
基于乳酸的深度共熔溶剂提取秸秆木质素对纤维素酶水解效率的影响
2022-10-13 09:55蒲福龙伍尚炜郑映玲郑玉意侯雪丹
化工进展 2022年9期
蒲福龙,伍尚炜,郑映玲,郑玉意,侯雪丹
(广东工业大学生物医药学院,广东 广州 510006)
废弃木质纤维素生物质的高值化利用是实现国家“碳达峰”“碳中和”目标的途径之一,其中生物质纤维素的高效水解则是生物质成功利用的关键,但是由于木质纤维素生物质的天然顽固性,使其直接利用较为困难,因此木质纤维素在酶解之前通常会进行预处理,打破其致密的结构、分离生物质组分以提高纤维素酶的可及性。常用的预处理技术通常有机械粉碎、稀酸预处理、碱预处理和水热法等,不同的预处理方法有不同的优缺点。除了寻找更优化的预处理方法,关于木质纤维素生物质中关键组分对纤维素酶解影响的研究也必不可少。一般而言,生物质底物中的木质素是影响纤维素酶解的主要成分,主要通过两个方面阻碍酶解:一方面,木质素在物理结构上阻碍酶与纤维素有效接触而影响酶水解效率;另一方面,木质素能非特异性地吸附纤维素酶,并且降低酶的活性从而对纤维素酶水解产生负面影响。有研究发现,水溶性木质素对含木质素的底物酶解具有促进作用,主要是通过和木质素底物对酶的竞争性结合促进酶解,这表明,无论是水溶性还是非水溶性木质素都与酶进行了不同程度的吸附和结合,所以木质素的非特异性吸附对酶解是一个很重要的影响因素。众多研究表明,木质素对纤维素酶的吸附程度与木质素的结构特性密切相关。而木质素的结构与性质则主要取决于生物质的预处理方法。不同的生物质预处理获得的木质素内酚羟基、脂肪族羟基与羧基的含量存在差异。一般而言,木质素中的酚羟基会与酶形成氢键而影响纤维素酶活性,而羧基和脂肪族羟基则会使木质素的疏水性减弱,从而影响酶的催化效率。
深度共熔溶剂(deep eutectic solvents,DES)是一种由氢键供体和氢键受体组成的可设计溶剂,已被用于生物活性萃取、生物催化、金属加工等各种领域。DES 对木质素优良的解聚能力,使其在木质纤维素生物质预处理和提取木质素方面有很好的应用前景。DES 在预处理过程中提供了酸碱催化体系并主要通过破坏木质素中的—O—4醚键使木质素进行解聚,从而使生物质组分分离。各种类型的醇类、羧酸类DES对多种生物质原料展现出优良的预处理效率,特别对木质素组分的脱除效果显著。然而,现有研究对各类DES提取木质素组分的结构性质及其对纤维素酶水解效率影响规律的认识仍然不足,需要系统地研究以理解溶剂性质、木质素结构性质与多糖酶水解效率之间的关系。因此,本文以基于乳酸的DES提取水稻秸秆中的4种木质素为研究对象,通过研究木质素对纤维素酶的吸附特性,结合木质素的结构特性分析,阐明疏水性作用、氢键相互作用和静电作用在木质素对纤维素酶的吸附和抑制方面的影响规律,揭示基于乳酸的DES提取木质素对纤维素酶水解效率的影响机理。
