
两种草坪草单种与混种对铅-锌-镉复合污染的生理响应
2022-11-04 08:11徐彦红张淑炜刘宇宇
草地学报 2022年10期
张 莉, 徐彦红, 张淑炜, 刘宇宇, 席 溢
(贵州大学动物科学学院, 贵州 贵阳 550025)
矿产资源的开发是造成土壤重金属污染的重要原因。如黔西北水城-赫章矿带是贵州铅(Pb)、锌(Zn)矿的主要产地,有研究显示,贵州省赫章县新官寨农业土壤中Pb、Zn、镉(Cd)、汞(Hg)、铜(Cu)、砷(As)平均含量分别为337 mg·kg-1,648 mg·kg-1,9.0 mg·kg-1,0.44 mg·kg-1,121 mg·kg-1和17 mg·kg-1,分别是贵州省农业土壤背景值的7.5,7.9,26.4,2.2,4.7和0.8倍[1]。微生物不能分化重金属,但重金属可以被微生物富集,土壤一旦被重金属污染,其自然净化的过程和人工管理将变得十分艰难[2]。研究发现重金属污染环境中能正常生长的植物,并将其用于土壤修复具有重要意义[3]。
植物修复是一种利用绿色植物来去除环境污染物的绿色修复技术,具有独特优势。其一,植物可以将污染物加工成可直接去除的物质形态或将其转化为毒性小甚至无毒的物质,从而对污染环境进行彻底治理;其二,一些植物种植后可进行多次修剪,从而最大程度实现减少土壤重金属污染的目的。超富集植物常被选择用于植物修复过程中,目前国内外报道的超富集植物已有721种,其中超过70%为镍超富集植物[4]。然而,专门针对Pb,Zn,Cd等重金属的超富集植物研究较少。
多年生黑麦草(LoliumperenneL.)属于禾本科黑麦草属,须根发达,抗性强,可富集多种重金属。研究表明[5],黑麦草对Pb,Zn,Cd的毒害具有较强的耐性,对尾矿砂的改良作用明显,可作为Pb,Zn矿区污染土壤生态修复的植物材料。徐卫红等[6]发现在锌污染的土壤中,当Zn2+浓度在0~520 mg·g-1时促进了多年生黑麦草的生长。余游等[7]研究发现,野地瓜和黑麦草套种的条件下,黑麦草地上部对Cd的富集量为5.64×103mg,野地瓜和多年生黑麦草套作提高了多年生黑麦草对Cd的富集能力。高羊茅(FestucaelataKeng ex E. Alexeev)是禾本科早熟禾亚科羊茅属多年生草本植物,是一种较好的绿色草坪用草。有研究表明,高羊茅具有一定的吸收和耐受土壤中重金属的能力[8]。徐佩贤等对四种冷季型草坪植物对Cd的耐受性与积累特性进行比较,结果表明高羊茅对Cd的耐受性最好,其次为多年生黑麦草高羊茅[9]。刘骐华等在铜、镉、铅对高羊茅种子萌发及幼苗生长影响的研究中,发现高羊茅在镉胁迫下表现修护最强的耐受性[10]。马志强等[11]研究发现一定浓度的茉莉酸甲酯可以增强高羊茅对镉污染土的修复能力。目前,重金属Pb,Zn,Cd对多年生黑麦草和高羊茅的影响主要集中于单一金属的胁迫,而关于重金属复合胁迫的研究较少。由于Pb,Zn,Cd在污染土壤中不是单独存在的,而是相互伴生存在的。因此,对于Pb,Zn,Cd重金属复合胁迫的研究具有重要的现实意义。本研究通过盆栽实验,研究多年生黑麦草和高羊茅单种与混种对Pb-Zn-Cd复合污染的生理响应,以期为Pb-Zn-Cd污染土中草坪草种植的适宜性研究提供一定的理论参考。
1 材料与方法
1.1 试验方法
