
增龄性肌肉减少症小鼠肌肉组织抗氧化能力的特点
2022-10-18 11:43吴丹妮王秦玉刘珊珊季铎淳王力可于春艳
吉林大学学报(医学版) 2022年5期
郭 丹,温 旭,吴丹妮,王秦玉,刘珊珊,季铎淳,王力可,王 丹,董 营,于春艳
(1.北华大学基础医学院病理教研室,吉林 吉林 132013;2.吉林医药学院附属医院病理科,吉林 吉林 132013;3.北华大学附属医院超声诊断科,吉林 吉林 132000)
肌肉减少症(肌少症)由于机体骨骼肌质量和力量逐渐丧失,导致患者跌倒、骨折、肢体残疾和死亡等不良事件的发生,造成其生活质量下降,给家庭和社会带来较大的负担。既往研究[1]中,关于肌少症的定义已经演变为包括与年龄相关的骨骼肌功能丧失。近期研究[2-3]表明:线粒体可调节年龄依赖性的身体衰退和肌肉内稳态,线粒体稳态在维持线粒体功能和骨骼肌功能中发挥重要作用。线粒体在提供能量的同时会产生大量氧自由基,活性氧(reactive oxygen species,ROS)生成和抗氧化剂之间的动态平衡与线粒体功能稳态密切相关。BORI等[4]认为:线粒体功能紊乱对肌少症疾病发展至关重要,但骨骼肌线粒体抗氧化能力特点尚不明确,抗氧化与肌减少症之间关系的分子基础尚未完全阐明。核因子E2相关因子2(nuclear factor E2-related factor 2,Nrf2)是一种转录因子,通过与靶基因启动子中的抗氧化应答元件(antioxidant response element,ARE)结合,在介导细胞内抗氧化反应中发挥关键作用,Nrf2及其靶基因与调控线粒体功能有关[5-6]。研究[7]表明:通过恢复Nrf2活性能够防止因衰老导致的肌肉和心脏功能障碍。Nrf2缺乏能够促进衰老腓肠肌自噬的增加,是肌少症发生的潜在机制[8]。然而,Nrf2抗氧化途径在以氧化代谢为主的肌肉中的作用及其机制目前尚未完全阐明。
本研究利用C57/6J小鼠,观察其增龄性肌肉组织中ROS水平和Nrf2 mRNA及蛋白表达水平的变化,检测Nrf2依赖的抗氧化途径相关基因mRNA表达水平,探讨以氧化代谢为主的肌肉组织增龄后抗氧化应激的特点,阐明ROS生成和抗氧化间的动态平衡在增龄性肌少症中的作用,为预防或延缓年龄相关肌少症的发生发展提供新的思路。
1 材料与方法
1.1 实验动物、主要试剂和仪器SPF级雄性C57BL/6J小鼠24只,体质量18~20 g(北京华阜康股份有限公司),动物生产许可证号:SCXK(京)2014-0004,饲养于通风和安静的环境中,定时喂食,自由饮水。