1 材料和方法
1.1 材料与试剂
水稻秸秆收集于本地农田,晒干后经机械粉碎至250~400µm 的粒径,于60℃烘箱过夜至恒重,密封袋收集并干燥保存;来源于酵母的复合纤维素酶(Celluclast 1.5L,酶蛋白浓度为37.17mg/mL,滤纸酶活为93.69FPU/mL,含5.63g/L 葡萄糖和1.13g/L木糖),诺维信(中国)生物技术有限公司天津分公司;氯化胆碱、乳酸、盐酸胍、精氨酸、甜菜碱盐酸盐、微晶纤维素(Avicel)、柠檬酸、柠檬酸钠,Aladdin 公司;硫酸、乙醇等试剂均为国产分析纯。
1.2 实验方法
1.2.1 DES的制备
将乳酸和各种氢键受体按9∶1 摩尔比均匀混合,装入100mL磨口瓶中,置于一定温度(乳酸-氯化胆碱、乳酸-盐酸胍为80℃;乳酸-甜菜碱盐酸盐、乳酸-精氨酸为100℃)下搅拌均匀,待呈现均一、澄清、透明状态的溶液后停止加热,然后将获得的DES[乳酸-氯化胆碱9∶1(LC)、乳酸-盐酸胍9∶1(LGH)、乳酸-精氨酸9∶1(LArg)、乳酸-甜菜碱盐酸盐9∶1(LBH)]装入带螺旋口的西林瓶中干燥、密封保存以备用。
1.2.2 木质素的提取
将DES与水稻秸秆粉末以10%的固液比混合,于120℃下恒温搅拌6h(200r/min),随后加入适量无水乙醇,多次清洗离心分离混合物。收集离心后的醇洗上清液,旋蒸除去无水乙醇,加入3倍体积的水,置于4℃下沉淀48h。离心分离沉淀物,并用去离子水清洗木质素沉淀,冷冻干燥后即为DES提取木质素。4 种木质素样品分别记为Lignin-LC(L-LC)、 Lignin-LGH (L-LGH)、 Lignin-LArg(L-LArg)、Lignin-LBH(L-LBH)。
1.2.3 木质素对微晶纤维素酶水解效率的影响
在含4.8mL 的50mmol/L 醋酸-醋酸钠缓冲液(pH=4.8)的25mL具塞三角烧瓶中加入100mg微晶纤维素(Avicel),于50℃下预热10min后分别加入20mg的L-LC、L-LGH、L-LArg、L-LBH,最后添加200µL 复合纤维素酶(93.69FPU/mL)启动水解反应,恒温振荡(150r/min),定时取样300µL,于100℃水浴5min灭活终止酶反应。以高效液相色谱(HPLC)测定葡萄糖浓度,以未加木质素的酶水解反应为对照。HPLC(安捷伦1260,配有Aminex HPX-87H 色谱柱和示差检测器)测试条件:流动相为5mmol/L 硫酸水溶液,流速为0.5mL/min,柱温、检测器温度分别为65℃和50℃。所有实验重复3次,相对误差小于5%。
1.2.4 木质素对纤维素酶的吸附特性