1.1.1试验设计 土壤经过风干、研磨和过5 mm筛后,将土壤装入规格为48 cm×33 cm×26 cm(长×宽×高)的塑料花盆(每盆装土21 kg)待用。以贵州省赫章县新官寨农业土壤中重金属浓度作为试验参照值[1],Pb(NO3)2,Zn(NO3)2·6H2O,Cd(NO3)2·4H2O溶于水后加入土壤中,Pb,Zn,Cd离子浓度分别为337 mg·kg-1,648 mg·kg-1,9 mg·kg-1,混匀,静置3个月作为供试土壤。基肥为复合肥,施肥量为52.5 g·m-2。选取籽粒饱满的高羊茅、多年生黑麦草种子用10% H2O2消毒10 min后用水冲洗几遍后撒种,待出苗一致后进行间苗,每盆种6行,每行留苗30株。混种,一行为多年生黑麦草,一行为高羊茅。盆与盆之间间隔10 cm,随机放置。植物生长过程中及时去除杂草,防治虫害,并不定期浇水,使田间持水量保持在60%左右。种植2个月后修剪一次,待其再生长一个月后,此时两种植物都处于营养生长阶段,测定不同种植方式下多年生黑麦草和高羊茅的地上部分生理指标。
1.2 供试材料
供试土壤取自贵州大学西校区试验基地,取样深度为20 cm以内表层土。其Pb,Zn,Cd含量分别为5.76 mg·kg-1,16.50 mg·kg-1,0.03 mg·kg-1。土壤基本理化性质为:pH值5.2,有机质3.8 g·kg-1,全氮0.59 g·kg-1,全磷0.46 g·kg-1,全钾18.23 g·kg-1,碱解氮26.5 g·kg-1,有效磷24.7 g·kg-1,速效钾182.5 g·kg-1。供试种子高羊茅(‘Gold Crown’)和多年生黑麦草(‘雅晴’),购于贵州众智恒生态科技有限公司。重金属添加剂以分析纯Pb(NO3)2,Zn(NO3)2·6H2O,Cd(NO3)2·4H2O的形式添加。
1.2.1测定内容与方法 叶绿素a、叶绿素b以及类胡萝卜素含量采用采用95%乙醇浸提法。生理指标检测均参考李合生[12]等方法,超氧化物歧化酶(Superoxide dismutase,SOD)活性采用NBT光化还原法;过氧化物酶(Peroxidase,POD)活性采用愈创木酚法;过氧化氢酶(Catalase,CAT)活性采用碘量法;丙二醛(Malondaldehyde,MDA)含量硫代巴比妥酸(TBA)显色法;游离脯氨酸(Proline,Pro)含量采用茚三酮比色法间接测定;可溶性糖含量采用蒽酮比色法;可溶性蛋白含量采用考马斯亮蓝法。巯基化合物的测定:还原型谷胱甘肽(Reduced glutathione,GSH)含量、半胱氨酸(Cysteine,Cys)含量、非蛋白巯基(Non-protein sulfhydryl,NPT)含量采用试剂盒测定,试剂盒由苏州科铭生物技术有限公司提供,具体操作步骤按照相关规定说明书进行。
耐性的综合评价:应用模糊数学隶属函数法综合评价Pb-Zn-Cd复合污染对不同种植方式下多年生黑麦草和高羊茅生理指标的影响以及同种种植方式下多年生黑麦草和高羊茅耐受性的比较,先利用公式X=(X-Xmin)/(Xmax-Xmin)或X=1-(X-Xmin)/(Xmax-Xmin)分别对多年生黑麦草和高羊茅的叶绿素a、叶绿素b、类胡萝卜素、叶绿素、光合色素、POD,SOD,CAT,MDA,Pro,可溶性糖,可溶性蛋白,Cys,NPT,GSH进行隶属函数值计算[13]。式中,X为参试植物某一指标的测定值,Xmax和Xmin分别为所有材料中该指标的最大值和最小值,然后把每一指标在不同种植方式下的隶属值累加求平均值,最后把每份材料各项指标隶属函数值累加求平均值,根据各材料平均隶属函数值大小确定多年生黑麦草和高羊茅对Pb-Zn-Cd复合污染的耐受性,平均值越大,耐性越强;反之,耐性越弱。
1.2.2数据处理方法 SigmaPlot 14.0用于整理数据和图形的制作,统计分析软件SPSS 20.0用于数据单因素方差分析(One-way ANOVA)和邓肯多重比较(Duncan’s multiple comparison)。
