
种子引发提高草类植物抗旱性的表现及机理
2022-11-04 09:12石晓琪米素娟钟天航韩云华
草地学报 2022年10期
石晓琪, 米素娟, 钟天航, 韩云华
(草地农业生态系统国家重点实验室 兰州大学草地农业科技学院, 甘肃 兰州 730020)
干旱在各种非生物胁迫中破坏性最强,是导致全球作物减产的主要原因之一。我国受旱灾影响的耕地面积占总受灾耕地面积的26%[1],每年由于干旱导致的粮食损失占到总产量的17%以上[2]。水分参与植物生命活动的全周期,干旱使植物可利用的水分减少,对植物形态、生理代谢、品质产量等方面产生不利影响,导致植物生长发育受阻[3-5]。
我国耕地面积有限,草类植物种植生产主要利用内蒙古、新疆、甘肃等干旱半干旱地区沙化、盐碱化较严重的边际土地,更易受到干旱的威胁[6]。生产实践中可通过抗旱育种、改善灌溉条件、种子处理等方式提高草类植物生产性能。种子引发是种子处理的一种,在提高植物抗旱性方面具有良好的效果和经济性[7]。以往种子引发相关综述多是关于作物、蔬菜抗旱方面的研究[7-8],关于种子引发在草类植物抗旱应用方面的综述还不多见。本文总结了种子引发技术在草类植物抗旱方面的影响及种子引发提高抗旱能力的机制,以期为种子引发在草类植物生产中的应用提供参考。
1 种子引发技术的定义和方法
1.1 种子引发技术
种子引发由Heydecker于1973年提出[9],通过渗透调节来控制种子缓慢吸水达到预发芽状态,有助于提升种子的萌发质量和抗逆性。种子引发技术的关键在于控制种子“萌而未发”,引发使种子达到预萌发状态,生理代谢活动激活,而胚根、胚芽不会突破种皮发芽[10]。通过引发处理使种子吸胀的缓慢吸水阶段被无限延长,为种子赢得足够的时间使种子的胚为萌发作好充分准备[11](图1)。
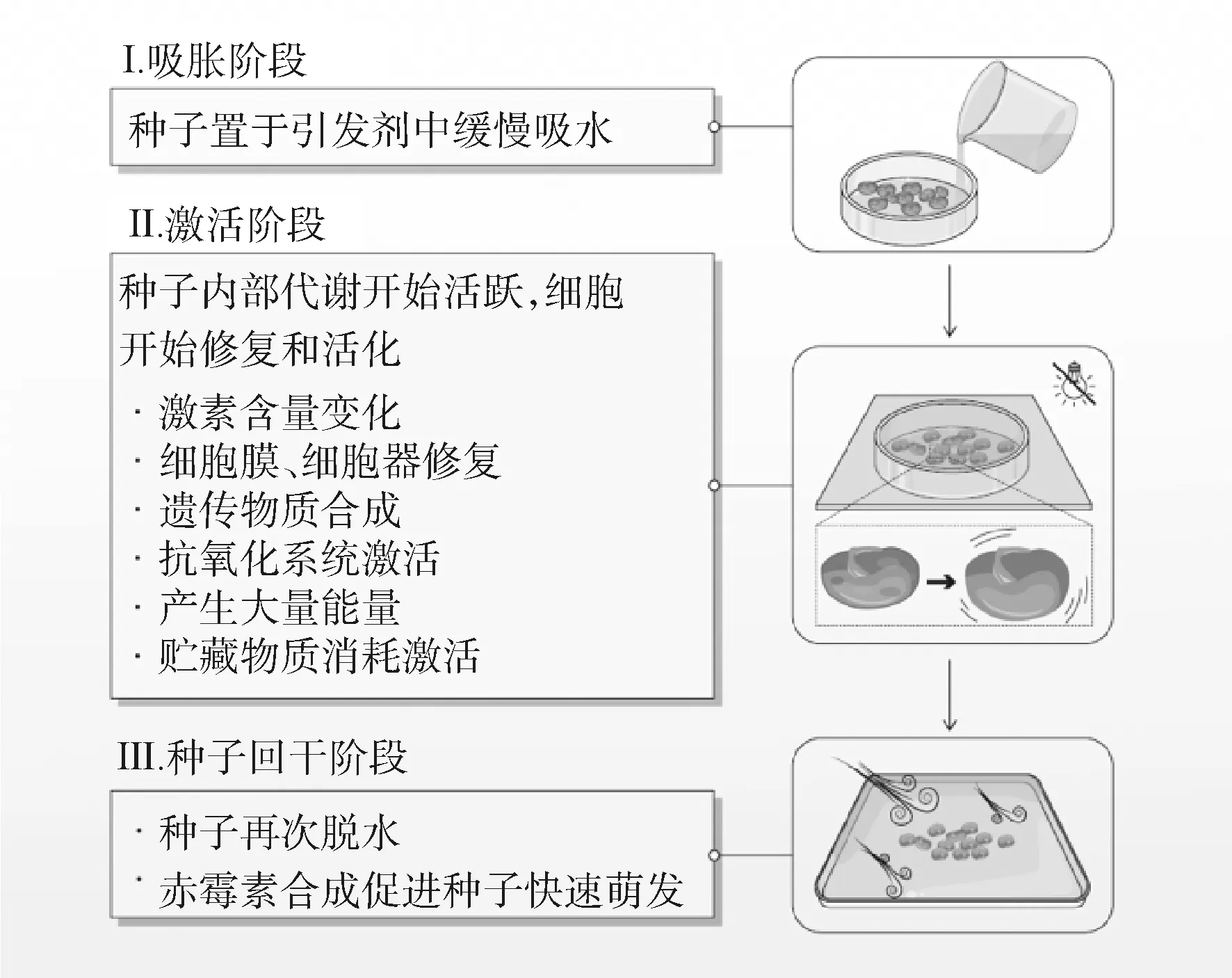
图1 种子引发过程Fig.1 Seed priming process
1.2 种子引发方法
种子引发有多种方式。水引发(Hydro-priming)通过调节给水条件,控制种子吸水,避免吸胀伤害,实现促进萌发的目的[12]。水引发技术具有简捷便宜、无毒无害、易推广和易应用的优势,但也存在吸湿不均,微生物过量繁殖的问题,导致种子感染或失活死亡[13]。渗调引发(Osmo-priming)是通过控制溶液水势,实现种子定量吸水的引发技术。常用溶液包括KCl,NaNO3,NaCl,KNO3,聚乙二醇(PEG)、甜菜碱等。引发溶液可以是单一药剂,也可以是多种药剂混合。固体基质引发(Solid matrix priming)是将种子、引发液与固体基质以一定比例混合,使种子胶体缓慢吸胀。常用固体基质有沙子、蛭石、聚丙烯酸钠胶等。生物引发(Bio-priming)指利用有益微生物作为引发剂,抑制病原微生物,分泌植物激素等,促进种子萌发。激素引发(Hormonal priming)是使用赤霉素(Gibberellic acid,GA)、脱落酸(Abscisic acid,ABA)、油菜素内酯(Brassinolide,BR)等激素浸种,提高植物抗逆性和发芽率的引发方法[14]。物理引发是利用如磁场、超声波等物理介质处理种子的引发方法[15]。