用50mmol/L 的醋酸-醋酸钠缓冲液(pH=4.8)作为溶剂配制复合纤维素酶蛋白溶液(0.02~1.60mg/mL 浓度梯度),然后加入定量的木质素使其浓度为4g/L,放置于2mL离心管中并密封。将所有样品置于4℃下孵育3h(150r/min)后取出,离心(4℃、10000r/min)取上清液,以Bradford 法测定蛋白浓度。根据初始溶液和离心所得上清液中游离酶含量的差异,计算木质素对酶蛋白的吸附量。结果符合Langmuir 吸附等温线,计算公式如式(1)和式(2)所示。
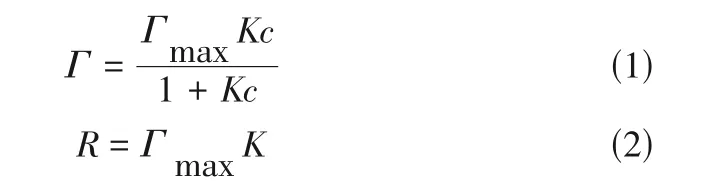
式中,为木质素吸附酶蛋白的量,mg/g木质素;为木质素最大吸附酶蛋白的量,mg/g 木质素;为Langmuir 常数,mL/mg 酶;为溶液中游离酶的浓度,mg/mL;为结合强度。
1.2.5 木质素疏水性
通过测定木质素对疏水性染料孟加拉玫瑰红的吸附,分析木质素表面疏水性。以50mmol/L 的醋酸-醋酸钠缓冲液(pH=4.8) 为溶剂配制含40mg/L 的孟加拉玫瑰红溶液,将其与不同质量的木质素混合,使木质素浓度梯度范围在2~10g/L,于50℃下孵育2h(150r/min)。低温离心(10000r/min、10min)后收集上清液,于543nm 处读取吸光值以测定游离染料含量。通过孟加拉玫瑰红的原始含量与游离含量的差异,得到木质素表面吸附染料的含量。分配系数(PQ)计算方法如式(3)。绘制木质素含量与PQ 值的回归曲线,获得木质素表面疏水性数据。

1.2.6 FTIR分析
采用研究型红外光谱仪对水稻秸秆和4种木质素样品进行红外分析,所用设备为Thermo-Fisher Nicolet 6700,波长范围为4000~400cm,通过32次扫描以0.48cm的分辨率记录光谱。
1.2.7 核磁分析
以定量磷谱(P-NMR)测定木质素组分羟基含量。精确称取40mg 木质素样品,溶解于500µL氘代溶剂中(氘代氯仿∶氘代吡啶=1∶1.6,体积比),然后,向溶液中加入50µL乙酰丙酮铬溶液(5.6mg/mL)和200µL内--羟基-5-降冰片烯-2,3-二甲酰亚胺(9.23mg/mL)。最后加入100µL磷化试剂(2-氯-4,4,5,5-四甲基-1,3,2-二氧磷烷溶液,TMDP)。混合15min 后以Bruker 600MHz(型号为AV600)核磁共振仪测定,采用TopSpin2.0软件处理数据,通过积分面积计算样品中酚羟基、醇羟基和羧基的含量。
以二维异核碳氢相关谱(2D HSQC NMR)分析木质素结构,参照本文作者课题组以前的测试条件并作部分修改。将120mg 木质素样品溶解于1.2mL 的DMSO-d中后转移至核磁管,采用Bruker 600MHz 核磁波谱仪进行测量,溶剂峰DMSO-d作为内标峰(C/H,39.5/2.49)进行校正。
2 结果与讨论
2.1 木质素对微晶纤维素酶水解效率的影响
