
高寒煤矿渣山植被建植年限对土壤细菌多样性的影响
2022-11-04 09:12道日娜苏贝贝李希来马林雄铁晓龙
草地学报 2022年10期
道日娜, 张 英, 苏贝贝, 李希来, 马林雄, 铁晓龙
(青海大学农牧学院,三江源区高寒草地生态省部共建重点实验室, 青海 西宁 810016)
青海省江仓煤矿区是青海省最大的矿区,海拔3 800 m左右,气候寒冷、干旱、氧缺乏、年积温低,生长季节短。机械堆积起来的一座座渣山,严重破坏了草地资源,加重了水土流失,并且其本身土壤结构性差、缺乏植物所需要的养分,导致矿区生态环境无法在短时间内进行恢复。为此,对高寒江仓煤矿区进行了人工建植。由于高寒矿区特殊的地理位置,多采用不覆土直接种植植被的修复方式,因此具有较大难度。
欧美等国家很早开始了对矿区生态系统的研究[1]。一些学者研究表明,人工建植比自然恢复更有利于矿区植被的快速恢复[2]。我国关于土壤细菌多样性的研究主要见于农田和丛林生态系统[3],大多在黄土丘陵区与低海拔区[4],一般采用排土以后与客土混合来进行土壤改良,而对高寒地区的研究屈指可数。吴道铭等[5]认为,芒属植物对重金属有较强的抗逆性,可以根据矿山生态状况、修复目的以及芒属植物的生物特性,制定矿山植被的恢复模式。孙梦媛等[6]研究了煤矿排土场不同复垦模式对土壤肥力的影响,得出豆科植物和杨树林混合复垦能有效提高土壤碱解氮、有机质含量,酶活性以及微生物量。以上研究能够表明植被恢复显著改善矿区土壤质量,但相关研究主要集中在高寒矿区生态修复的土壤理化性质测定和植被重建方面[7]或土壤酶活性以及植物生物量方面,缺少对不同植被建植年限土壤特征演变的研究报道。研究表明,不同建植年限下,地上植被、土壤养分含量、土壤酶活性以及微生物多样性以及物理结构均有显著变化,年限越长,越有利于土壤物理结构的改良以及有机质的积累[8-9]。
土壤微生物,例如土壤细菌、真菌、放线菌及微藻类等主导并参与了有机物的分解、植物的生长等重要过程[10-11],受微环境的影响比较大[12-13],对土壤质量变化具有重要指示作用[14]。土壤细菌参与土壤中多数养分的转化[15],其在土壤微生物中占绝对优势[16-17]。因此,土壤细菌多样性可以用来衡量建植后土壤质量状况,监测矿区环境变化[18-20]。土壤酶是指一些在土壤中具有生物催化能力的特殊蛋白质化合物的总称,主要来源于土壤中植物、动物及微生物活动的产物[21]。土壤酶在土壤生态系统物质能量的转化循环过程中起着重要作用,参与土壤中各种生物化学过程[22-23]。土壤酶的活性大致反映了某一种生态环境条件下土壤生物化学过程的相对强度,因此,可以反映土壤肥力状况和土壤质量的生物活性[24-25]。细菌和酶共同参与土壤养分循环以及转化、土壤的发育以及更新[26],进而促进植物的生长,是最能反映土壤质量的敏感指标[27]。在微生物群落研究方面,大多采用磷脂脂肪酸法,该方法不能全面反映土壤微生物的群落结构以及多样性的变化,因此具有一定的局限性。近年来高通量测序的发展,使快速、准确的获取土壤微生物的信息成为可能。因此,本研究主要以江仓矿区渣山不同建植年限的草地为研究对象,利用高通量测序技术分析土壤细菌多样性对复绿草地的响应特征,测定土壤养分含量以及酶活性,在此基础上探讨矿区渣山不同建植年限的土壤养分含量、酶活性与土壤细菌多样性的关系,以期为未来的高寒矿区建植与生态重建工作提供理论基础。
1 材料与方法
1.1 研究区概况