近年来,纳米颗粒植物互作的研究突飞猛进,多项研究表明,纳米颗粒够促进植物生长、防治植物病害等。纳米引发(Nano-priming) 也作为一种新的引发技术而受到大家关注。该引发技术可以使种子在贮藏过程中维持较高的种子活力,增强种子萌发抗逆能力,提高萌发率、幼苗整齐度,并且激活了萌发过程中与植物抗逆性表达的相关基因[16-17]。
除引发剂自身化学特性的差异外,不同引发方式控制种子吸水的速度也各不相同。一般来说,固体基质引发能更好的控制引发用水量,避免种子吸胀损伤。控制引发液水势也有类似效果。此外,种子回干的时间、温度等条件同样影响引发结果[18]。为取得理想引发效果,需要选用适宜的引发方法并控制回干条件。
2 种子引发对草类植物抗旱性的影响
表1总结了种子引发在草类植物中的研究,各种引发方法均有良好的抗旱效果,具体表现为促进萌发、提升幼苗生长性能、改善田间表现等。
2.1 引发促进干旱条件下种子萌发
种子萌发和幼苗阶段对水分最敏感。缺水推迟发芽并降低最终发芽率和发芽整齐度,导致作物生产性能下降[19]。种子引发可提高多种草类植物种子萌发和出苗抗旱性。田雨等发现渗透引发降低了紫花苜蓿(Medicagosativa)种子发芽对水势的需求,使种子快速发芽,提高了在发芽阶段的抗旱能力[20]。水引发提升了干旱胁迫下沙棘(Hippophaerhamnoides)种子的萌发特性,未引发时种子干旱临界萌发条件为-0.6 MPa,引发后提升为-0.9 MPa,沙棘的抗旱适应性增强[21](表1)。在干旱条件下,用0.40%~1.20% NaCl引发处理的夏枯草(Prunellavulgaris)种子,用赤霉素引发白花蛇舌草种子(Oldenlandiadiffusa),均能够提高种子的发芽率、发芽指数、活力指数并促进幼苗的生长[22-23]。PEG引发可提高无芒雀麦(Bromusinermis)、苇状羊茅(Festucaarundinacea)、羊茅(Festucaovina)种子的发芽率[24]。甘氨酸甜菜碱引发显著提高了干旱条件下草地早熟禾(Poapratensis)、狗牙根(Cynodondactylon)的发芽率[25]。水引发和尿素引发长穗偃麦草(Agropyronelongatum)种子,可以改善其种子在干旱胁迫下的发芽指数[26]。
种子引发还可提高低活力种子萌发的抗旱性。干旱条件下,激素引发提高了长穗偃麦草老化种子的发芽率和发芽速率,50 ppm的生长素引发使自然老化的种子的发芽率提高了18%,细胞分裂素处理提升了种子活力指数[27]。
2.2 引发改善草类植物抗旱性和田间表现
水分胁迫导致幼苗根系代谢紊乱,部分功能丧失,活力减弱,根系生长被抑制[28-29]。研究发现,甜菜碱、甘露糖、亚精胺引发提高了干旱胁迫下白三叶(Trifoliumrepens)根系活力,改善了根的吸水能力,显著增强了幼苗的抗旱能力[30-32]。田新会发现,黄腐酸引发苜蓿显著提升干旱下幼苗的根长、根粗,增强幼苗抗旱能力[33]。由此可见,种子引发对苗期抗旱性能提升具有重要作用。
