
秦岭石生苔藓结皮的微生物群落组成和多样性特征
2022-11-09 12:01周雯娟卜崇峰韦应欣
西北植物学报 2022年9期
周雯娟,卜崇峰,2*,韦应欣
(1 西北农林科技大学 水土保持研究所,陕西杨陵 712100;2 中国科学院 水土保持研究所,陕西杨陵 712100)
生物结皮是旱区生态系统的重要组成部分[1-2],苔藓结皮则是生物结皮演替的高级阶段[3-4]。苔藓结皮是由苔藓及其浓密的假根粘结土壤颗粒而形成的一种地表覆盖物[5-8],由于其独特的生理结构和功能而受到广泛关注。国内对西北沙漠地区、黄土高原地区[9-12]等干旱缺水地区的土生苔藓结皮的研究已经相当深入,对其生态效应及物种组成都有一定的了解。苔藓结皮可以保持水土[13-14]、促进种子萌发和维管植物的生长;苔藓结皮将大气中的N2固定或还原成可利用的氮素形态,是荒漠生态系统氮素的主要贡献者。Russow等[15]认为内盖夫沙漠中生物结皮的年固氮量在10~41 kg/(hm2)。微生物群落是苔藓结皮发挥生态功能的主要成分,在物质循环和能量流动过程中发挥着重要作用[16-18]。
但是,受气候、土壤、植被、地形地貌等诸多因素的影响[19-20],不同地区苔藓结皮微生物物种组成及丰度也有所不同[21-22]。黄土高原苔藓结皮的优势细菌门包括酸杆菌门(24.3%)、变形杆菌门(23.8%)、绿弯菌门(15.8%)和放线菌门(14.5%),真菌群落以子囊菌门(68.0%)和担子菌门(23.8%)为主[23]。放线菌门、变形菌门、酸杆菌门、蓝细菌门、拟杆菌门等则是毛乌素沙地苔藓结皮层常见的细菌菌门类群,真菌群落以子囊菌门和担子菌门为主。
根据生境的不同,苔藓分为水生苔藓、石生苔藓、土生苔藓和木生苔藓4种类型[24]。石生苔藓生于岩石表面,常形成丛生状或垫状群落[25],具有吸水快、蓄水量大的特点[26],是裸露岩石上的先锋生物,苔藓生长过程中分泌的有机酸促使岩石风化,并且释放养分[27-28],为其他植物的着生提供条件。秦岭是中国中部重要的植被覆盖区[29],大量的地表缺少土壤、岩石裸露,由于降水丰沛、气候适宜,石生苔藓结皮发育非常广泛。遗憾的是,已有研究多集中在旱区的土生苔藓结皮,有关秦岭湿润区石生苔藓结皮及其微生物的研究报道还寥寥无几,这极大地限制了对于其发育机制和生态功能发挥机理的揭示。
为此,本研究针对秦岭山脉北麓的石生苔藓结皮,利用高通量测序技术,分析其微生物的群落组成和多样性特征;并测定其养分状况,深入探讨了养分指标与微生物群落的关系。研究结果有助于了解湿润地区苔藓结皮的养分状况和微生物群落组成,为探究湿润区苔藓结皮在岩石上的附着、风化机制提供理论基础,为后续室内培育和野外恢复提供指导意见。