研究地点位于青海省海北州刚察县江仓煤矿区,该区地处高寒地带,海拔3 800~4 200 m,最低气温—35.6℃,最高气温19.8℃,年平均气温—2.8℃,年降水量477.1 mm,多集中在夏季,年蒸发量1 049.9 mm,为中低山,气候严寒。江仓矿区内分布大量高寒草甸、沼泽,属高寒沼泽湿地。坡面的基质均由煤矸石与堆积的渣土组成,阳坡,沿渣山底部边缘砌筑围堰,以增加坡面的稳定性。样地建植方法一致,均有人工种植垂穗披碱草(Elymusnutans)、冷地早熟禾(Poacrymophila)、星星草(Puccinelliatenuiflora),以2∶1∶1的比例分别种植于2015年 5月、2017年5月以及2019年5月下旬,播量8.094 3 kg·hm-2,播后覆盖无纺布。在此期间,无施肥、浇水、放牧等情况。矿区无干扰的原始植被的海拔3 788 m,地理坐标为99°38′E,38°03′N,优势种主要为小嵩草(Kobresiapygmaea)、矮嵩草(Kobresiahumilis)、细叶苔草(Carexrigescens)。
1.2 研究方法
2019年9月20日赴野外样地,在试验区(如图1)选取2019年(编号Y19)、2017年(编号Y17)、2015年(编号Y15)建植的草地,用消过毒的无菌铲采集0~20 cm的根围土壤样品,采集渣山外围原生植被土壤(编号YS)为对照,重复3次,装入自封袋中混合均匀,低温下带回室内进行化学性质、酶活性与细菌群落特征的测定。

图1 试验区样地布局图Fig.1 The geographical location of the layout of the sample plot
1.3 测定方法
(1)土壤化学性质测定:土壤全氮(Total nitrogen,TN)含量采用半微量凯氏定氮法,全磷(Total phosphorus,TP)含量采用氢氧化钠熔融法,全钾(Total potassium,TK)含量采用氢氧化钠熔融法,碱解氮(Available nitrogen,AN)含量采用碱解扩散法,速效磷(Available phosphoru,AP)含量采用碳酸氢钠浸提法,速效钾(Available potassium,AK)含量采用醋酸铵浸提法,土壤酸碱度采用电级法(水土比2.5∶1)进行测定,土壤有机质(Organic matter,OM)含量采用重铬酸钾容量法测定[28-29]。
(2)土壤酶活性测定:土壤蔗糖酶活性采用3,5-二硝基水杨酸比色法,淀粉酶活性采用二硝基水杨酸比色法,脲酶活性采用靛酚蓝比色法,磷酸酶活性采用磷酸苯二钠比色法,过氧化氢酶活性采用高锰酸钾滴定法[30]。
(3)土壤细菌多样性的测定:先进行基因组DNA的提取,称取土壤样品,在液氮中研磨呈细粉末,振荡混匀;将上清转移到新的离心管中离心2 min;在吸附柱中加入溶液50 μL(10 mL D2600-50T)和离心后的上清液,并加入漂洗液后离心;拿出吸附柱干燥后,加洗脱液,放在新的离心管中离心;然后将离心管中的液体重新加入吸附柱中离心,所得物为土壤微生物。将DNA溶液进行电泳;再以稀释后的基因组DNA为模板,来进行PCR基因扩增。采用Illumina MiSeq测序平台的通用引物对土壤细菌16SrDNAV3-V4区域进行PCR扩增。在进行实验的时候,需做对照实验(无菌去离子水),代替模板DNA。扩增完成后对扩增产物进行纯化、定量和均一化[31]。利用Illumina MiSeq进行上机测序及分析。
1.4 数据测定
采用Excel 2007 进行数据整理,运用SPSS对土壤养分及酶活性采用单因素方差分析(One-Way ANOVA)。利用Duncan复极差法进行差异显著性检验(P<0.05)。土壤酶数据运用Origin 64做柱状图。细菌多样性使用Silva(细菌)数据库比对,用Blast软件物种比对注释。细菌群落结构与土壤因子的关系,采用百迈客云平台的RDA分析法进行土壤因子与土壤真菌的相关性分析。
2 结果与分析
2.1 不同建植年限草地土壤化学性质及酶活性变化
不同建植年限的草地土壤化学性质的结果如表1所示。研究区域土壤为弱碱性土壤。Y19与Y17的pH值差异显著(P<0.05),而Y15与YS之间没有显著性差异。各样地之间OM,AK,AP,TK,TN含量有差异显著性(P<0.05)。Y19和Y17样地之间TP和AN含量无显著差异;Y15和YS样地之间TP,AN含量无显著差异;但Y19和Y17与Y15和YS之间呈现出了显著性差异(P<0.05)。随着建植年限的增长,TP含量、TK含量、AN含量呈上升趋势;而TN含量、AP含量、AK含量和OM含量随着建植年限的增长而上升,但恢复到一定年限时,呈下降趋势。