Nrf2和GAPDH单克隆抗体(美国Proteintech公司),过氧化物酶体增殖子活化受 体γ共 激 活 因 子1α(peroxisome proliferatorsactivated receptor γ coactivator-1 alpha,PGC-1α)、Nrf2及 抗 氧 化 应 激 基 因Nqol、Cat、Gclc、Sod1、Sod2和GAPDH引物由上海生工生物有限公司合成,TRIzol试剂(美国Invitrogen公司),PureLink RNA mini试剂盒(美国Ambion公司),预混型逆转录试剂(北京全式金生物技术股份有限公司),实时荧光定量PCR(real-time fluorescence quantitative PCR,RT-qPCR)检测试剂盒CFX96(美国Bio-Rad公司),ROS检测试剂盒(上海碧云天生物技术有限公司)。蛋白质电泳装置、转移系统和蛋白浓度分析仪(美国Bio-Rad公司),凝胶成像系统(德国Analytikjena分析仪器公司)。
1.2 HE染色观察各组小鼠比目鱼肌组织病理形态表现24只小鼠随机分为3月龄组(3 M组)、12月龄组(12 M组)和22月龄组(22 M组)。采用10%水合氯醛麻醉各组小鼠,剪开胸廓,暴露心脏,将灌注针头经左心室刺入主动脉,止血钳固定,剪开右心房,生理盐水快速灌注冲洗直至肝脏变白,采用4%多聚甲醛磷酸盐缓冲液(pH 7.4)灌注固定,取出各组小鼠完整比目鱼肌,置于4%多聚甲醛中固定24 h。常规脱水、透明、包埋和切片制作石蜡切片,HE染色后封片,Olympus显微镜下观察并拍照。
1.3 各组小鼠骨骼肌质量指数(skeletal muscle mass index,SMI)计算采用分析天平称量各组小鼠体质量和比目鱼肌质量,计算SMI。SMI=小鼠比目鱼肌质量(mg)/小鼠体质量(g)。
1.42′,7′-二氯荧光素二乙酸酯(2′,7′-dichlorodihydrofluorescein diacetate,DCFH-DA)荧光探针法检测各组小鼠肌肉组织中ROS水平称取20 mg新鲜比目鱼肌组织,按10mL·g-1RIPA组 织 裂 解 液 裂 解,取200 μ L匀 浆,4℃、12 000 g离心10 min,弃沉淀,按照ROS检测试剂盒说明书,于96孔细胞培养板中加入90 μL组织匀浆上清液和1 mmol·L-1DCFH-DA 10 μL,37℃温箱孵育30 min,于488 nm和535 nm波长处检测组织 匀 浆 中2′,7′-二 氮 荧 光 素 (2′,7′-dichlorofluorescein,DCF)荧光分布。以相对荧光强度代表ROS水平。