2 结果与分析
2.1 不同处理对两种草坪草光合色素含量的影响
在无污染处理下,两种草坪草的混种(HL,HF)和单种相比降低了多年生黑麦草和高羊茅的叶绿素a、叶绿素b、类胡萝卜素、叶绿素以及光合色素的含量。Pb-Zn-Cd复合污染下高羊茅单种和无污染处理相比,叶绿素a、叶绿素b、类胡萝卜素、叶绿素以及光合色素的含量被提高,多年生黑麦草的叶绿素a、叶绿素b、类胡萝卜素、叶绿素以及光合色素的含量被降低,但未达到显著差异;但是混种模式下(PHL,PHF),多年生黑麦草叶绿素a、叶绿素b、类胡萝卜素、叶绿素以及光合色素的含量被提高,高羊茅叶绿素a、叶绿素b、类胡萝卜素、叶绿素以及光合色素的含量被降低,均未达到显著差异(表2)。
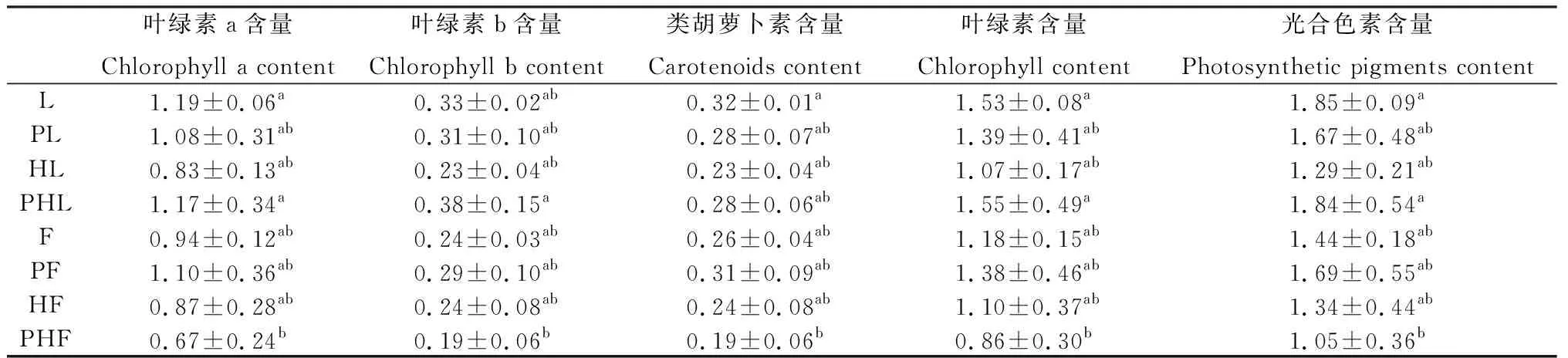
表2 不同处理对两种草坪草光合色素含量的影响Table 2 Effects of different treatments on photosynthetic pigment content of two turfgrasses 单位:mg·g-1
2.2 不同处理对两种草坪草抗氧化酶系统的影响
由图1A所示,添加Pb-Zn-Cd均提高了多年生黑麦草和高羊茅的POD活性;Pb-Zn-Cd污染下,相对于单种(PL,PF)而言,混种(PHL)对多年生黑麦草和高羊茅POD活性影响不显著;混种(PHF)显著提高了高羊茅POD的活性(P<0.05)。未添加Pb-Zn-Cd时,相对于单种(L,F)而言,混种模式下(HL,HF)高羊茅和多年生黑麦草POD活性变化也不明显。由图1B可知,添加Pb-Zn-Cd时,均降低了多年生黑麦草和高羊茅的SOD活性,相对于单种(PL,PF)而言,混种(PHL,PHF)提高了多年生黑麦草和高羊茅SOD活性;当不添加Pb-Zn-Cd时,对于单种(L,F)而言,混种(HL,HF)降低了多年生黑麦草SOD酶活性,提高了高羊茅SOD酶活性。由图1C所示,在单一和混合种植模式下,多年生黑麦草和高羊茅的CAT的活性变化差异较大;无论是单一种植还是混种,添加Pb-Zn-Cd对多年生黑麦草CAT活性没有显著影响,与高羊茅单种相比(F,HF),添加Pb-Zn-Cd(PF,PHF)高羊茅的CAT活性被显著提高(P<0.05);当不添加Pb-Zn-Cd时,与单种(L,F)相比,混种(HL,HF)更为明显的降低了高羊茅和多年生黑麦草的CAT活性(P<0.05)。
Pb-Zn-Cd的添加(PL,PHL,PHF)均增加了多年生黑麦草和高羊茅的MDA的物质含量(除高羊茅单种),与单一种植模式相比(HL,HF),混种(PHL,PHF)增加了多年生黑麦草和高羊茅MDA的物质含量。当不添加Pb-Zn-Cd时,与单一种植模式(L,F)相比,混种(HL)减少了多年生黑麦草MDA的物质含量,混种(HF)显著降低了高羊茅MDA的物质含量(P<0.05)(图2)。
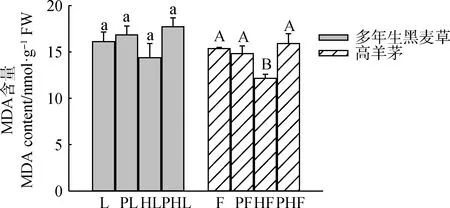
图2 不同处理对两种草坪草丙二醛含量的影响Fig.2 Effects of different treatments on MDA content in two turfgrasses注:不同小写字母表示多年生黑麦草不同处理间的差异显著(P<0.05);不同大写字母表示高羊茅不同处理间的差异显著(P<0.05)Note:Different lowercase letters indicate the differences between different treatments of perennial ryegrass at the 0.05 level. Different capital letters indicate differences among different treatments of tall fescue at the 0.05 level