表1 不同建植年限草地土壤化学性质Table 1 Soil chemical properties of grassland in different construction years
对不同复绿年限草地土壤酶活性的变化进行分析,如图2所示,随着建植年限的增长,土壤酶活性呈上升趋势。脲酶活性在不同建植年限之间有显著差异(P<0.05),与Y19相比,Y15脲酶活性增加了64.6%。过氧化氢酶的活性在Y19时最低,Y15时最高,与YS相比,建植土壤的过氧化氢酶的活性较大。由图2(c)可知磷酸酶的活性在各建植年限的土壤中差异显著(P<0.05);在Y15时活性达到最高,但与YS相比,降低了43.7%。淀粉酶和蔗糖酶活性在各样地的值随着建植年限的增长呈现出上升的趋势,但建植一段时间过后才能恢复到原生植被的土壤状态。总体来说,随着建植年限的增长,土壤养分含量逐渐增加,使得土壤微生物分泌的土壤酶增加。

图2 不同复绿年限草地土壤酶活性Fig.2 Soil enzyme activities in different years of construction grassland
2.2 不同建植年限草地土壤细菌多样性分析
对不同建植年限草地土壤细菌群落丰富度与多样性进行分析,如表2所示,Ace,Shannon,Chao指数在各样地都出现了显著性的差异(P<0.05),Simpson指数无显著差异。所有样本覆盖度均在99%以上,表明测序结果可靠。Y17的细菌丰富度(Ace指数与Chao指数)最高,YS的最低。从Shannon指数来看,Y17的最高,为8.59;Y15的最低,为6.47。表明Y17的细菌多样性最高,Y15则相反。

表2 不同建植年限草地土壤细菌群落丰富度与多样性指数Table 2 The richness and diversity index of soil bacterial community in grassland with different greening years
不同建植年限草地土壤细菌群落物种组成分布,如图3所示。随着建植年限的增长,细菌物种组成也发生了较大变化。Y19当中,鞘氨醇单胞菌属(Sphingomonas)、未培养到的Micrococcac、未培养到的Intrasporan为优势菌属。而在Y17和Y15中,优势菌属为鞘氨醇单胞菌属(Sphingomonas)、未培养到的Subgroup-6,Ellin6067。YS中的优势菌属为鞘氨醇单胞菌属(Sphingomonas)和苍白杆菌(Ochrobactrum)。Ellin6067在Y17与Y15中的相对丰度在3.5%~6.6%之间,而在Y19与YS中只占到了0.1%左右。苍白杆菌(Ochrobactrum)在Y19 中占到了6.5%左右,而在其他样本中占到了0.1%以下,甚至更少。Y19中未培养到的Micrococcac占到了19.5%左右,在其他当中只占到了0.7%~1.3%。鞘氨醇单胞菌属(Sphingomonas)在4个样本中都是以优势菌属的形式存在,也证明了该属的菌株有着较强的环境适应性。
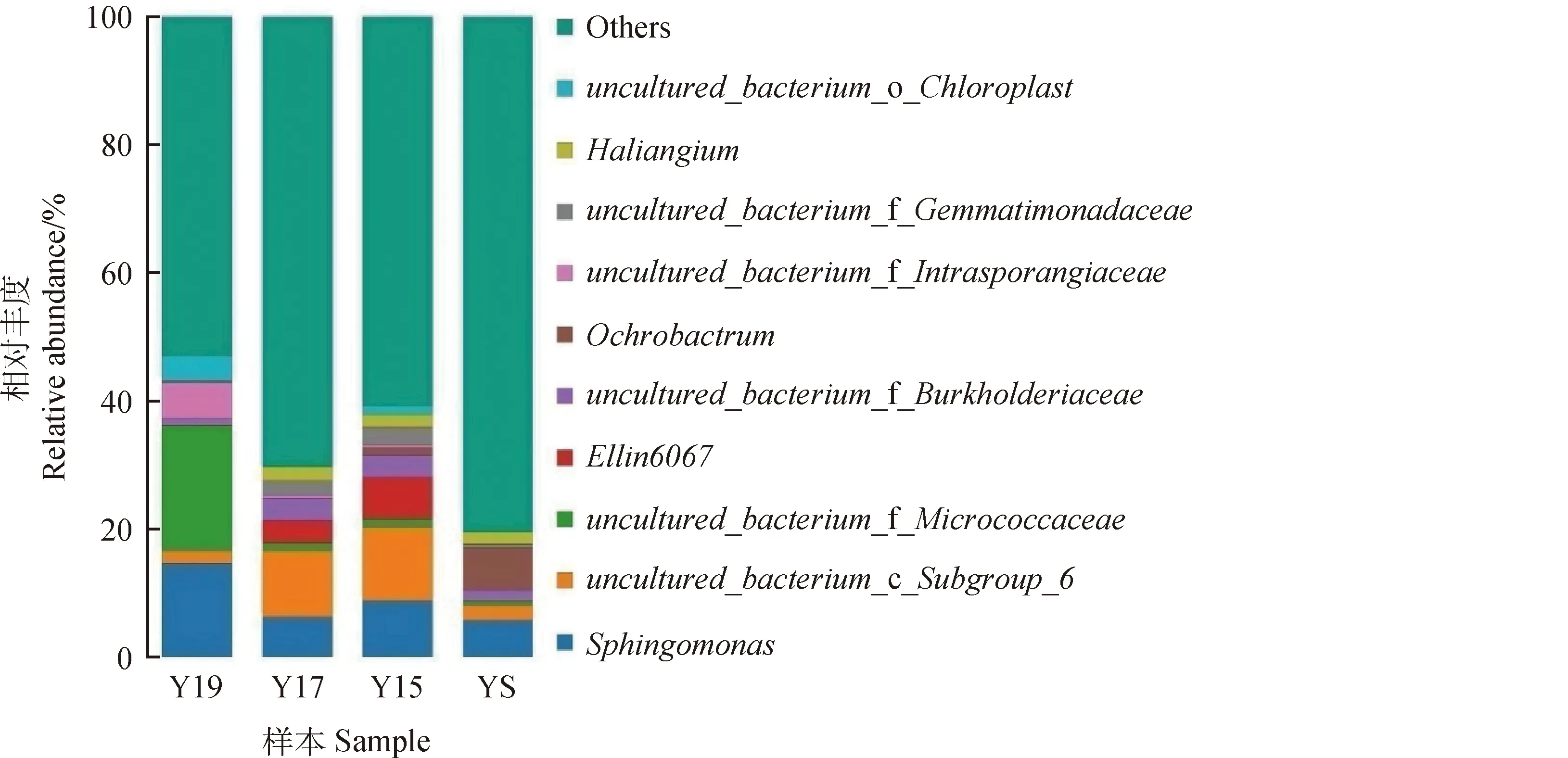