由于特殊的结构性质,木质素对纤维素酶的抑制作用普遍存在。遗憾的是,现有研究对DES 溶解木质素影响纤维素酶活性的规律仍认识不足。因而,本文首先以不添加木质素的纤维素酶水解微晶纤维素反应为对照,考察4 种木质素(L-LC、LLGH、L-LArg、L-LBH)的添加对微晶纤维素酶水解效率的影响,结果如图1所示。木质素的添加降低了微晶纤维素的酶水解效率,在水解28h 后,木质素对酶水解的抑制作用大小顺序为:L-LC>L-LGH>L-LArg>L-LBH。具体而言,L-LC 组的木质素添加使葡萄糖的释放量相对于对照组降低了14.7%,而L-LGH 使葡萄糖的释放量降低较少(4.5%)。可见,不同的DES 提取的木质素对纤维素酶的抑制作用存在明显的差异性。
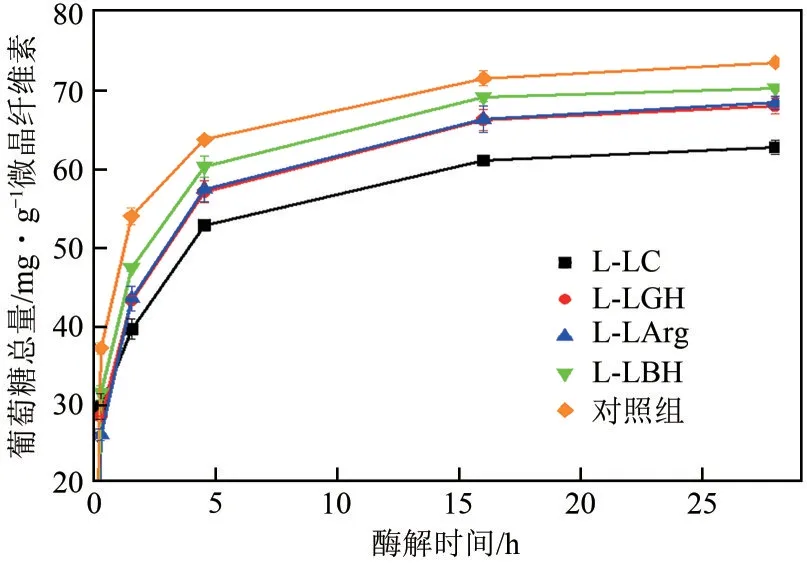
图1 微晶纤维素酶水解过程曲线
本文所用的4种基于乳酸的DES中,氯化胆碱作为氢键受体的研究最为普遍,已有研究表明其在溶解和提取木质素中表现出优良的性能,并有报道指出氯化胆碱在作用过程中与氢键受体之间的协同作用机制,即Cl通过离子相互作用而破坏木质素结构中的—O—4醚键而提取木质素。而甜菜碱盐酸盐与氯化胆碱比较接近,但其结构中含有羧基,疏水性比氯化胆碱弱,提取疏水性的木质素的能力则相对稍弱。当盐酸胍作为氢键受体时,其对生物质的作用能力也较强,原因是其强酸性和其与乳酸供体间的优良协同效应。可见,木质素的结构性质对其抑制纤维素酶的作用效力影响较大,木质素对纤维素酶的吸附作用主要源于疏水作用、静电作用和氢键作用的综合情况。因而,需要进一步探究这4种DES提取的木质素对纤维素酶吸附能力,以分析其对纤维素酶抑制强弱的原因。
2.2 木质素对纤维素酶的吸附性
众多研究表明,纤维素底物中木质素可通过对纤维素酶的非特异性吸附而影响酶催化效率,为了研究DES 所提取出的木质素对纤维素酶的吸附能力和吸附量,绘制了纤维素酶在L-LC、L-LGH、L-LArg和L-LBH上的Langmuir吸附等温线,如图2所示。根据吸附等温线,分别计算了纤维素酶的最大吸附量、Langmuir 亲和吸附常数和结合强度(表1)。结果显示纤维素酶在4种木质素上的吸附特性符合Langmuir 等温线模型,纤维素酶在木质素上的最大吸附量具有明显的差异,如LC提取的木质素对纤维素酶的最大吸附量最大,为46.35mg/g;吸附量最小的为LArg,仅4.52mg/g。而LGH 和LBH 提取的木质素对纤维素酶的最大吸附量比较接近,前者稍高。
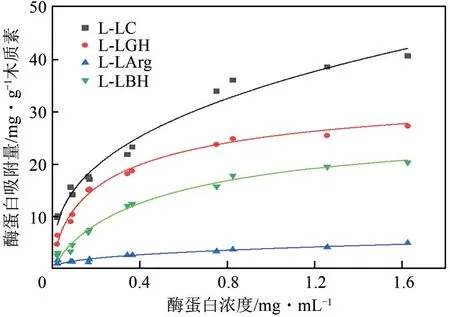
图2 木质素对纤维素酶蛋白的Langmuir吸附作用
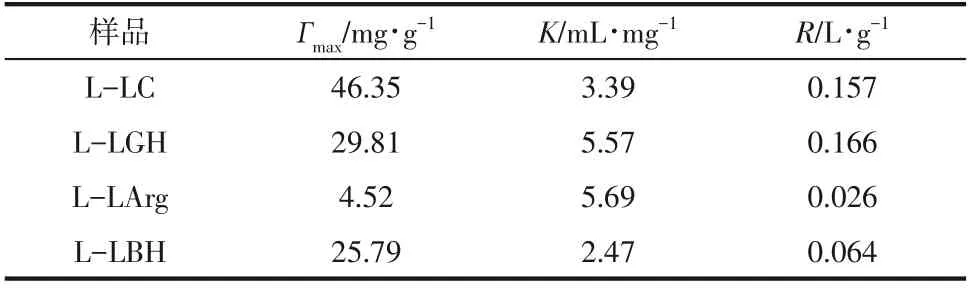
表1 Langmuir吸附等温线参数