2.3 不同处理对两种草坪草渗透调节物质含量的影响
从图3A可知,对单种而言(L,F),Pb-Zn-Cd的添加(PL,PF)提高了多年生黑麦草和高羊茅游离脯氨酸的营养物质含量,而混种方式下,Pb-Zn-Cd的添加(PHL)显著降低了多年生黑麦草游离脯氨酸的含量,与单种相比(PF),混种(PHF)提高了高羊茅游离脯氨酸的含量(P<0.05)。未污染处理下,与多年生黑麦草单种(L)、高羊茅单种(F)相比,两种草坪草混种(HL,HF)对多年生黑麦草和高羊茅游离脯氨酸含量没有显著影响。从图3B可知,对单种(L,F)而言,添加Pb-Zn-Cd降低了单一物种的可溶性糖含量,而添加Pb-Zn-Cd增加了混种模式下(PHL,PHF)多年生黑麦草和高羊茅可溶性糖的含量;当不添加Pb-Zn-Cd时,与单一种植模式相比(L,F),混种模式下(HL,HF),多年生黑麦草和高羊茅可溶性糖含量均降低;在添加Pb-Zn-Cd时,与单一种植模式相比(PL,PF),混种模式下(PHL,PHF),多年生黑麦草和高羊茅可溶性糖含量均显著提高(P<0.05)。由图3C可知,添加Pb-Zn-Cd(PL,PHL,PHF)会增加多年生黑麦草和高羊茅的可溶性蛋白含量(高羊茅的单一物种除外);与无污染高羊茅混种(HL)相比,添加Pb-Zn-Cd(PHL)显著提高高羊茅可溶性蛋白的含量(P<0.05)。当不添加Pb-Zn-Cd时,与单一种植模式(L,F)相比,混种模式下,多年生黑麦草的可溶性蛋白的含量增加,高羊茅的可溶性蛋白含量显著减少(P<0.05)。
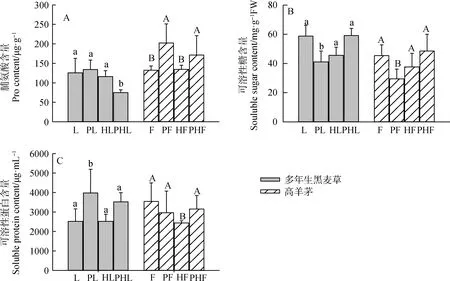
图3 不同处理对两种草坪草渗透调节物质含量的影响Fig.3 Effects of different treatments on osmotic adjustment substance content in two turfgrasses注:不同小写字母表示多年生黑麦草不同处理间的差异显著(P<0.05);不同大写字母表示高羊茅不同处理间的差异显著(P<0.05)Note:Different lowercase letters indicate the differences between different treatments of perennial ryegrass at the 0.05 level. Different capital letters indicate differences among different treatments of tall fescue at the 0.05 level
2.4 不同处理对两种草坪草巯基化合物含量的影响
从图4A可知,对于单一物种而言(L,F),Pb-Zn-Cd的添加(PL,PF)显著降低了多年生黑麦草和高羊茅Cys的含量(P<0.05),而混种模式下,(PHL)降低了多年生黑麦草Cys的含量,但未达到显著差异,(PHF)显著降低了高羊茅Cys的含量(P<0.05)。未污染处理下,相对于多年生黑麦草、高羊茅单种而言(L,F),混种(HL,HF)对多年生黑麦草和高羊茅半胱氨酸含量的影响不显著。