图3 不同建植年限草地土壤细菌物种组成分布Fig.3 Composition and distribution of soil bacterial species in grassland with different years of construction注:一种颜色代表一个物种,色块长度表示物种所占相对丰度比例;为使视图效果最佳,柱状图只显示丰度水平前10的物种,并将其他物种合并为Others在图中显示,Uncultured代表未得到培养的物种Note:Acolor represents a species,and the block length represents the relative abundance of species. To maximize the view effect,only the top 10 species with abundance level are shown in the histogram,and the other species are combined into Others as shown in the figure. Uncultured represents the species that have not been cultured
由图4可知,物种在三个样本中的分布不太均匀。放线菌门(Actinobacteria)离Y19近且相对丰度较大,而离其他两个顶点较远。变形菌门(Proteobacteria)离顶点Y15较近,分布较多;而离Y19相对较远,分布较少。拟杆菌门(Bacteroidetes)在Y19中分布的较多,Y15中的分布较少。酸杆菌门(Acidobacteria)在Y17的分布较多,而绿弯菌门(Chloroflexi)在Y19和Y17中的分布较多,在Y15中不存在。
在属分类水平上,根据不同建植年限草地土壤细菌群落在属水平的物种注释以及丰度信息,选取了土壤细菌丰度排名前20个物种及其在每个样本中的丰度信息绘制了热图,可以使高丰度和低丰度的物种分块聚焦。结果显示(图5),Y19的主要细菌属为uncultured-bacterium-f-Micrococcac,Sphingomonas,Massilia,uncultured-bacterium-f-Intrasporangi,uncultured-bacterium-o-Chloroplast、uncultured-bacterium-c-Subgroup-6,相对丰度在3.0%~19.6%之间。YS的主要菌属为Ochrobactrum,uncultured-bacterium-f-Lachnospira、乳球菌属(Lactobacillus)、Brevibacterium,相对丰度在2.5%~6.6%之间。Y17的主要菌属为uncultured-bacterium-f-Saprospiraceae,uncultured-bacterium-f-A4b,相对丰度在3.1%~4.7%之间。Y15的主要菌属为Ellin6067,uncultured-bacterium-o-Saccharimonadales,丰度在2.0%~6.6%之间。Y17与Y15的优势菌群的区域颜色相近,表明它们有较高的群落结构相似性。而Y19与其他样本、YS与其他样本的颜色变化明显不同,说明他们的结构差异明显。

图4 不同建植年限草地土壤细菌三元相图Fig.4 Ternary phase diagram of soil bacteria in grassland with different years of construction注:等边三角形的顶点表示各样本,点的大小代表属的相对丰度Note:The vertices of the equilateral triangle represent each sample,and the size of the points represents the relative abundance of the genus

