基因家族顺势作用元件Fig.3 Putative cis-acting element analysis of BvGSTs gene family in Beta vulgaris
2.4 甜菜BvGSTs基因的染色体定位
52 个基因中的30 个基因在甜菜的8条染色上分布,但其余22 个基因定位在scaffold上,未定位到染色体上,仍需要进行探索(见图4)。甜菜染色体上的各个基因分布密度不同,在5 号染色体上分布最多,为11 个,并集中于40~50 Mb;6号染色体上的多位于50~60 Mb;7号染色体上基因相对分布均匀,其中和位 于10~20 Mb,位于20~30 Mb,其他5 个基因位置在30~50 Mb;其他染色体均在不同位置分布了数量不等的。相对于各个染色体的长度,6、5、2、8、1 号染色体上各成员多集中于下部,9 和4号染色体上的成员分布下部。7号染色上各成员的分布较均匀。利用MCScan X 分析,发现基因家族中存在4 处串联基因重复,多集中于Tau亚家族和Phi 亚家族,形成多个基因簇,其中5号染色体出现2处串联复制基因簇。

图4 甜菜BvGSTs基因家族染色体分布与基因复制Fig.4 Chromosome distribution and gene replication of BvGSTs gene family in Beta vulgaris L.
2.5 甜菜体内响应镉胁迫的BvGSTs 基因表达GO功能富集分析
为了更好地探索基因在镉胁迫下的分子机制,对52 个基因进行了GO 功能富集分析(见图5),共得到了36 个GO 条目。从Value 值来看,谷胱甘肽转移酶活性、转移酶活性、转移烷基或芳基(甲基除外)基团、次级代谢、对毒性物质的反应、毒素代谢、毒素分解代谢过程和解毒作用这8 个GO 功能最为显著,综合来看,在甜菜镉胁迫下可能发挥着重要的解毒作用。

图5 甜菜BvGSTs家族基因在镉胁迫下的GO富集Fig.5 GO enrichment analysis of BvGSTs family genes in sugar beet under cadmium stress
2.6 镉胁迫下甜菜BvGSTs基因家族成员的表达特性
基于转录组测序结果发现,鉴定出的全部成员均与镉胁迫相关。如图6 所示,甜菜地下部中的33 个基因在转录过程中受到镉胁迫的正向调控,19 个基因呈下调表达趋势。在甜菜地上部中,、、和均显著的受到了镉胁迫的正向调控,镉胁迫较为明显的抑制了、、及的转录表达。
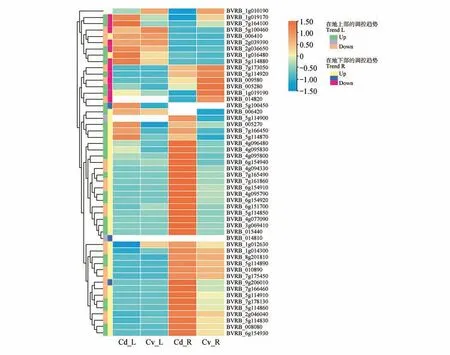
图6 甜菜BvGSTs家族基因在镉胁迫下的差异表达Cd_L 和Cd_R.甜菜BvGSTs 基因在镉胁迫下,地上部与地下部cDNA 文库中的表达量;Cv_L 和Cv_R.甜菜BvGSTs 基因在正常生长条件下,地上部与地下部cDNA文库中的表达量Fig.6 Differential expression of BvGSTs in Beta vulgaris under cadmium stressCd_L and Cd_R. Expression levels of BvGSTs gene in the leaf and the roof of sugarbeet’s cDNA libraries under cadmium stress;Cv_L and Cv_R.Expression levels of BvGSTs gene in the leaf and the roof of sugarbeet’s cDNA libraries under normal growth conditions
总体上看,镉胁迫下,在甜菜地上部与地下部均呈上调表达的共有17 个,其中和的差异表达较明显。在地上部和地下部呈下调共表达的基因为6 个(、、和)。结合系统进化树分析可以推断,在地下部,Tau 亚家族基因在镉胁迫时主要起正向调控作用,部分Zeta 亚家族基因在镉胁迫时起负向调控作用;在地上部,镉胁迫诱导Phi表达,抑制Tau表达。差异表达大的普遍分布于Tau 家族与Phi家族,在后续的镉胁迫研究中可以多关注Tau 家族与Phi 亚家族。
为了进一步确定基因家族在镉胁迫下的生物学功能,在转录组测序结果中筛选在地上部和地下部均差异表达显著的基因:(|log(FoldChange)|≥1;0.05)。将这4个基因利用qRT-PCR 技术分析它们在0.5 mmol·L的镉胁迫处理6 h 时,在地上部与地下部的表达量。如图7 所示,在镉胁迫下,在甜菜地上部组织中,呈 下调趋势,呈上调趋势;在甜菜地下部组织中,4 个差异表达显著的基因都呈上调趋势。在这4 个基因中,除为Phi 亚家族,其余3 个基因均来源于Tau 亚家族。qRT-PCR 的结果与转录组测序的结果相符合,也进一步证实了这4 个参与甜菜镉胁迫应答。