结合强度代表了木质素对纤维素酶的总体吸附强度,其强弱规律与最大吸附量基本一致,除了L-LGH的结合强度比L-LC相对稍高。而吸附平衡常数代表结合亲和力的大小,除了L-LArg外,3种木质素显示的规律性与最大吸附量和吸附强度趋势基本一致,且这3种木质素对纤维素酶的吸附能力与微晶纤维素酶水解效率呈负线性相关性(图3),即吸附能力越强、吸附量越大,微晶纤维素的酶解效率越低,也说明木质素对酶的吸附能力越大,其对酶的抑制作用越明显。Wang 等也发现类似的现象:乙酸提取杨木木质素比球磨木质素对纤维素酶的吸附量大,前者比后者对酶水解的抑制作用则更强。在Yao等关于木质素对纤维素酶作用机制研究中,也证实了木质素对纤维素酶水解的抑制作用与其对纤维素酶吸附能力正相关,并提出非特异性吸附是影响纤维素酶水解效率的主要因素。
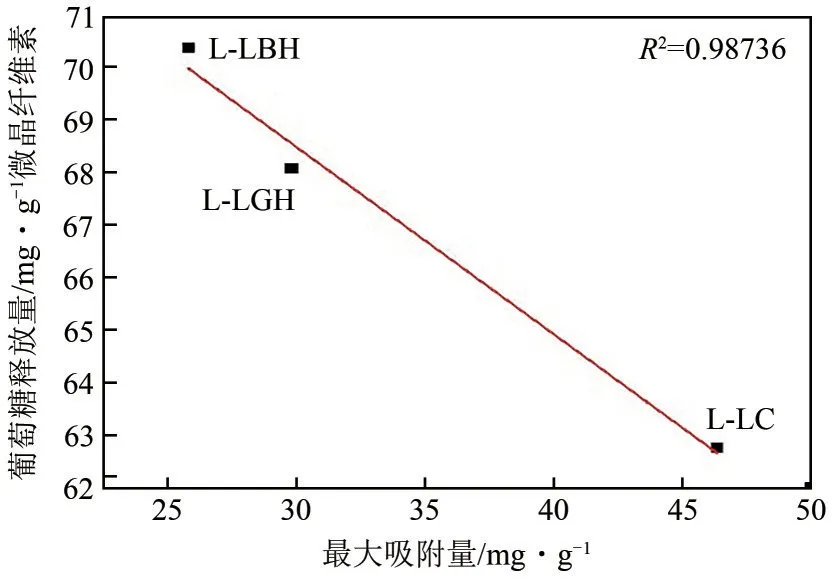
图3 最大酶吸附量与微晶纤维素酶水解效率的关系
因此,就本文而言,3种木质素对纤维素酶的抑制程度大小为L-LC>L-LGH>L-LBH,与其对纤维素的吸附能力大小规律基本相同。相比之下LLArg 组比较特殊,不符合这一规律。为进一步解释该现象,测定了各DES 提取木质素的组成成分,发现其他3 种木质素的纯度均为85%以上,而LLArg 提取的木质素含量仅55%(含11%的纤维素)。所以L-LArg对纤维素酶的吸附强度和最大吸附量较低的原因可能是是粗木质素样品中纤维素含量较高。同时,因为其中含有大量的纤维素组分,纤维素酶的底物结合域则与其天然底物纤维素组分有较强的亲和力,因而L-LArg 显示了最强的酶结合亲和力。由于天然的结合趋势对微晶纤维素的酶水解作用起促进作用,导致L-LArg 对酶解效率的抑制作用相对较弱。
2.3 木质素的疏水性对纤维素酶吸附作用的影响
从吸附性研究结果可知,木质素对纤维素酶的吸附很大程度上是抑制微晶纤维素酶水解的主要原因。如前文所述,木质素对纤维素酶的吸附作用包括疏水作用、静电作用和氢键作用。其中,疏水相互作用被认为是影响纤维素酶吸附非常重要的因素,如Sakkos 等研究结果证明,疏水性越强的底物对纤维素酶的吸附亲和性越强。本研究中讨论的4种DES提取的木质素疏水性大小顺序为:LLArg>L-LC>L-LHG>L-LBH(表2)。除L-LArg外,其余3种DES提取的木质素的疏水性大小与其最大纤维素酶吸附规律一致。由于纤维素酶具有疏水性,因此木质素疏水性越大,相应的吸附纤维素酶的量越多,对纤维素酶水解效率的抑制程度也越大。因而木质素疏水性与最大酶蛋白吸附量呈现明显的正线性相关性,且相关性较高(图4)。在利用对甲苯磺酸提取的甘蔗渣木质素对纤维素酶吸附性研究中,同样发现木质素疏水性与纤维素酶的吸附性的正相关性。Song等研究DES(LC)提取柳树、玉米秆木质素对纤维素酶的抑制作用时,也发现类似的现象。L-LArg 木质素的疏水性大的原因可能是大量的结晶纤维素组分残留,纤维素内部较为紧密牢固的氢键网络使其疏水性较大。这种疏水性区域是纤维素酶的天然底物,即使其与纤维素酶结合,也不会体现较强的抑制作用。因此,即使L-LArg 疏水作用最大,其对纤维素酶的最大吸附量最小,因而对纤维素酶的抑制作用也最小。