图5 不同建植年限草地土壤细菌物种丰度热图Fig.5 Heat map of soil bacterial species abundance in grassland with different greening years注:热图中着色方格对应的值为样品在属分类上的Z值,即为样品在该分类上的相对丰度和所有样品在该分类的平均相对丰度的差除以所有样品在该分类上的标准差所得到的值;方格颜色越红,说明该样品相对丰度越高Note:The value corresponding to the colored squares in the heat map is the Z value of the sample in the genus classification,that is,the value obtained by dividing the difference between the relative abundance of the sample in this classification and the average relative abundance of all samples in this classification by the standard deviation of all samples in this classification;The redder the grid color,the higher the relative abundance of the sample
不同建植年限草地土壤细菌群落的主成分分析结果表明(图6),主成分1(PC1)、主成分2(PC2)以及主成分3(PC3)的样品差异性贡献率为7.12%,27.46%和65.42%,是主要的差异来源。Y19位于PC1的正值区域和PC2的负值区域及PC3的正负两侧。Y17位于PC1的负值区域和PC3的正值区域以及PC2的正负两侧。Y15位于PC1,PC3的负值区域。YS位于PC1和PC2以及PC3的正值区域。这4个样本之间,间隔位置较大,细菌群落结构发生了较大变化。

图6 不同建植年限草地土壤细菌主成分分析Fig.6 Principal component analysis of soil bacteria in grassland with different years of construction
2.3 不同建植年限草地土壤化学性质、酶活性与细菌多样性
不同建植年限草地土壤化学性质、土壤酶活性与土壤细菌的相关性分析结果如图7所示,第一排序轴和第二排序轴解释了细菌群落变化的30.55%和19.23%。土壤有机质(OM)、全氮(TN)、土壤酶的射线都较长,因此对土壤细菌群落的影响较大。碱解氮(AN)、有机质(OM)、全氮(TN)与全磷(TP)含量与蔗糖酶(SC)、脲酶(UE)、过氧化氢酶(CAT)、淀粉酶(AMY)和磷酸酶(PA)等土壤酶活性,以及酸杆菌门(Acidobacteria)等细菌门类相对丰度之间呈正相关性,彼此之间受影响较大。而全钾(TK)、速效磷(AP)、速效钾(AK)与芽单胞菌门(Gemmatimonadetes)、酸杆菌门(Acidobacteria)、绿弯菌门(Chloroflexi)、装甲菌门(Armatimonadetes)、拟杆菌门(Bacteroidetes)等细菌门类相关性较强。其中,变形菌门(Proteobacteria)、硬壁菌门(Firmicutes)对环境的适应性强,耐低温。总体来说,土壤化学性质、土壤酶活性以及土壤细菌之间相互影响、相互制约。

图7 不同建植年限草地土壤化学性质、土壤酶活性与 土壤细菌相关性分析Fig.7 Correlation analysis of soil chemical properties,soil enzyme activities and soil bacteria in grassland with different greening