1.5 RT-qPCR法检测各组小鼠肌肉组织中PGC-1α、Nrf2和抗氧化基因mRNA表达水平取冻存比目鱼肌组织(30±10)mg,加入液氮充分研磨,采用TRIzol试剂提取总RNA,PureLink RNA mini试剂盒纯化,Nanodrop分光光度计NP-1 000测定260 nm和280 nm波长处吸光度(A)值,计算样本总RNA浓度并评估其纯度。采用预混型逆转录试剂,将总RNA(1 μ g)逆转录为cDNA,0.5 μg cDNA采用RT-qPCR试 剂盒进行PCR扩增,CFX96 RT-qPCR检测系统检测,PCR引物序列见表1。PCR反应条件:95℃、10 min预变性;95℃、15 s,60℃、1 min,40个循环;95℃、15 s,60℃、1 min,95℃、15 s。以GAPDH为内参,采用2-ΔΔCt法计算目的基因mRNA表达水平。ΔCt=目的基因Ct值-GAPDH Ct值。
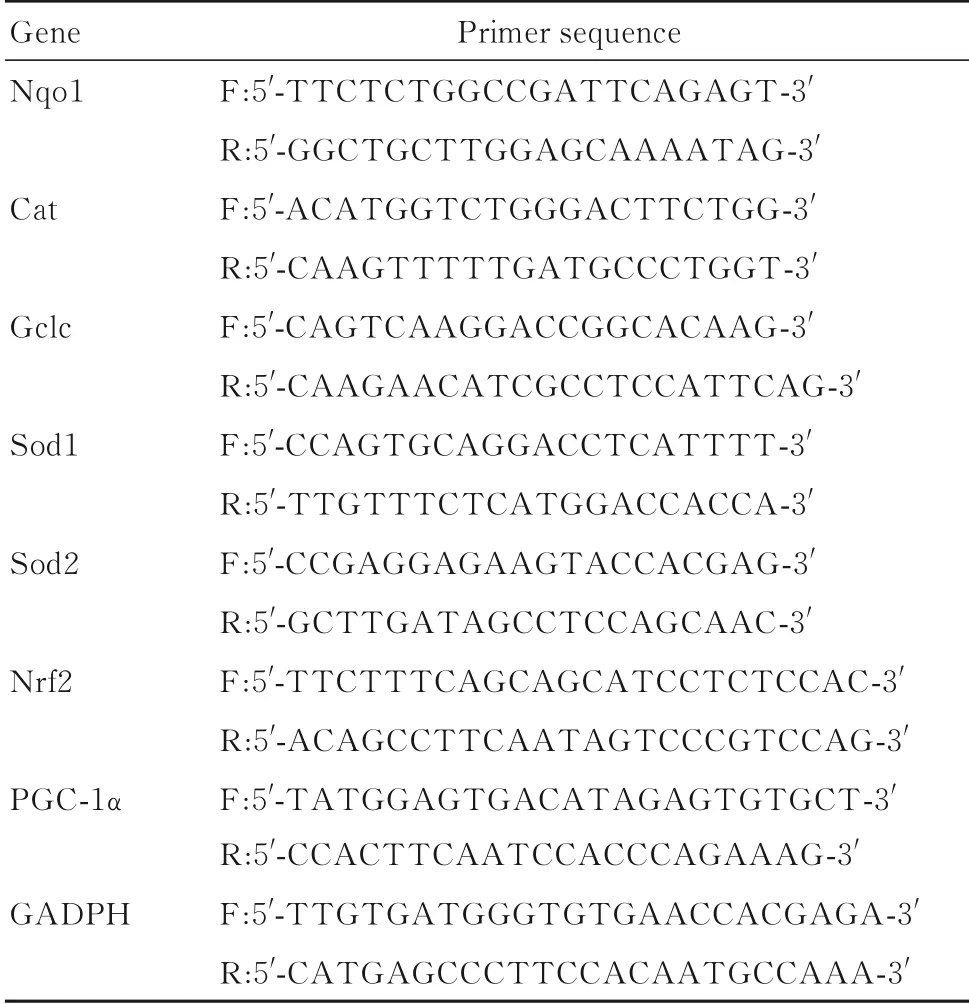
表1 PCR引物序列Tab.1 Primer sequences of PCR
1.6 Western blotting法检测各组小鼠肌肉组织中Nrf2蛋白表达水平称取(20±5)mg冻存比目鱼肌组织,按10 mL·g-1RIPA组织裂解液裂解,采用Bio-Rad法测定蛋白浓度,配制12 %聚丙烯酰胺凝胶,GAPDH等量蛋白上样对照,30 μg蛋白质样品SDS-PAGE电泳,将蛋白转至PVDF膜上,10%脱脂奶粉室温封闭1 h,TBST漂洗3次,每次10 min;加入抗体(Nrf2 1∶1 000,GAPDH 1∶5 000),4℃孵育过夜。次日TBST漂洗3次,每次10 min,加入辣根过氧化物酶标记的二抗(1∶2 000),室温摇床孵育2 h,TBST漂洗3次,每次10 min。ECL显色,凝胶图像分析系统分析蛋白条带灰度值,计算目的蛋白表达水平。目的蛋白表达水平=目的蛋白条带灰度值/GAPDH蛋白条带灰度值。
1.7 统计学分析采用SPSS 26.0统计软件进行统计学分析。各组小鼠SMI值,肌肉组织中ROS水 平,PGC-1α及 抗 氧 化 基 因Nqol、Cat、Gclc、Sod1和Sod2 mRNA表达水平,肌肉组织中Nrf2mRNA和蛋白表达水平均符合正态分布,以-±s表示,多组间样本均数比较采用方差分析,组间样本均数两两比较采用LSD-t检验。以P<0.05为差异有统计学意义。
2 结果
2.1 各组小鼠比目鱼肌病理形态表现3 M组小鼠比目鱼肌颜色以红色为主,形态规则,大小均匀,肌纤维周围可见细胞核;12 M组小鼠比目鱼肌颜色仍以红色为主,肌纤维形态规则,横截面积增大,部分肌纤维细胞核数量增多,少许肌纤维可见变性改变;22 M组小鼠比目鱼肌白色肌纤维增多,形态不规则,大小不均匀,肌纤维细胞核数量增多并出现核内移现象,肌细胞间隙明显增大,肌纤维变性增多。见图1。