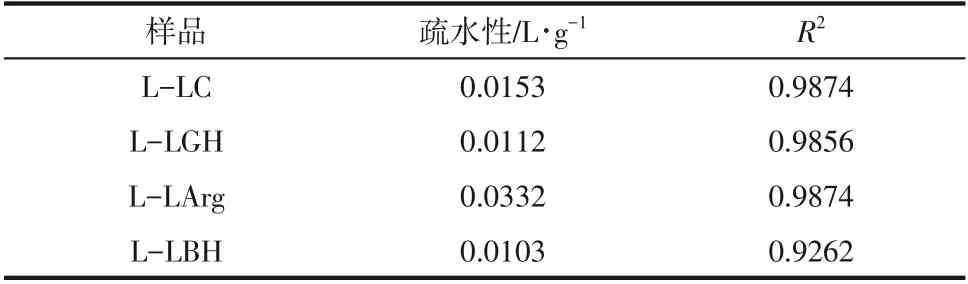
表2 木质素疏水性数据
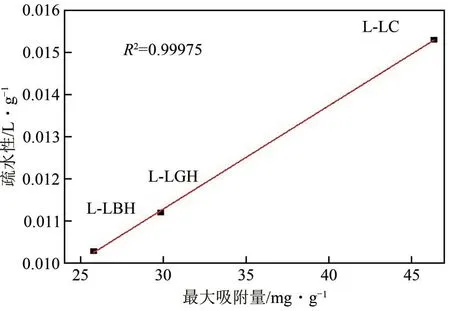
图4 木质素对酶蛋白的最大吸附量与木质素疏水性的关系
2.4 氢键作用对纤维素酶抑制效应的影响
为了进一步探究DES 提取木质素对纤维素酶的抑制作用机制,本文通过解析木质素的结构性质(红外光谱、P-NMR、HSQC)以深入探究木质素结构性质,尤其是氢键、静电相互作用对纤维素酶的抑制效应的影响。首先,红外光谱(图5)显示木质素的特征峰在1605cm、1423cm和1514cm处,为芳环骨架碳碳键的拉伸振动峰,所有木质素样品均在此处出现谱带,但L-LArg 的谱带明显弱于其他三个,说明4种DES提取木质素已具备基本骨架结构,但L-LArg 中木质素纯度较其他3 种相对较低。3360cm处峰为木质素分子中的羟基伸缩振动,说明木质素中含有丰富的酚羟基基团。而L-LC和L-LGH表现出更强的谱带,说明这两种木质素中含有更多的羟基基团。推测在较为强烈的DES预处理条件下(L-LC、L-LGH实验组),木质素中更多的酚亚结构被释放出来,因而更易与纤维素酶形成氢键相互作用。2930cm和2860cm处为脂肪链和O—CH基团中的CH—伸缩振动峰,LLArg 在2860cm处表现出明显较弱的谱带,说明L-LArg 在DES 提取过程中损失的甲氧基较多,而另外3 种木质素的甲氧基保留相对较多。在2930cm处,4 种木质素均显示较强谱带,证明脂肪链发生的断裂较少。所有木质素均在1740cm和1220cm处有明显的谱带,证明木质素中有羧基存在。据报道,羧基通过削弱疏水相互作用而阻碍酶在木质素上的吸附,同时由于许多纤维素酶组分因其等电点而表现出的总负电荷而加强静电斥力。然而,添加木质素仍然会导致纤维素酶的非生产性吸附,这证实静电斥力不足以对抗纤维素酶和木质素组分之间的疏水作用而形成的氢键。
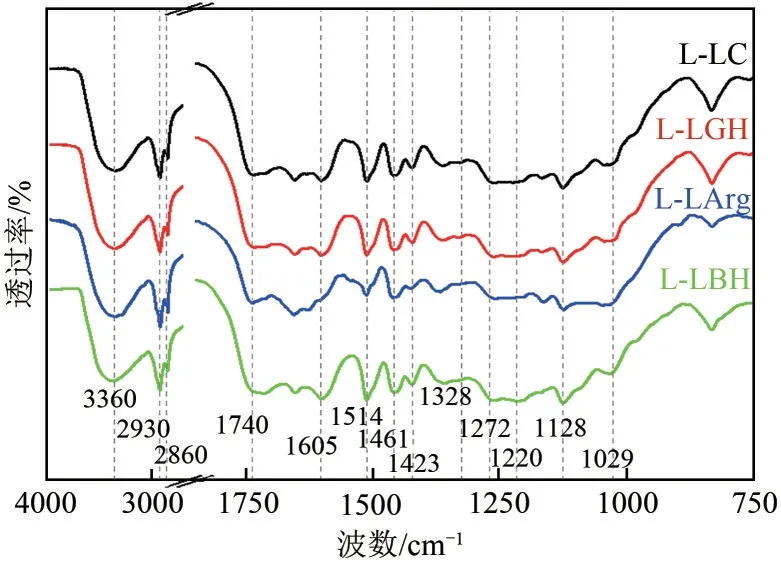
图5 木质素的红外光谱