1 材料和方法
1.1 研究区概况
研究区位于秦岭北麓周至县的泥峪、骆峪和车峪(图1),地势呈西南高东北低,山区占76.4%,岩石多为深变质的片岩及火成岩。区内河流众多、河床陡峻、水流湍急,植物资源丰富。属于温带大陆性季风气候,低山区年温差小,年均温10 ℃左右。气候湿润,多年平均降水量为850.52 mm,由南向北递减。夏季多暴雨,秋冬多阴雨,洪涝灾害比较频繁。采样点都临近溪流,空气湿度大,植被茂盛,羽枝青藓[Brachytheciumplumosum(Hedw.) B.S.G.]、匐灯藓(PlagiomniumcuspidatumT.Kop.)和鳞叶藓[Taxiphyllumtaxirameum(Mitt.) Fleisch.]是研究区的优势苔藓种。
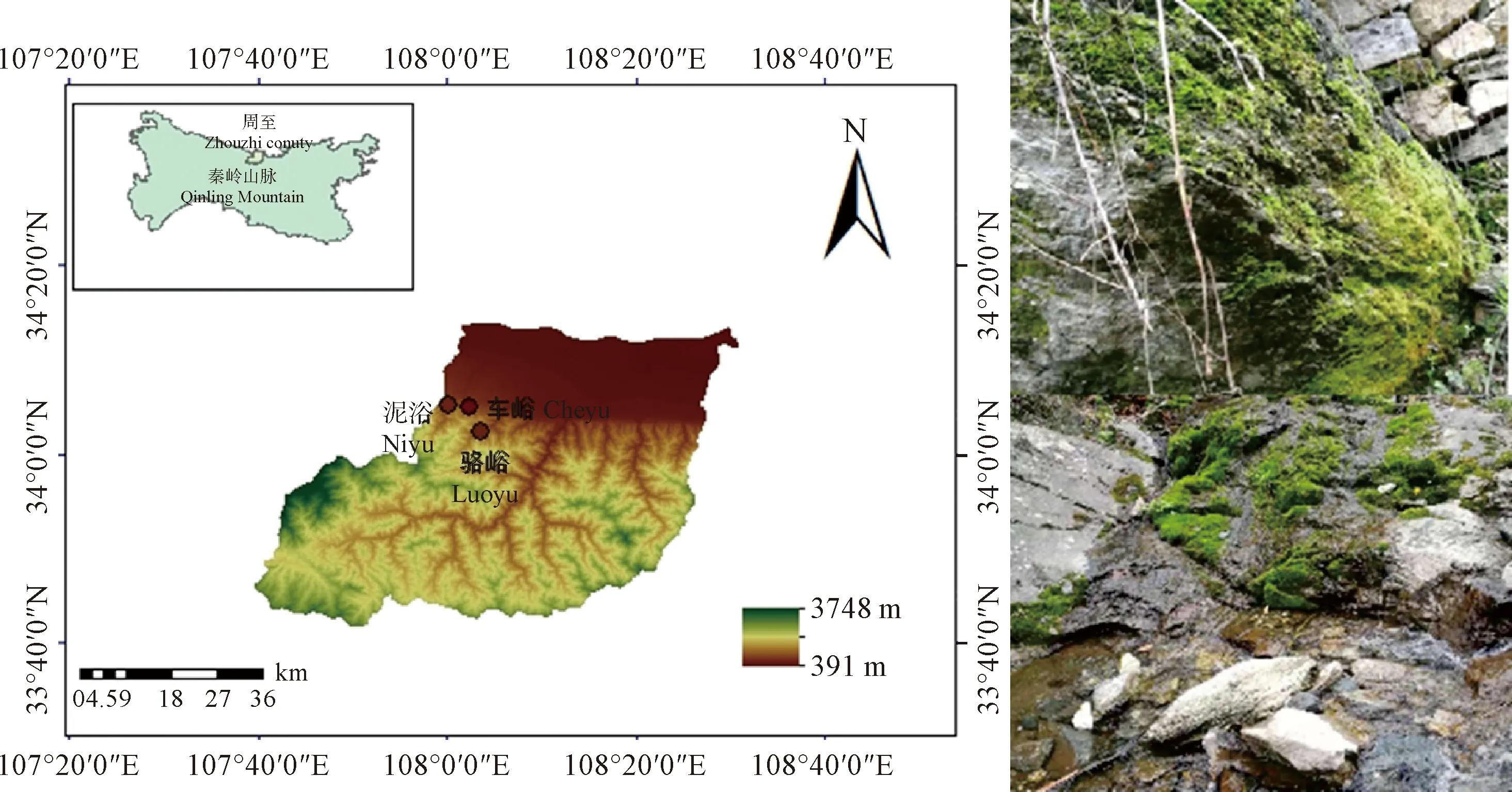
图1 秦岭石生苔藓结皮研究区概况
1.2 样品采集
经过多次的野外调研分析,最终在泥峪、车峪、骆峪选择了5个经、纬度大致相同的、结皮生长良好、无人为破坏的采样点(表1),按5点采样法采集石生苔藓结皮层并混合均匀,每个样点3个重复;并用毛刷刷取基质层(>4 g),装入自封袋带回。一部分于-80 ℃冷冻用于微生物测序,一部分自然风干后过筛用于土壤理化性质测定。

表1 秦岭石生苔藓结皮采样点基本信息
1.3 实验方法
(1) 养分测定:用减量法称取过0.149 mm筛孔的基质土壤各2.0 g,分别用重铬酸钾氧化-外加热法、半微量凯氏法、碱熔-钼锑抗比色法、碱熔-火焰光度法测定有机质(OM)、全氮(TN)、全磷(TP)、全钾(TK)。
(2) 微生物测序:对样本DNA进行提取,利用琼脂糖凝胶电泳检测DNA的纯度和浓度,取适量的样本DNA于离心管中,使用无菌水稀释样本至1 ng/μL。以稀释后的基因组DNA为模板,扩增16S的V4区和ITS1区引物,使用1×TAE浓度2%的琼脂糖胶电泳纯化PCR产物。选择主带大小在 400~450 bp 之间的序列,割胶回收目标条带。使用Illumina公司TruSeq DNA PCR-Free Library Preparation Kit建库试剂盒进行文库的构建,构建好的文库经过 Qubit 定量和文库检测,合格后,使用IonS5TMXL测序平台测序。
1.4 数据处理