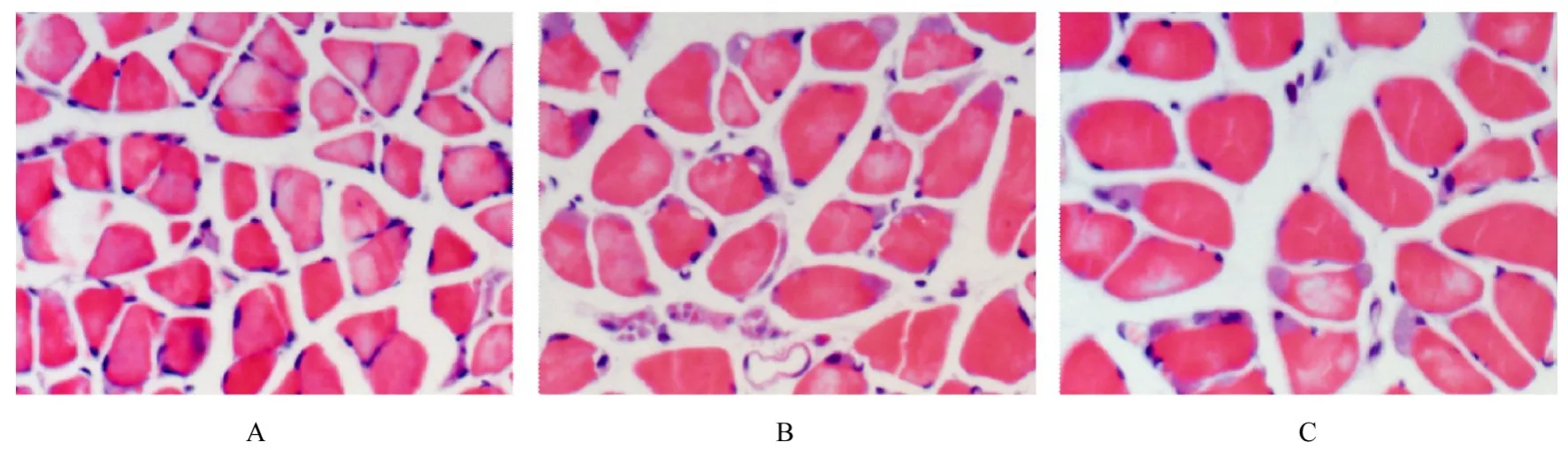
图1 各组小鼠比目鱼肌组织病理形态表现(HE,×400)Fig.1 Pathomorphology of soleus muscle tissue of mice in various groups(HE,×400)
2.2 各组小鼠骨骼肌SMI与3 M组[(0.61±0.08)mg·g-1]比较,12 M组和22 M组小鼠SMI[(0.60±0.07)和(0.54±0.09)mg·g-1]均降低,但差异无统计学意义(P>0.05);与12 M组比较,22 M组小鼠SMI值差异也无统计学意义(P>0.05)。
2.3 各组小鼠肌肉组织中ROS水平DCFH-DA荧光探针法检测结果显示:与3 M组[(1.12±0.08)AU]比较,12 M组和22 M组小鼠肌肉组织中ROS水平[(1.45±0.42)和(2.00±0.19)AU]均升高(P<0.05);与12 M组比较,22 M组小鼠肌肉组织中ROS水平明显升高(P<0.01)。
2.4 各组小鼠肌肉组织中PGC-1α和Nrf2 mRNA表达水平RT-qPCR结果显示:与3 M组比较,12 M组和22 M组小鼠肌肉组织中PGC-1α mRNA表达水平均明显升高(P<0.05或P<0.01);与12 M组比较,22 M组小鼠肌肉组织中PGC-1α mRNA表达水平降低,但差异无统计学意义(P>0.05)。与3 M组比较,12 M组小鼠肌肉组织中Nrf2 mRNA表达水平差异无统计学意义(P>0.05),22 M组小鼠肌肉组织中Nrf2 mRNA表达水平明显升高(P<0.01);与12 M组比较,22 M组小鼠肌肉组织中Nrf2 mRNA表达水平明显升高(P<0.01)。见表2。
2.5 各组小鼠肌肉组织中抗氧化基因Nqol、Cat、Gclc、Sod1和Sod2 mRNA表达水平与3 M组比较,12 M组小鼠肌肉组织中抗氧化基因Nqol、Cat、Gclc、Sod1和Sod2 mRNA表 达 水 平 均 明 显升高(P<0.01);与12 M组比较,22 M组小鼠肌肉组织中抗氧化基因Nqol、Cat、Sod1和Sod2mRNA表达水平明显升高(P<0.05或P<0.01),Gclc mRNA表达水平降低(P<0.05)。见图2。
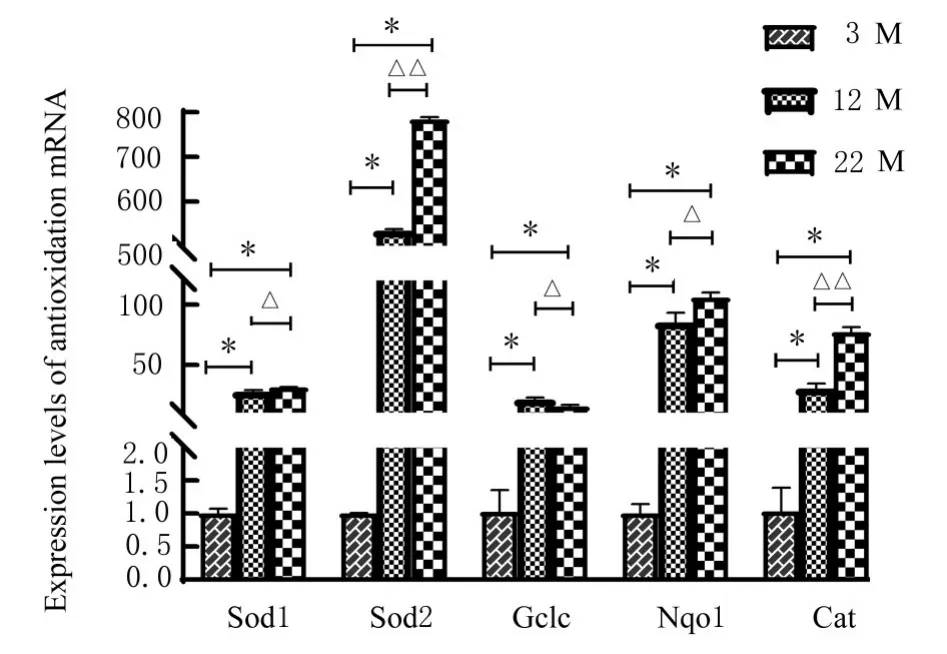
图2 各组小鼠肌肉组织中抗氧化基因mRNA表达水平Fig.2 Expression levels of antioxidant gene mRNA in muscle tissue of mice in various groups