2D HSQC NMR谱图(图6)显示了4种木质素的基本骨架结构,在侧链区[图6(a)]可以观察到较强的甲氧基信号峰。很明显,L-LArg 的所有峰信号均弱于其他3种木质素,这说明LArg的预处理强度最弱,对木质素的提取能力最弱,这与L-LArg最低的木质素得率(3.7%)所反映的规律一致。—O—4 和—对应的信号峰(A 和C)虽然都能检出,但是峰强度均较弱,这说明了4种DES在实验的木质素提取环境下对—O—4 和—的破坏程度都较高,4种木质素中L-LBH的—O—4和—保留最多。对于-5 的峰信号(B),仅在L-LC 和L-LBH的谱图中检出,而L-LGH和L-LArg中的-5均被破坏。另外,除了L-LArg外,其他3种木质素中阿拉伯糖单元(Ara)的信号均被检测到,说明LC、LGH、LBH在提取木质素的同时也提取了一定量的阿拉伯糖。L-LGH 和L-LBH 谱图中的少量木糖单元信号峰说明LGH和LBH对木糖有轻微的提取作用。芳香区谱图的规律与侧链区基本一致,S、G和H结构均能被很好地检测到,说明4种木质素中均含有S、G、H结构。而且峰强度大小排序为LLC>L-LGH>L-LBH>L-LArg,这基本印证了前面所提到的木质素对酶抑制程度大小排序的大致规律。
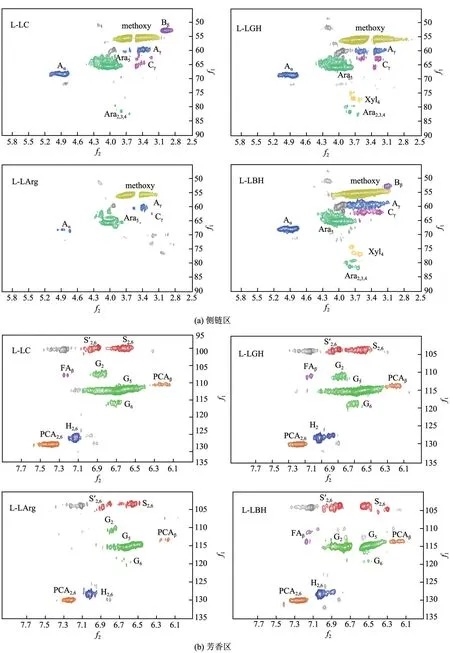
图6 木质素的HSQC图谱
P-NMR 结果表明(图7)4 种DES 木质素酚羟基含量和总羟基含量与其对纤维素酶的吸附能力之间有明显的线性相关性,两者含量越高,其对酶蛋白的吸附量越大。例如,L-LC 的酚羟基含量最大,为3.37mmol/g,所以也最大,为46.35mg/g;L-LArg的酚羟基含量最小,为0.966mmol/g,也最低(4.52mg/g);L-LGH 与L-LBH 的酚羟基含量接近,所以值也接近。此外,除了L-LArg 外,羟基含量与微晶纤维素酶水解效率线性负相关,即羟基含量越高、酶水解效率越低。L-LC 含有的脂肪族羟基、酚羟基和总羟基含量明显高于其他3种木质素,说明L-LC 更易与纤维素酶形成氢键结构,从而影响纤维素酶的水解活性。对于脂肪族羟基,含量的多少与纤维素酶的吸附量规律一致,该规律与纤维素酶水解的抑制程度大小相同(LLArg除外),即脂肪族羟基含量越高,越易促进木质素与纤维素酶相互作用,对酶的抑制作用则越强。酚羟基的含量,与脂肪族羟基规律存在差异(羟基含量L-LC>L-LBH>L-LGH>L-LArg)。由于LBH提取的木质素中纤维素含量(1.44%)低于LLGH 提取的木质素(4.03%),因而,其对木质素与碳水化合物连接键的破坏程度相对较多,酚羟基含量更高。总羟基含量是脂肪族羟基和酚羟基之和,4种木质素的总酚羟基含量基本上反应了DES的木质素提取作用程度,即L-LC>L-LBH>L-LGH>L-LArg,这体现了4种木质素与纤维素酶之间的氢键作用强弱规律。总体来说,木质素对纤维素酶的吸附作用大小,氢键作用仅是影响因素之一。
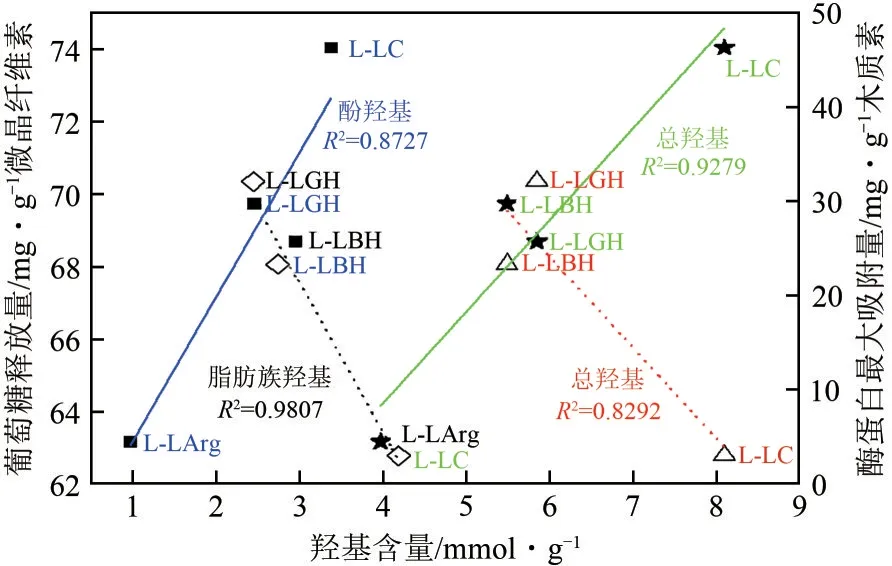
图7 羟基含量与酶解效率、酶蛋白最大吸附量的关联图
2.5 静电作用对纤维素酶抑制作用的影响