使用Cutadapt软件对微生物测序数据进行质控,得到最终有效数据。利用Uparse软件对有效数据在97%的水平上序列聚类成为OTUs,以OTUs中出现频数最高的序列作为OTUs的代表序列。用Mothur算法与SILVA132的SSUrRNA数据库进行分类注释,并在门水平进行相对丰度分析。使用Qiime软件(Version 1.9.1)计算结皮微生物群落的α多样性指数。数据分析均利用Excel 2016和SPSS 2019软件进行,各样点变量之间的显著差异采用单因素方差分析和多重比较分析,土壤因子和微生物群落结构之间的相关性采用Spearman相关性分析,设置显著水平P<0.05。使用R语言软件对土壤因子和微生物群落进行冗余分析(redundancy analysis,简称RDA)。
2 结果与分析
2.1 苔藓结皮层的养分特征
对结皮层养分含量进行单因素方差分析和多重比较分析测定,结果显示(图2):研究区内全氮含量丰富,5个采样点间均差异显著。其中,CY-2的全氮含量最高,为8.26 g/kg;LY-1的全氮含量最低,为4.16 g/kg。研究区内全磷含量最高的样点是CY-1(1.21 g/kg),其次是NY-1和CY-2,且彼此间均无显著差异。全磷含量最低的样点是LY-1,仅为0.88 g/kg,与其他采样点之间差异显著。LY-1的全钾含量显著高于其他样地,达到18.10 g/kg;5个样地土壤有机质含量表现趋势与全氮表现一致,其中含量最高的是CY-2的131.05 g/kg,最低的是LY-1的89.44 g/kg。
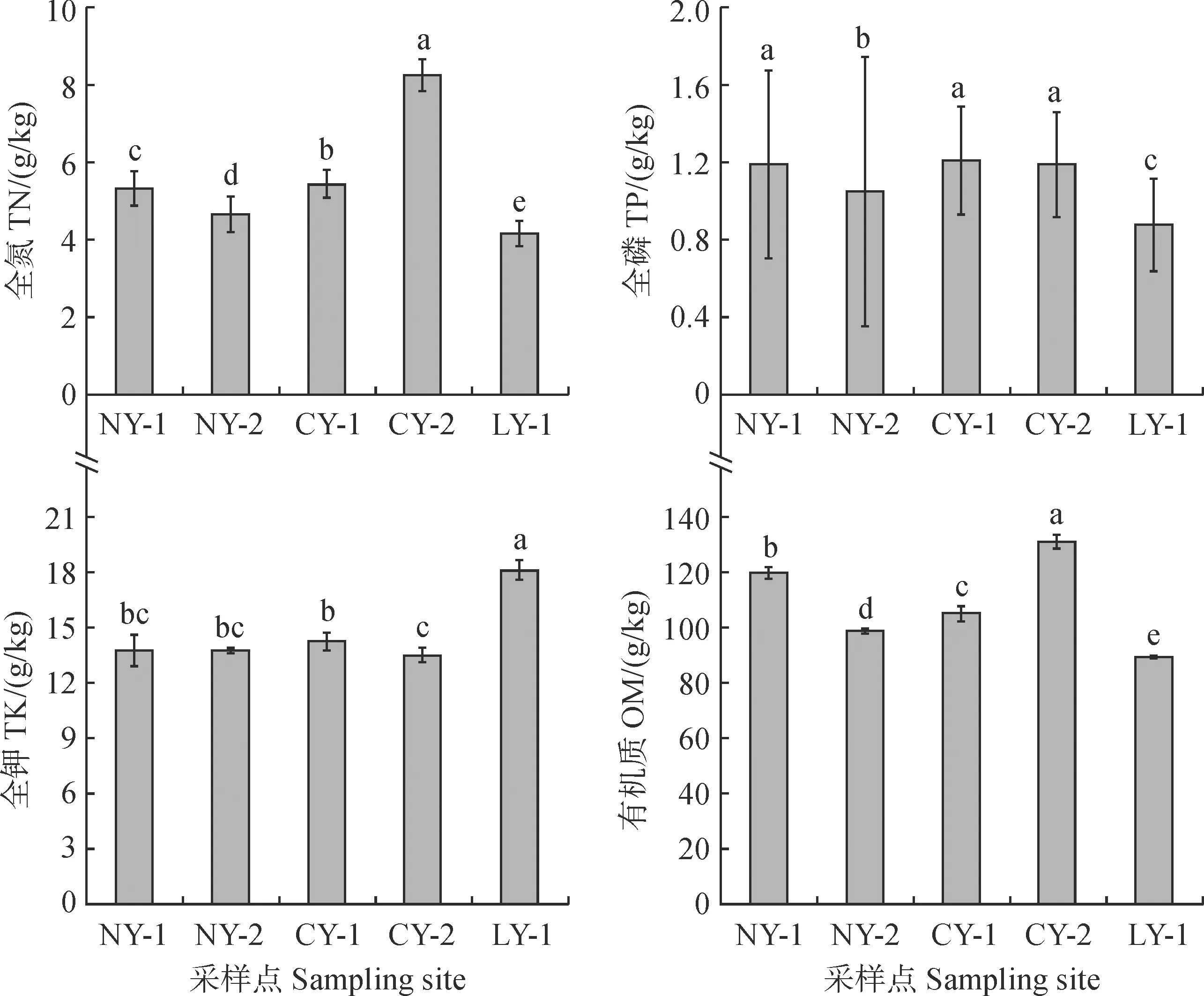
图2 秦岭石生苔藓结皮不同采样点的养分含量比较
2.2 苔藓结皮层的微生物群落组成与多样性
表2和表3反映的是细菌和真菌的α多样性。样本的测序深度指数(Coverage指数)均在0.976~0.999的区间内,说明测序结果能够展示样品中绝大部分信息。对于细菌而言,不同采样点的群落多样性由大到小依次为:CY-1>CY-2>NY-1>LY-1>NY-2;丰富度:CY-1>CY-2>NY-1>LY-1>NY-2。真菌的群落多样性由大到小依次为:CY-1>NY-1>LY-1>NY-2>CY-2;丰富度:CY-1>NY-2>LY-1>NY-1>CY-2。对α多样性指数进行单因素方差分析发现各样地的species、shannon、chao1指数均无显著差异。

表2 秦岭石生苔藓结皮的细菌α多样性指数

表3 秦岭石生苔藓结皮的真菌α多样性指数