2.6 各组小鼠肌肉组织中Nrf2蛋白表达水平与3 M组比较,12 M组小鼠肌肉组织中Nrf2蛋白表达水平明显降低(P<0.01),22 M组小鼠肌肉组织中Nrf2蛋白表达水平明显升高(P<0.01);与12 M组比较,22 M组小鼠肌肉组织中Nrf2蛋白表达水平明显升高(P<0.01)。见图3。
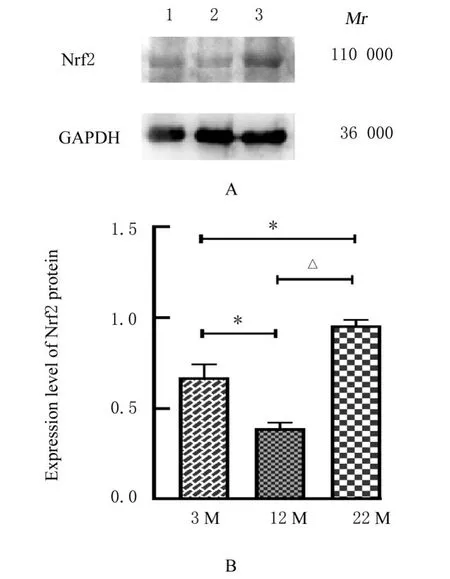
图3 各组小鼠肌肉组织中Nrf2蛋白表达电泳图(A)和直条图(B)Fig.3 Electrophoregram(A)and histogram(B)of expression of Nrf2 protein in muscle tissue of mice in various groups
3 讨 论
年龄增长相关的骨骼肌老化是导致原发性肌少症发生的主要原因,衰老过程导致骨骼肌质量变化,因而骨骼肌质量和功能的下降是衰老最显著的结果。比目鱼肌是以慢肌纤维氧化型为主的肌纤维,其肌红蛋白和线粒体数量较多。本研究HE染色结果显示:22 M组小鼠比目鱼肌白色肌纤维增多,形态不规则,肌纤维细胞核数量增多并出现核内移现象,肌细胞间隙增大,细胞数量减少。比目鱼肌形态学表现和骨骼肌SMI值降低,提示骨骼肌的数量和肌细胞形态发生改变,增龄性比目鱼肌表型发生肌少症的改变。
衰老等因素特异性损害骨骼肌线粒体功能[9]。研究[10-11]报道:衰老骨骼肌线粒体密度降低,基质空泡化、嵴变短,大部分线粒体出现去极化或无功能。骨骼肌内的线粒体分为肌膜下线粒体和肌纤维间线粒体,前者为肌细胞膜活性转运和基因转录提供ATP,后者为肌肉组织中收缩肌丝提供ATP,与氧化磷酸化过程有关[12-13]。PGC-1α作为线粒体生物发生和氧化呼吸的主要调节因子之一,能够增加氧化磷酸化,影响ROS产生[14-15]。线粒体电子传递链是细胞内ROS的主要来源之一,本研究结果显示:增龄性比目鱼肌中ROS水平和PGC-1α mRNA表达水平均升高,与以往研究[16]结果一致。氧化应激时PGC-1α诱导ROS解毒酶升高,抑制ROS水 平。研 究[17]表明:PGC-1α水平升高可维持骨骼肌横截面积,阻止比目鱼肌早期分解代谢途径,缓解比目鱼肌萎缩。
研 究[18]显 示:PGC-1α通 过 激 活Nrf2而 调 控抗氧化基因表达,下调PGC-1α表达,则完全抑制了Nrf2结合谷氨酰半胱氨酸连接酶催化亚单位(glutamate-cysteine ligase catalytic subunit,GCLC)基因的启动子元件,降低超氧化物歧化酶2(superoxide dismutases 2,SOD2)和谷氨酰半胱氨酸连接酶(glutamate cysteine ligase,GCL)蛋白表达水平。