由图4B可知,与无污染单种相比(L),Pb-Zn-Cd的添加(PL)降低了多年生黑麦草NPT的含量,与无污染混种相比(HL),Pb-Zn-Cd的添加(PHL)显著提高了多年生黑麦草NPT的含量(P<0.05),无论是单种还是混种,Pb-Zn-Cd的添加(PF,PHF)均提高了高羊茅NPT的含量。未添加Pb-Zn-Cd时,与多年生黑麦和高羊茅单一种植相比(L,F),混种模式下(HL,HF),多年生黑麦草和高羊茅NPT的含量显著减少(P<0.05)。从图4C可以看出,对于单一物种而言(L,F),Pb-Zn-Cd的添加(PL,PF)对多年黑麦草没有显著影响,但显著增加了高羊茅GSH的含量(P<0.05),而在混合种植模式下,与无污染混种相比(HL,HF),Pb-Zn-Cd的添加(PHL,PHF)均显著提高了多年生黑麦草和高羊茅GSH的含量(P<0.05)。与无污染单种相比(L,F),混种(HL)对多年生黑麦草GSH的含量无明显影响,但混种模式下(HF)高羊茅GSH的含量有所增加。
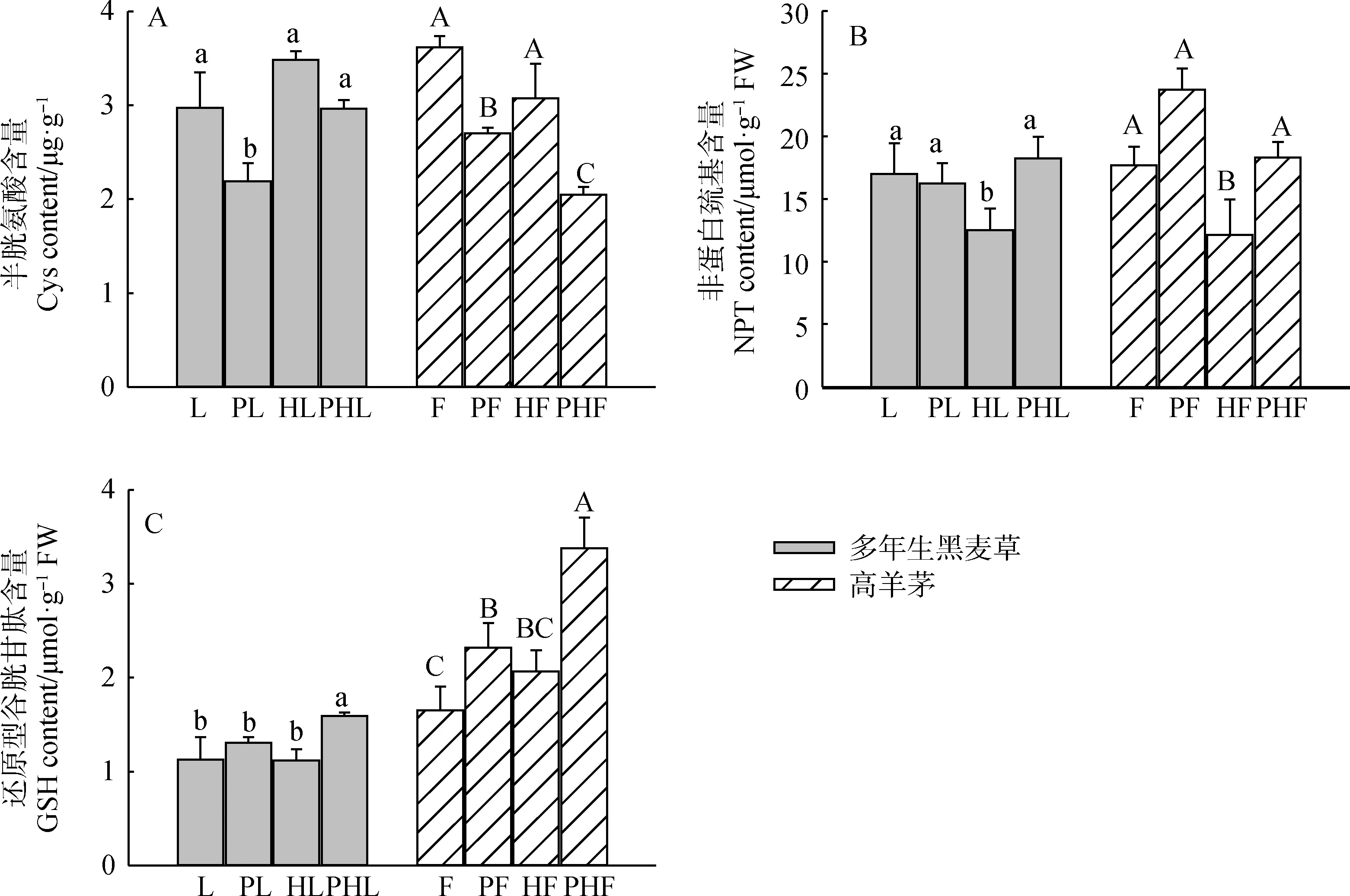
图4 不同处理对两种草坪草巯基化合物含量的影响Fig.4 Effects of different treatments on the content of sulfhydryl compounds in two turfgrasses注:不同小写字母表示多年生黑麦草不同处理间的差异显著(P<0.05);不同大写字母表示高羊茅不同处理间的差异显著(P<0.05)Note:Different lowercase letters indicate the differences between different treatments of perennial ryegrass at the 0.05 level. Different capital letters indicate differences among different treatments of tall fescue at the 0.05 level
2.5 不同种植方式对多年生黑麦草和高羊茅的耐Pb-Zn-Cd的综合评价
不同的种植方式多年生黑麦草和高羊茅的各生理指标不尽相同,为避免单独指标所带来的片面性,同时又可克服多个指标带来的复杂性,采用模糊数学隶属函数法全面而准确的评价Pb-Zn-Cd复合污染对不同种植方式下多年生黑麦草和高羊茅生理指标的影响,对多年生黑麦草和高羊茅的叶绿素a,叶绿素b,类胡萝卜素,叶绿素,光合色素,POD,SOD,CAT,MDA,Pro,可溶性糖,可溶性蛋白,Cys,NPT,GSH进行隶属函数值计算,得出不同种植方式对多年生黑麦草和高羊茅的耐Pb-Zn-Cd的综合评价值(表3)。由综合评价得出多年生黑麦草各个处理的耐受性为:无污染单种>污染混种>污染单种>无污染混种;高羊茅各个处理的耐受性为:污染单种>无污染单种>污染混种>无污染混种。Pb-Zn-Cd污染下,多年生黑麦草和高羊茅的耐受性比较:单种时,高羊茅>多年生黑麦草;混种时,多年生黑麦草>高羊茅。
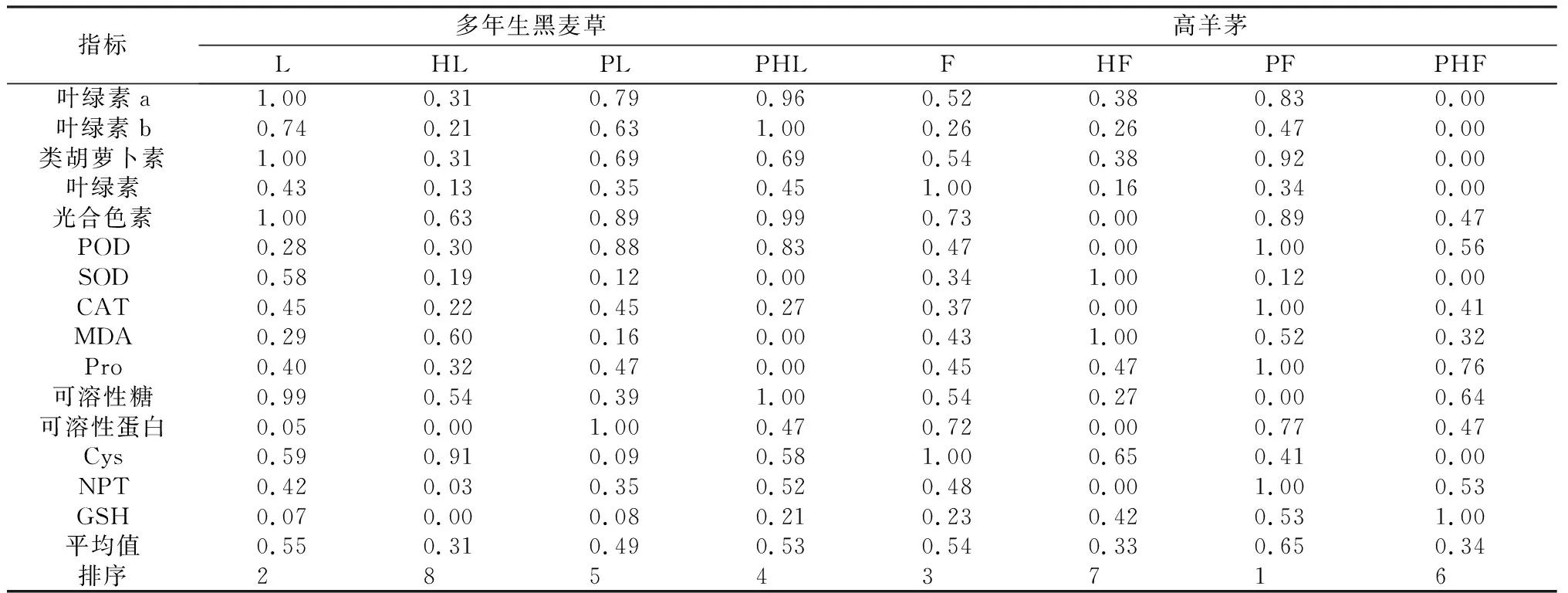
表3 多年生黑麦草和高羊茅的不同种植方式耐Pb-Zn-Cd的综合评价值Table 3 Comprehensive evaluation values of Pb-Zn-Cd tolerance of perennial ryegrass and tall fescue in different planting ways
3 讨论
3.1 重金属污染对植物光合色素含量的影响