5个采样点苔藓结皮层中细菌相对丰度如图3所示。变形菌门(Proteobateria)、拟杆菌门(Bacteroidetes)、放线菌门(Actinobacteria)、酸杆菌门(Acidobacteria)、蓝细菌(Cyanobacteria)、浮霉菌门(Planctomycetes)、绿弯菌门(Chloroflexi)、芽单胞菌门(Gemmatimonadetes)、疣微菌门(Verrucomicrobia)为研究区石生苔藓结皮层细菌群落的主要组成菌群,占细菌群落的95%以上。

图3 秦岭石生苔藓结皮的细菌群落相对丰度
对NY-1采样点来说,变形菌门占主导地位,相对丰度为33.22%,经LSD多重比较结果显示,NY-1采样点的变形菌门与NY-2和LY-1采样点具有显著差异(P<0.05),与其他2个采样点的差异不显著;其次是拟杆菌门和放线菌门,其相对丰度分别为24.97%和12.14%,拟杆菌门的相对含量与其他4个点之间均无显著差异,放线菌门仅和LY-1采样点有显著差异(P<0.05)。NY-2采样点的苔藓结皮层中细菌的相对丰度表现为拟杆菌门>变形菌门>酸杆菌门>变形菌门,其中,放线菌门的相对含量与LY-1有显著差异(P<0.05),变形菌门与NY-1采样点的有显著差异(P<0.05)。NY-2采样点的拟杆菌门和酸杆菌门在含量上与其他采样点之间均无显著差异。车峪2个采样点的细菌含量均表现出相同的趋势,均为变形菌门>拟杆菌门>酸杆菌门>放线菌门,且4种细菌相对含量在两地均无显著差异。不同于其他采用点,LY-1采样点的主要优势菌群为放线菌门,其相对含量高30.34%,显著高于其他4个采样点的放线菌的相对含量。
在属水平上,排名前十的菌属是未知蓝细菌属(unidentified_Cyanobacteria)、深铁杆菌属(Ferruginibacter)、人参地黄色土源菌属(Flavisolibacter)、未知酸杆菌属(unidentified_Acidobacteria)、马赛菌属(Massilia)、苔藓杆菌属(Bryobacter)、土壤红色杆菌属(Solirubrobacter)、克罗氏菌属(Crossiella)、黄杆菌属(Flavobacterium)和金黄杆菌属(Chryseobacterium)。其中,未知蓝细菌属是属水平上丰度最高的菌属,在NY-1和CY-2都有很高的含量。马赛菌属是变形菌门中含量最高菌属,在LY-1大量分布且显著高于其他样点(P<0.05)。酸杆菌门主要由苔藓杆菌属和未知酸杆菌属组成,未知酸杆菌属在5个采样点均有分布,且无显著差异;NY-2样点中的苔藓杆菌属丰度显著高于其他样点(P<0.05)。深铁杆菌属是一种反硝化细菌,能够还原硝态氮,在NY-1和CY-1中大量分布,而在LY-1和CY-2点分布较少。