此外,Nrf2能够协同调节抗氧化剂谷胱甘肽(glutathione,GSH)产生和再生,通过调控Nqol等调节ROS水平,使Nrf2为细胞提供重要的细胞保护反应[19]。在小鼠肌营养不良模型中,使用Nrf2激活剂肌肉功能得以改善,提示Nrf2缺乏增强了老年骨骼肌中线粒体ROS产生[20]。本研究结果显示:随着增龄性改变,小鼠肌肉组织中ROS水平升高,Nrf2 mRNA和蛋白表达水平均升高,小鼠肌肉组织中抗氧化基因Nqol、Cat、Sod1和Sod2 mRNA表达水平升高,提示增龄性肌肉组织中部分抗氧化能力与Nrf2依赖抗氧化途径有关。
研究[21-22]证实:线粒体不仅是ROS的主要来源,也是氧化损伤的靶点,随着年龄的增长,ROS产生增加。Nrf2是一种组成性活性转录因子,能够感知氧化还原平衡并控制抗氧化防御系统的基因,由于比目鱼肌以氧化代谢为主,高水平的氧化磷酸化水平自然暴露于过氧化氢和超氧化物等,由此推测比目鱼肌对氧化还原失衡可作出迅速反应[23-24]。线粒体具有调节氧化还原稳态功能,使ROS不会过度产生或累积,进而激活分解代谢途径[25-26]。研究[27]显示:线粒体中ROS水平升高可导致适应性反应,增强对氧化应激的抵抗力,并可能导致氧化应激的长期降低。本研究结果显示:增龄性比目鱼肌组织中Nrf2 mRNA和蛋白及抗氧化基因表达水平升高,ROS水平升高,提示增龄性改变中,比目鱼肌具有一定的氧化应激抵抗能力,然而ROS并未完全清除。根据衰老的自由基理论,ROS会产生反馈环,其形成增加可能会对细胞造成累积性损伤,从而加剧线粒体功能障碍[28]。提示细胞核中心化和细胞数量减少等骨骼肌老化的表现可能与ROS水平升高有关。HARMAN[29]表明:静止或力竭运动均会产生过量ROS,导致氧化应激相关的组织损伤和肌肉收缩力受损。氧化损伤是一种将衰老与慢性疾病联系的机制,包括骨骼肌质量和功能的逐渐丧失。因此,减轻氧化损伤是预防或延迟慢性病发作或延长健康期的潜在途径[2]。Nrf2依赖的抗氧化信号途径可能是治疗以氧化代谢为主的肌肉减少症的一个潜在治疗靶点。
猜你喜欢
体育科技文献通报(2022年3期)2022-05-23
中国畜牧杂志(2021年10期)2021-12-03
医学综述(2021年16期)2021-12-01
国际放射医学核医学杂志(2021年10期)2021-02-28
肉类研究(2020年9期)2020-12-14
灌篮(2020年36期)2020-05-16
科技资讯(2020年3期)2020-04-07
运动(2018年14期)2018-07-16
体育科学(2018年3期)2018-04-20
肉类研究(2017年8期)2017-11-16