植物光合作用是植物获取能量的重要途径,植物能量含量与光合作用之间有着非常密切的关系。重金属胁迫下,植物的叶绿素含量会显著降低,叶绿体膜结构会受到一定程度的损伤,抑制叶绿体内的光合酶合成,导致植物叶片气孔关闭,使光合作用的强度降低[14]。前人研究显示,在重金属Pb2+的胁迫下,多年生黑麦草的叶绿素含量显著高于对照组,而在重金属Cd2+的胁迫下,多年生黑麦草的叶绿素含量呈现出明显的下降趋势[15];在Pb,Zn,Cd,Pb-Zn和Cu-Cd胁迫下,多年生黑麦草和高羊茅的叶绿素含量都呈显著下降趋势,在Cu单一胁迫下,多年生黑麦草和高羊茅的叶绿素含量呈上升趋势[16]。在Zn2+和Mn2+的胁迫下,两种禾本科植物的叶绿素a含量和叶绿素b含量以及叶绿素a /b的比值比对照组均显著降低[17]。本研究中,Pb-Zn-Cd污染下,提高了高羊茅叶绿素a、叶绿素b、类胡萝卜素、叶绿素以及光合色素的含量,降低了多年生黑麦草叶绿素a、叶绿素b、类胡萝卜素、叶绿素以及光合色素的含量,未达到显著差异。可能是因为在该浓度Pb-Zn-Cd复合污染下,能够促进高羊茅植物体内光合色素相关酶的合成,多年生黑麦草对该浓度Pb-Zn-Cd复合污染下的耐受性较差,部分与光合色素合成的相关酶受到抑制,也有可能是Pb-Zn-Cd的复合胁迫致使多年生黑麦草叶片中的Fe2+含量下降,加上Pb-Zn-Cd自身对叶绿素的新陈代谢具有消极影响,可能会导致叶绿素的降解受到抑制、生物合成受阻。Pb-Zn-Cd污染下,混种提高了多年生黑麦草叶绿素a、叶绿素b、类胡萝卜素、叶绿素以及光合色素的含量,降低了高羊茅叶绿素a、叶绿素b、类胡萝卜素、叶绿素以及光合色素的含量,均未达到显著差异。可能是因为这种混种模式可以促进多年生黑麦草植物体内光合色素相关酶的激活和合成,但却抑制了高羊茅植物体内光合色素相关酶的激活和合成。
3.2 重金属污染对植物抗氧化酶系统的影响
氧化损伤是重金属胁迫下植物的重要作用机制。在重金属胁迫下,植物中的活性氧物种(Reactive oxygen species,ROS)将过度产生和积累[18],高水平的ROS一般会破坏细胞的辅助成分,如细胞膜、核酸和叶绿素等[19],造成氧化损伤,影响植物正常的生长发育。低浓度的重金属会激活植物的抗氧化管理系统,诱导SOD,POD和CAT的抗氧化蛋白酶活性增加,并去除植物中的活性氧物种[20],抗氧化酶的诱导性是减少氧化变色损伤的重要保护运行机制,其中SOD在诱导下降解增加,催化超氧阴正离子转化为过氧化氢和氧气;过氧化氢也是自由基的一种,它也会对细胞膜造成氧化损伤,POD和CAT可以催化过氧化氢产生无毒的水和氧气分子,使植物体内的自由基保持在一个较低水平,避免自由基对植物造成的氧化损害,过氧化氢酶是植物在逆境下代谢的关键保护酶[21]。HU等人[22]研究发现,在浓度为1 000 mg·L-1的Pb胁迫下,高羊茅的SOD和POD活性均增加。对14份禾本科牧草的耐铅性研究发现,在重金属铅胁迫下,植物的POD和SOD活性均呈先上升后下降的趋势,CAT活性整体呈下降趋势[23]。在本研究中,在Pb-Zn-Cd污染下,多年生黑麦草和高羊茅的POD活性均增加、高羊茅的CAT活性也增加、多年生黑麦草和高羊茅的SOD活性降低;与单一种植模式相比,混种模式下,多年生黑麦草和高羊茅的SOD活性成分均有增加,高羊茅CAT活性显著增加(P<0.05),但对高羊茅的POD活性和多年生黑麦草的POD和CAT活性没有显著影响。与Alonso-Blázquez等[24]研究结果不同。在重金属污染下,SOD的活性降低可能是因为在该浓度的Pb,Zn,Cd复合胁迫和混种条件下,植物体内所有的活性氧清除酶系统和具生理抗性特征的生理活动被抑制,SOD的活性在此条件下会逐渐被降低,从而导致了植物体内过多的超氧阴离子自由基得不到很好的清除,Pb-Zn-Cd胁迫产生的超氧阴离子没有被SOD很好的清除,在清除自由基的这个过程中,抗氧化酶系统之间存在协同作用,当SOD酶活性降低时,CAT和POD的活性就会升高。