years注:TN表示全氮;TP表示全钾;TK表示全钾;AN表示碱解氮;AP表示速效磷;AK表示速效钾;OM表示有机质;pH表示酸碱度;SC表示蔗糖酶;UE表示脲酶;CAT表示过氧化氢酶;PA表示磷酸酶;AMY表示淀粉酶;Acidobacteria表示酸杆菌门;Actinobacteria表示放线菌门;Firmicutes表示硬壁菌门;Gemmatimonadetes表示芽单胞菌门;Proteobacteria表示变形菌门;Rokubacteria表示己科河菌门Note:TN indicate total nitrogen;TP indicate total potassium;TK indicate total potassium;AN indicate alkali-hydrolyzed nitrogen;AP indicate available phosphorus;AK indicate available potassium;OM indicate organic matter;pH indicate pH value;SC indicate sucrase;UE indicate urease;CAT indicate catalase;PA indicate phosphatase;AMY indicate amylase
3 讨论
3.1 不同建植年限草地土壤化学性质与酶活性
对高寒煤矿渣山进行建植可提高土壤养分含量、酶活性以及细菌群落多样性。土壤养分和酶活性能直接或间接反映土壤肥力状况。张小琴等[32]发现随着植被恢复时间的增加,氮、磷、钾等逐渐积累,pH值降低[33]。杨鑫光等[34]、王锐等[35]等研究认为,高寒煤矿在建植短期内土壤速效氮含量呈现出先上升后下降的趋势。在本研究中,速效氮含量在建植1年和3年之间无显著差异,而在建植5年时显著增加,并未出现下降趋势。造成这种差异的原因可能与土壤生物化学过程有关,还需进一步进行分析研究。本试验中,建植1年时,土壤pH值为8.48,建植5年时pH值为7.91,虽然pH值有所降低,但依旧呈弱碱性。这是由于建植时间较短,植被生长过程中损耗了大量养分,并未得到及时的补充。建植5年的全氮含量与建植3年相比含量降低,是由于植物的生长发育主要受到氮素的影响,建植初期植物可用氮素缺乏,随着建植时间的延长,土壤氮素随之减少。全钾含量随着建植年限的增长呈上升趋势;而速效钾含量呈先上升后下降的趋势。由于渣山气温低,海拔高,有机质分解缓慢,导致速效磷与速效钾含量建植5年时呈下降趋势,因此渣山在建植后需及时补充磷、钾等速效肥料,从而满足植被生长所需。土壤有机质是土壤微生物能量的来源和养分的储备库[36-37],在植物生长过程中极其重要[38],能够反映植被盖度以及生物量的状况[39]。在本试验中,随着建植年限的增长,土壤有机质含量呈现出先增加后下降的趋势。而诸多的研究发现,随着建植时间的增长,有机质含量呈显著增长趋势[40-43]。从理论上讲,煤矿渣山气候寒冷,土壤有机质分解缓慢,引起土壤有机质呈上升状态。造成这种差异的原因,还需进一步进行试验与探讨。
植被恢复后,土壤脲酶与土壤蔗糖酶的活性表现出上升的趋势。随着建植年限的增长,煤矿渣山草地土壤酶活性均表现出上升趋势。土壤酶的主要来源是根系分泌物与植物残体分解物。在植被建植前期,土壤养分含量较低,水土流失较为严重;而在后期,植物凋落物积累,根系分泌物增加,养分含量逐渐升高,土壤酶活性也就相应的提高。建植年限短,蔗糖酶活性较低的原因可能是由于地上生物量少,减少了蔗糖酶的产生。在李鹏飞等[44]、胡华英等[45]等的研究中,随着植被的恢复土壤磷酸酶呈现出无规律的变化。这与本试验结果不一致。土壤磷酸酶受较多因素的影响,是由土壤微生物、植被丰富度的不同所导致。
3.2 不同建植年限草地土壤细菌多样性