图7 qRT-PCR分析4个BvGSTs在镉胁迫下的转录表达特性Fig.7 Transcriptional expression characteristics of 4 BvGSTs by qRT-PCR under cadmium stress
3 讨论与结论
目前已在拟南芥、玉米、水稻、马铃薯()等物种中鉴定出基因,并发现它们广泛参与植物体中生长发育的解毒和抗氧化过程。GST 能够与谷胱甘肽(GSH)共价形成三肽疏水亲电底物,进而调控细胞氧化还原状态等,介导调控由于重金属逆境所引起的基因表达。甜菜适应非生物胁迫的能力较强,在重金属土壤污染修复方面有较大的潜力。实验室前期对甜菜在镉胁迫下的转录组进行了测序分析,并初步探索了其中的2 个差异表达基因在镉胁迫下的功能。研究推测甜菜基因家族成员可能不同程度的参与到甜菜的镉胁迫耐受调控网络中。
为了全面地了解和掌握甜菜体内的基因家族成员,本研究基于甜菜全基因组数据共挖掘出52个基因,它们分属于7个亚家族,占高等植物基因家族的87.5%。利用Expasy 对基因家族理化性质进行了分析,发现90% 的基因偏酸性,59%为不稳定蛋白。通过亚细胞预测,可发现大部分的位于细胞质,少数的基因位于细胞核,这说明作为可溶性的,它们几乎都在细胞质中发挥功能。这与已有的研究中确定的的亚细胞位置相符。将筛选鉴定得到的甜菜GSTs 的52 个蛋白序列与模式植物拟南芥AtGSTs 的53 个蛋白序列进行系统建树,可初步将甜菜的GSTs分为了7个亚家族,分 别 为Tau、Phi、Theta、Zeta、Lambda、TCHQD、DHAR。Tau 家族及Phi 家族在植物中分布较广泛,与植物体中基因组分布特征结果相一致。在进化过程中,Phi是由Tau亚家族分化而来,逐渐成为一个新的亚家族,但在植物的抗逆特性、解毒作用等特性上仍有相似性。已经有研究报道证明拟南芥中的的过表达会促进抗氧化酶的活性,进而增强植物对非生物胁迫的耐受性;会调控种子萌发时的渗透耐受性。因此通过甜菜与拟南芥的系统发育分析,可以推测Tau 亚家族及Phi 亚家族中与与亲缘关系较近的基因或许在抗逆作用中也有着可待挖掘的潜力。每个亚家族中的的保守基序、基因结构和保守结构域是相似的,但不同亚家族之间有所不同。基因亚家族之间在结构上存在着特异性,这可能决定不同亚家族发挥不同生物学功能。在基因家族中,基因串联重复会对基因家族遗传进化起着重要作用,水稻基因家族的进化是通过基因复制,然后由于复制基因的亚功能化或新功能化而保留下来。染色体定位分析中,半数的位于scaffold,未定位于染色体上,说明对于基因的研究还仍需探索。其余30 个基因分别分布于甜菜的8条染色体上,其中5号、7号染色体上分布基因较多,并且存在两处串联复制基因簇,四处串联基因重复,均属于Tau 家族和Phi 家族,这可能也是导致Tau及Phi亚家族分化的一个原因。
在以往对于基因家族功能的研究中,可知基因家族成员能够响应多种生物与非生物的胁迫,如机械损伤、重金属等。在对顺式作用元件的分析中发现,基因家族成员能够响应多种环境应激,推测顺式作用元件的存在可能会影响在受到环境胁迫时转录表达。研究发现,()可以提高真菌对Cd 的耐受性;水稻和基因的大量表达在不影响酵母在正常生长条件下生长状态的情况下,可以通过较高的GST 活性提高菌体对重金属Cr(vI)的抗性。为了解基因家族成员在镉胁迫下的表达特性,本研究对转录组数据中的基因表达水平进行了差异显著性分析,并发现所有甜菜基因家族成员的转录表达均受到镉胁迫不同程度的调控。在甜菜应答镉胁迫的机制中,Tau亚家族在地下部发挥着正调控作用,而Phi亚家族主要成员在地上部起正向调控作用,这个结果与和表达的数据十分相似。为了深入分析在镉胁迫下的转录表达特性,试验选择差异表达显著 的这4 个基因对它们进行qRT-PCR 分析。结果显示与转录组测序结果相符,并且其都来源于Tau 亚家族或Phi 亚家族。我们可以推断,或许在面对镉胁迫时,甜菜GST 家族中的Tau 和Phi 亚家族在耐受性的提高上有着较大的潜力。
由此,在后续的研究我们可以多关注甜菜GST 家族中的Tau 亚家族与Phi 亚家族,这将为进一步探究甜菜在镉胁迫应答机理中的作用提供科学依据。
猜你喜欢
悦游 Condé Nast Traveler(2021年4期)2021-01-13
农民致富之友(2019年24期)2019-08-20
科学之谜(2019年3期)2019-03-28
科学之谜(2018年8期)2018-09-29
环球时报(2017-09-19)2017-09-19
农村百事通(2017年1期)2017-02-16
中学生理科应试(2016年4期)2016-11-19
恋爱婚姻家庭·养生版(2016年9期)2016-09-07
飞碟探索(2015年9期)2015-11-05
红领巾·探索(2015年9期)2015-09-10