种子引发可显著改善干旱情况下草类植物的田间表现。陶奇波研究发现用水、PEG和亚精胺引发使干旱环境下无芒隐子草温室和田间的出苗率分别提升4%~17%和9%~19%[34]。PEG(-0.6 MPa)和抗坏血酸(AsA)引发苜蓿种子提高植物在缺水条件下的株高、生物量等[35-36](表1)。闵丹丹发现用PEG和KNO3使紫花苜蓿在干旱条件下的田间生物量提升53%~57%[37]。
3 种子引发提高草类植物幼苗抗旱性机制
如图2所示,干旱胁迫影响细胞中水分的自由度,种子内活性氧(Reactive oxygen species,ROS)大量积累。过量的ROS使细胞脂质和各种膜结构过氧化受损,细胞内溶质外泄。脂质过氧化同时导致丙二醛(Malondialdehyde,MDA)含量增加,细胞膜进一步受到破坏,透性加剧,渗出液电导率上升[38-40]。活性氧自由基能直接与核酸作用,影响核酸结构和DNA复制,最终导致基因表达受阻,细胞功能受损[41-42]。
种子引发能够激活或改变干旱胁迫下植物的形态结构、生理生化过程和分子调控途径(表1),有助于草类植物幼苗耐旱能力的提升(图3)。

图2 植物对干旱的响应(根据文献[38-42]以及表1总结绘制)Fig.2 Plant response to drought stress(plotted from references[38-42] andTable 1)
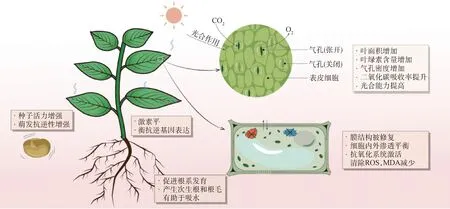
图3 种子引发对植物抗旱性的影响(根据文献[43-84]以及表1总结绘制)Fig.3 The effect of seed priming on plant drought resistance(plotted from references[43-84] and Table 1)
3.1 形态机制
根是吸收土壤水分、感受外部刺激的重要部分,当土壤缺水时根系迅速向植株发出信号,同时调整根系形态及数量[43]。增加根冠和根芽比例使植物在干旱胁迫下能够最大限度地吸收水分,并从土壤中汲取营养物质,从而适应干旱环境,增强植物早期发育能力[44-45]。研究发现,引发处理能提升植物幼苗的根冠比、根芽比、平均根重,侧根数量和根长[46-47],改善植物抗旱性。
植物角质层能够减轻植物非必要水分蒸腾并且抵御外界不良环境,与植物抗旱性紧密相关[48]。研究发现,水引发和2,4-表油菜素内酯引发可以提高幼苗气孔导度,显著增加角质含量,减少蒸腾耗水[49],提高植物水分自由度。
3.2 生理生化机制
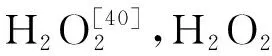
3.2.2促进细胞膜修复 种子引发能够促进细胞膜的修复与活化,改变细胞膜离子通透性。例如,用GA3引发青绿苔草(Carexleucochlora)种子,能够修复细胞膜,改变其离子通透性[53](表1)。引发延长了种子萌发的激活阶段,使种子有充分的时间修复膜损伤[54]。植物遭到水分胁迫后,细胞膜的透过性下降,导致外界非必要元素进入细胞内,同时细胞液和离子外渗。通过引发可以恢复细胞膜离子的选择透过性,维持细胞正常渗透压,因此增强植物的抗旱能力[14]。