除了氢键相互作用,木质素与纤维素酶之间的静电相互作用也是影响两者之间吸附能力以及酶水解抑制程度的一项因素。有报道指出分离木质素中的羧基含量随着预处理程度的增加而增加,部分原因是木质素-碳水化合物复合物中酯键的断裂,从而释放了羧基官能团。本研究中,4 种木质素的羧基含量与酶蛋白吸附量、纤维素酶水解效率之间并无明显相关性(图8)。羧基含量最高的为L-LC,L-LBH 次之,L-LGH 最低,这3 种木质素羧基含量与总酚羟基含量大小一致,基本上符合预处理强度越大形成的羧基含量越多的规律。此外,羧基易在酶反应体系解离,从而使木质素带负电荷。文献表明,纤维素酶也带有负电荷,带负电的木质素易与纤维素酶形成静电斥力,从而降低木质素对纤维素酶的吸附量。然而本文中的几种木质素的羧基总体含量不高,羧基产生的静电斥力不足以消除更强的疏水相互作用和氢键相互作用力。所以,疏水相互作用和氢键相互作用主要决定了木质素对纤维素酶的吸附力及抑制作用大小,该结论与Song 等在研究对苯甲磺酸提取的木质素对纤维素酶吸附作用时的结果一致。
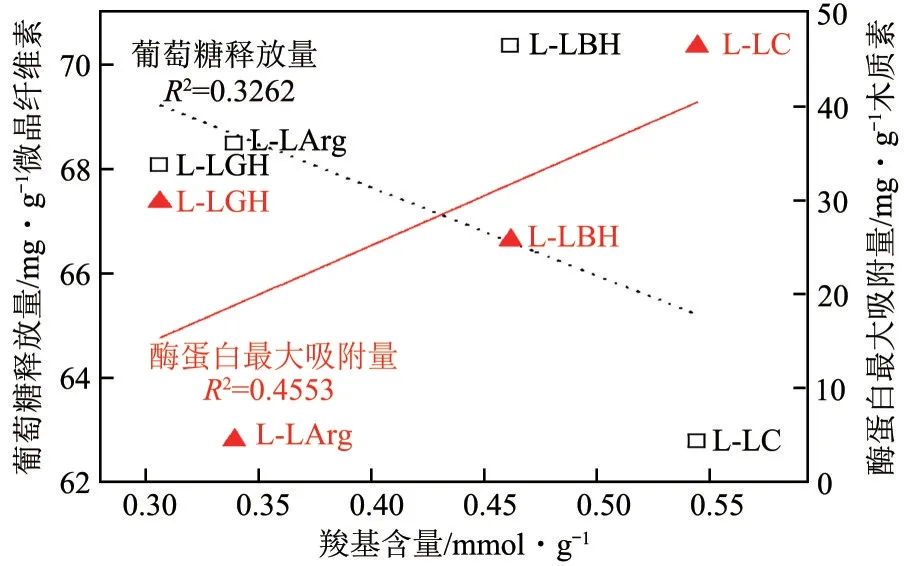
图8 羧基含量对酶解效率、木质素疏水性和木质素对酶蛋白的最大吸附量的影响
3 结论
(1)由木质素对纤维素酶的吸附性研究结果可知,4 种DES 提取木质素的吸附特性曲线符合Langmuir等温曲线模型,且木质素对酶蛋白的吸附性越强,其对酶的抑制效应也越大。
(2)木质素疏水性与酶蛋白的吸附性能也有高度的线性正相关性,即疏水性越大,其对酶蛋白吸附能力越强,纤维酶水解效率则越低。
(3)木质素的基本结构性质分析表明DES 提取强度越大,木质素产生的(脂肪、酚)羟基含量和羧基含量越高,与酶蛋白的氢键作用则越强,因而对纤维素酶水解效率的抑制程度越大。
(4)羧基的静电斥力效应虽可削弱吸附作用,但对总体的酶蛋白吸附和活性抑制程度影响较小。
(5)4种DES对木质素的提取能力和结构破坏作用能力最强的是LC,LGH 和LBH 基本相当,LArg 最弱,说明氯化胆碱作为氢键受体易于与乳酸氢键供体协同作用,以更好地破坏木质素和碳水化合之间的连接键、木质素的—O—4键,而关于这4种DES的氢键受体在木质素提取过程中作用机制的差异性有待进一步深入探究。
猜你喜欢
汽车维护与修理(2022年4期)2022-08-22
化工进展(2022年7期)2022-08-01
发明与创新·中学生(2019年10期)2019-11-17
中国造纸(2019年6期)2019-09-10
小天使·二年级语数英综合(2019年12期)2019-01-11
体育时空(2016年11期)2017-01-06
中学化学(2015年12期)2016-01-19
中学化学(2015年12期)2016-01-19
分析化学(2015年4期)2015-06-08
化学教学(2015年1期)2015-03-19