MDA是膜脂质过氧化的最终产物,在外界环境的刺激下,生长中的植物会失衡膜脂的过氧化程度,产生大量不利于植物生长的活性氧。实际上,MDA含量通常被用作衡量植物根系在胁迫下受损程度的综合指标[25]。本研究的结果显示,无论是重金属污染还是在没有污染情况下,混种模式对多年生黑麦草和高羊茅的MDA含量没有显著影响,与于保港等[26]对莎草/蚕豆间作的科学研究结果相似。在Pb-Zn-Cd未污染下,与单种相比,混种对多年生黑麦草和高羊茅的MDA含量的影响不显著,混种多年生黑麦草和高羊茅的MDA物质含量低于单种,可能是在混种下植物细胞膜脂质过氧化水平减小,植株膜透性减小,进而MDA含量下降;在Pb-Zn-Cd污染下,混种对多年生黑麦草和高羊茅的MDA含量的影响与单种相比不显著,混种多年生黑麦草和高羊茅的MDA含量高于单种。MDA含量增加是植物膜质过氧化水平加剧的表现,在Pb-Zn-Cd胁迫下,虽然抗氧化酶一定程度的增加起到保护作用,但是多年生黑麦草和高羊茅植物体的细胞膜仍然会受到伤害,从而导致细胞膜会受到一定的损伤。
3.3 重金属污染对植物渗透调节物质的影响
脯氨酸是一个重要的渗透调节物质,在逆境胁迫中,游离氨基酸的合成得到加强,游离脯氨酸通过渗透作用调节植物细胞,从而提高植物抵抗逆境的能力,保护自身机制免受最大程度的伤害[27]。在重金属胁迫下,由于重金属离子的存在,外部渗透势降低,植物细胞内的水分以低渗透势低流失到外部,造成植物缺水和常见的植物损害。脯氨酸含量的增加能够降低植物细胞内的渗透势,提高植物的持水能力,从而避免缺水对植物赞成的损害[28]。刘圆等人在3种藜科植物对复合重金属污染的生理响应的研究中,发现在不同浓度金属的物和胁迫下,游离脯氨酸的含量降低[27]。Pb2+浓度在1 000 mg·L-1以下,多年生黑麦草游离脯氨酸(Pro)含量呈上升趋势,Pb2+浓度超过1 000 mg·L-1,多年生黑麦草游离脯氨酸(Pro)含量呈下降趋势[29]。这表明,重金属浓度不同,植物体内渗透调节物质的变化也随之不同。本研究中,对单种而言,Pb-Zn-Cd的添加提高了多年生黑麦草和高羊茅游离脯氨酸的含量,这表明,在Pb-Zn-Cd污染单种模式下,使植物体内水分亏缺严重,两种草坪草通过合成大量的游离脯氨酸来提高两种草坪草的保水能力,从而缓解重金属胁迫对植物造成的伤害。而混种方式下,Pb-Zn-Cd的添加显著降低了多年生黑麦草游离脯氨酸的含量,提高了高羊茅游离脯氨酸的含量,这表明混种增加重金属对多年生黑麦草的胁迫作用,加之混种增加了对多年生黑麦草细胞膜系统的破坏,因而刺激植物生成的脯氨酸减少,但本研究中,混种对高羊茅脯氨酸的合成没有显著的影响。
可溶性蛋白可以参与植物细胞的渗透调节,从而提高植物的抗逆性,它不仅是植物筛选抗性的指标之一,也是了解植物体总代谢的重要指标[30]。在植物中,可溶性蛋白质主要是不与膜系统特异结合的酶,可溶性蛋白含量越高,其生理生化反应和代谢活动越强。一般来说,在不利条件下,由于蛋白质分解的加速,蛋白质的合成受到抑制,植物中蛋自质的含量降低[31]。何冰等人[32]发现,在6 mmol·L-1的高铅浓度处理下,杨梅叶片可溶性蛋白的含量表现出显著下降趋势,这显著抑制了杨梅叶片的代谢活性。本研究显示,无污染下,混种降低了高羊茅的可溶性蛋白含量,而多年生黑麦草的可溶性蛋白含量变化不明显,可能是在混种下,高羊茅表现出混种劣势;在Pb-Zn-Cd污染下,混种对两种植物的可溶性蛋白含量没有显著影响。