3.2.3改善有机物渗透调节能力 干旱胁迫下,植物适应逆境的能力与植物细胞渗调能力有关[55]。植物受到水分胁迫后,虽通过调节自身生理生化过程积累一些渗调物质,但含量较少仍不足以抵抗胁迫导致的损害[56]。种子引发处理后,一方面能够有效提高脯氨酸、甘露醇、甜菜碱、可溶性糖等有机物质含量,有助于维持细胞渗透压,调节植物生理过程,以应对胁迫损伤[57-59];另一方面,这些有机物质本身具有一定的生理活性,如可溶性糖的积累有利于清除超氧阴离子、减轻氧化毒害、维持叶片相对含水量和保持叶片气孔开放[60-61];脯氨酸在维持膜完整性、清除ROS以及维持光反应中心电子流等生理生化过程中发挥着重要作用,这些过程与植物耐旱性密切相关[62]。研究发现,用水引发苜蓿能够提高种子脯氨酸含量[63],有助于改善幼苗的抗旱表现(表1)。
3.2.4提高代谢酶活性 种子内贮藏的淀粉经淀粉酶水解成小分子糖类,既为种子萌发提供能量,又使渗透物质增多,维持了细胞渗透压,增强种子抗逆性[64-65]。研究发现甘露糖预处理白三叶种子,提高了其在干旱胁迫下淀粉酶的活性,减轻干旱胁迫导致的淀粉代谢抑制[31]。此外,引发还可通过提升醛缩酶、葡萄糖-6-磷酸脱氢酶、磷酸脂酶、酯酶、乙醇脱氢酶、β-甘露糖酶、异柠檬酸裂解酶、DNA修复酶等酶的活性[66],从而提升植物抗旱能力。
3.2.5调节激素含量 研究发现一些内源激素的含量与植物抗逆基因的表达相关[67]。这些内源激素构成植物细胞内特定的防御信号途径[60]。ABA是调节植物的生长发育的重要激素之一,可通过调节气孔的运动,使植物组织内的水分保持平衡,通过增强根的通透性,提高水的可吸收性,使得植物的抗逆性得到提升[68]。拟南芥(Arabidopsisthaliana)种子用非蛋白氨基酸b-氨基丁酸引发后,通过ABA的积累、响应胁迫基因的表达和关闭气孔等方式,提高了对水分胁迫的抗性[69]。杜利霞等研究发现,用6-BA引发赖草(Leymussecalinus)种子能够显著提升ABA,GA含量,加速种子代谢,促进种子萌发[70]。
3.2.6改善幼苗光合特性 光合作用作为植物不可缺少的基础生理过程,直接影响生物产量。光合作用对干旱环境高度敏感,在干旱条件下,光合作用受到抑制,表现为叶绿素含量、净光合速率、气孔导度、蒸腾速率、和细胞间CO2浓度等降低[49],光合作用受阻,植物生长发育不良[28,71]。研究表明,通过纳米铁引发柳枝稷(Panicumvirgatum)能够提高干旱胁迫下其幼苗的光合特性,使幼苗的光合效率提升,抗氧化系统能力增强,减轻干旱胁迫对柳枝稷的损伤[72]。
3.3 分子机制
种子引发有利于遗传物质修复和诱导相关抗旱基因表达,促进了细胞活动周期的恢复,促进了抗旱相关蛋白的合成。同时引发使植物留下“胁迫记忆”,有助于之后快速应对类似胁迫。
3.3.1提高DNA复制效率 成熟种子中存在处于不同分裂时期的细胞[73]。干旱胁迫细胞产生一些信号分子,抑制G1期到S期的过渡,减缓有丝分裂中的DNA复制[74-75]。种子引发能够加快细胞核DNA复制,推动细胞从G1期向S或G2期转化,将细胞周期推进到有丝分裂的阶段[76-77],缓解了干旱的不利影响。无芒隐子草经过引发处理,显著提高了G2期的细胞比例,即DNA复制效率明显提高,有效增强了无芒隐子草种子的活力,有助于改善其田间出苗情况[34]。