对煤矿渣山不同建植年限草地土壤细菌多样性分析发现,细菌丰富度与多样性呈现出先增加后下降的趋势,建植3年时达到最高值。土壤细菌丰富度的增加主要是由于根系分泌物的增加和养分利用率的提高[46]。本研究中,在属水平物种分布图中可看到,鞘氨醇单胞菌属(Sphingomonas)在各个样地均为优势菌属。从三元相图中可以看到,放线菌门(Actinobacteria)、变形菌门(Proteobacteria)、拟杆菌门(Bacteroidetes)、酸杆菌门(Acidobacteria)与绿弯菌门(Chloroflexi)为优势菌门。变形菌门(Proteobacteria)的丰度随着建植年限的增加而上升,这是由于养分含量与酶活性增加,促进了土壤细菌的丰度增加。放线菌门(Actinobacteria)与绿弯菌门(Chloroflexi)的丰度随建植年限的增长而呈下降趋势。这是因为放线菌门(Actinobacteria)与绿弯菌门(Chloroflexi)在较为贫瘠的土壤环境中具有生存优势[47-48]。
3.3 影响土壤细菌多样性的关键因子
土壤酶由植物根系、微生物和动物等分泌,参与土壤养分合成或者分解等过程,与土壤微生物直接或间接地影响土壤生化反应,因此土壤酶活性、土壤理化性质与土壤细菌存在着密切的联系。刘善江等[49]学者指出,土壤酶是各种生化反应的催化剂,同时也是土壤细菌活性强弱的表现和影响因子之一,印证了本文结果。土壤全氮、速效氮含量与土壤酶活性对土壤细菌的影响较大,说明土壤全氮、速效氮含量与土壤酶活性是土壤中较为重要的环境影响因子。Lauber等[50]研究发现土壤pH值与土壤细菌群落关系密切相关,这与本试验结果有差异。这种差异可能是由于土壤酶活性的影响因素还有其他菌类或细菌种类,并且作用方式不同所导致。蔗糖酶、脲酶、过氧化氢酶、淀粉酶和磷酸酶等土壤酶活性,以及酸杆菌门(Acidobacteria)、己科河菌门(Rokubacteria)、变形菌门(Proteobacteria)、硬壁菌门(Firmicutes)等细菌门类相对丰度之间呈正相关关系,彼此之间受影响较大。其中,蔗糖酶和脲酶是土壤碳、氮循环的限制因子之一。马转转等学者[51]的研究表明,土壤细菌群落与土壤脲酶、蔗糖酶活性呈显著相关关系,这与本试验结果相一致。朱灵等[52]研究发现,土壤酶活性与绿弯菌门(Chloroflexi)、变形菌门(Proteobacteria)、装甲菌门(Armatimonadetes)与芽单胞菌门(Gemmatimonadetes)相对丰度存在显著正负相关性,但与其他门类的相关性不显著,这与本试验结果有些差异。这可能是由于植被的不同直接或间接影响酶活性与土壤细菌,并且土壤中的细菌类群较多,土壤酶活性与土壤细菌之间有着复杂而非单一的关系。有些土壤养分、土壤酶活性与土壤细菌群落关系不显著,这可能是由于土壤酶活性的影响因素还有其他菌类或细菌种类繁多,并且作用方式不同所导致。
4 结论
对煤矿渣山不同建植年限(1年、3年、5年)草地土壤酶活性与土壤细菌多样性研究发现:随着建植年限的增长,TP,TK,AN含量及土壤中酶活性呈上升趋势,TN,AP,AK和OM含量刚开始时随着建植年限的增长而上升,但土地恢复到一定年限时,呈下降趋势;4个样地中鞘氨醇单胞菌属(Sphingomonas)为共有的优势菌属;土壤OM和TN含量及土壤酶活性对土壤细菌群落的影响较大;土壤细菌相对丰度与土壤养分、酶活性存在密切的联系,随建植年限的增长,植被覆盖度、根系分泌物增加,土壤养分含量及酶活性增加,最终促进细菌多样性提高,从而生态环境逐渐改善。
猜你喜欢
科学技术与工程(2022年26期)2022-11-01
当代水产(2022年8期)2022-09-20
中国农学通报(2022年14期)2022-06-01
油气田环境保护(2022年2期)2022-05-09
昆明医科大学学报(2022年2期)2022-03-29
食品安全导刊(2021年20期)2021-08-30
现代畜牧科技(2021年4期)2021-07-21
世界竹藤通讯(2020年5期)2020-11-11
天津农林科技(2020年3期)2020-08-13
当代工人(2019年22期)2019-12-20