可溶性糖是一种调节细胞渗透压力的小分子物质,它在维持细胞扩张、酶活性、维持细胞膜系统稳定性和植株光合作用等方面起着重要的生理作用[33]。植物可以通过在逆境中提高可溶性糖以及其他有机物质的含量,从而细胞的渗透压得到调节,植物适应逆境的能力就会得到提高[34]。孙小霞[35]认为高浓度的铅促使高羊茅可溶性糖的增加;李珊等[36]的研究发现随着铅浓度的升高,栝楼(Trichosantheskirilowii Maxim.)幼苗体内可溶性糖含量逐渐下降,因此,重金属胁迫下不同种植物体内的可溶性糖是复杂多变的。本研究显示,在Pb-Zn-Cd未污染下,与单种相比,混种多年生黑麦草和高羊茅的可溶性糖含量下降,可能是因为混种刺激植物生成的渗透调节物质减少;在Pb-Zn-Cd污染下,多年生黑麦草和高羊茅在混合种植模式下的可溶性糖含量较单一种有所增加,这可能是由于Pb-Zn-Cd胁迫下植物的淀粉水解增加、光合产物产量缓慢以及水分亏缺导致的碳水化合物积累所致。
3.4 重金属污染对植物巯基化合物含量的影响
Cys是无机化合物硫酸盐同化的初始产物,也是谷胱甘肽合成的限速因子,对Cd来说,Cys的合成是因为它参与了植物的解毒过程。在本研究中,在Pb-Zn-Cd复合胁迫下,无论是单一种植模式还是混种模式,多年生黑麦草和高羊茅中Cys的含量均降低。这可能是因为该浓度的Pb-Zn-Cd复合胁迫降低了两种草坪草对金属离解的螯合能力,削弱了两种草坪草对重金属胁迫的耐受性,这与韩淑梅[37]和其他人在孔雀草的研究结果不一致。巯基化合物在植物的抗金属胁迫中有重要作用,由细胞中的各种金属和准金属诱导能促使NPT的合成,其防御金属毒性作用主要通过螯合金属离子来实现[38]。本研究中,无论是单种还是混种,Pb-Zn-Cd的添加均提高了多年生黑麦草和高羊茅NPT的含量,这表明重金属处理能够诱导多年生黑麦草和高羊茅NPT的合成。有研究表明,NPT含量的增加促进了低分子量巯基化合物的合成,如GSH和植物络合素(PCs),这些物质通过金属硫醇的形式参与重金属的解毒过程[39]。GSH可以保护体内重要的植物免受氧化和失活。它们具有抗氧化、整合和解毒作用,共同参与清除植物毒素[40]。本研究中,对于单种而言,Pb-Zn-Cd的添加提高了高羊茅GSH的含量,而混种模式下,均提高了多年生黑麦草和高羊茅GSH的含量,说明在重金属胁迫下,刺激了高羊茅GSH的合成,混种能刺激多年生黑麦草GSH的合成,GSH合成为植物PCs的合成提供合成前体,PCs通过与重金属络合保护植物免受毒害。
4 结论
在Pb-Zn-Cd复合胁迫下,多年生黑麦草和高羊茅单种与混种表现出一定的耐受性,根据多年生黑麦草和高羊茅单种与混种(1∶1)苗在Pb-Zn-Cd复合污染下的生理响应和耐受性综合评价,得出结论,在Pb-Zn-Cd复合胁迫下,高羊茅单种具有较好的抗逆性,多年生黑麦草和高羊茅1∶1混种使多年生黑麦草有较好的抗逆性。
猜你喜欢
奥秘(创新大赛)(2022年8期)2022-09-14
中国果业信息(2021年10期)2021-12-07
昆明医科大学学报(2021年8期)2021-08-13
中国饲料(2021年8期)2021-05-17
阅读(科学探秘)(2020年8期)2020-11-06
新农业(2019年9期)2019-09-17
绿色科技(2019年2期)2019-05-21
女性天地(2016年10期)2017-04-25
农家顾问(2016年7期)2016-05-14
医学研究杂志(2015年9期)2015-07-01