3.3.2诱导抗逆关键基因表达 种子引发可以调节多个应激反应途径的基因表达和蛋白质表观修饰,提高对恶劣环境的耐受性[40]。水通道蛋白是一种细胞膜上的蛋白质,在细胞膜上形成小孔,控制水分在细胞上的进出,对种子和植株抗旱胁迫过程的水分运输有重要作用[78]。种子经引发处理后,水通道蛋白基因BnPIP1的表达被激活[79]。干旱条件下,2,4-表油菜素内酯引发处理中植物幼苗角质相关基因的表达量是对照的3倍[49]。
在干旱胁迫下,硝普钠(100 μmol·L-1)引发烟草(Nicotianatabacum)种子处理中脯氨酸合成关键酶P5CS的基因表达量均有不同程度的提高,提高了脯氨酸含量[60]。拟南芥种子引发后,DNA糖苷酶AtOGG1基因表达上调,有助于消除DNA病变,提高对非生物胁迫的耐受力[80]。水引发蒺藜苜蓿(Medicagotruncatula)种子使SOD基因与APX基因上调,有助于提高幼苗耐受力[81]。
3.3.3胁迫记忆 引发处理使种子萌发时期处于较温和的胁迫中,使植物萌发后留下“胁迫记忆”,使植物再遇到类似胁迫可以快速反应或更好的应对[82]。引发作为一种轻度胁迫,能够提前激活部分抗逆基因和蛋白质,抗逆能力在种子回干后被保留[83]。种子预先暴露在胁迫下,能改变其后续表现,拟南芥植物表现为转录速率增加和逆境反应基因(RD29B和RAB18)的转录水平提高[84]。

表1 种子引发对植物抗旱的影响Table 1 The effects of seed priming on plant drought resistance

续表1
4 问题与展望
近年来,种子引发研究进展迅速,许多新引发技术应运而生。随着种子引发技术的研究不断深入,其机理揭示也愈发清晰。但目前在草类植物引发研究中也存在着一些问题。相较作物、蔬菜等,种子引发技术在草类植物上的研究重视程度不足。种子引发效果在种子种类、品种、种子批次间存在差异[93],草类植物种类繁多,尚未有可以大规模商业化应用的种子引发技术。关于种子引发后种子回干方面的研究较少。种子回干条件的不同,如回干时间、回干温度等,会对种子理化过程产生不同变化,从而影响引发效果;引发后种子的贮藏寿命研究不够充分,影响其实际应用效果。现有研究多集中在室内种子的萌发阶段与幼苗阶段,对引发后田间效果的检验及后续产量变化方面关注不够,研究内容也以引发后植物形态和生理的变化为主,而缺少关于分子层面的研究。
随着对山水林田湖草沙共同体理念深入人心,种子引发技术在草类植物中的研究势必更加充分,在降低引发成本的同时,改善引发效果。同时分子生物学和生物信息学等技术不断深入发展,基因组学、转录组学、蛋白质组学、表型组学等组学手段结合种子引发方面的研究也逐步深入,有助于进一步解析种子引发技术提高草类植物抗旱性的内在机制。
猜你喜欢
作物研究(2022年4期)2022-09-21
今日农业(2021年10期)2021-11-27
今日农业(2021年14期)2021-11-25
落叶果树(2021年6期)2021-02-12
今日农业(2020年13期)2020-12-15
种子(2020年11期)2020-12-05
中国生态农业学报(中英文)(2020年2期)2020-02-13
建材发展导向(2019年11期)2019-08-24
中国火炬(2015年12期)2015-07-31
中国火炬(2013年3期)2013-07-24