图4是在门水平上真菌群落物种相对丰度图,排名前十的真菌门是子囊菌门(Ascomycota)、担子菌门(Basidiomycota)、被孢霉门(Mortierellomycota)、毛菌门(Mucoromycota)、壶菌门(Chytridiomycota)、罗兹菌门(Rozellomycota)、单毛壶菌门(Monoblepharomycota)、球囊菌门(Glomeromycota)、油壶菌门(Olpidiomycota)、芽枝菌门(Blastocladiomycota)。子囊菌门在采样点的平均相对丰度最高(46.33%)。CY-2处的子囊菌门含量显著高于其他采样点(P<0.05),其他4个采样点之间的子囊菌门群落相对丰度均无显著差异。NY-2采样点的担子菌门含量最高,且与其他采样点之间无显著差异。被孢霉菌门在5个采样点之间的相对丰度表现为NY-1>LY-1>NY-2>CY-1>CY-2,且两两之间差异不显著。担子菌门在NY-1点相对含量更高,但与其他4个采样点差异不显著。
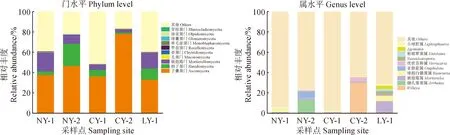
图4 秦岭石生苔藓结皮的真菌群落相对丰度
在属水平上,Willeya、被孢霉属(Mortierella)、健孔菌属(Arrhenia)、球孢白僵菌属(Beauveria)、小球腔属(Leptosphaeria)、亚脐菇属(Omphalina)、疣状苔藓属(Verrucaria)、Neosulcatispora、Agonimia、粉褶覃菌属(Entoloma)是主要的优势菌属。Willeya是子囊菌门中含量最高的菌属,大量分布在CY-2,此外在LY-1和NY2也有极少量分布。疣状苔藓属在5个采样点均有分布,CY-2的含量显著高于其他4点。球孢白僵菌属是子囊菌门中丰度排名第二的菌属,主要分布在LY-1和NY-1中。被孢霉门中主要以被孢霉属为主,也是LY1含量最高的菌属,丰度达11.69%。健孔菌属、亚脐菇属和粉褶覃菌属是担子菌门中占优势的菌属。健孔菌属也是NY-2最主要的菌属,此外NY-2采样点还有相当含量的亚脐菇属存在。
2.3 苔藓结皮层微生物群落与养分的关系
2.3.1 细菌群落与养分的关系将不同采样点的苔藓结皮层门水平上相对丰度占前十的细菌类群与土壤养分因子进行冗余分析,得到RDA图(图5)。第一轴和第二轴对细菌菌落的解释比例分别为85.12%和6.75%,与第一轴关系最密切的是全钾,与第二轴关系最密切的是有机质,两轴累计解释变量达91.87%,说明土壤因子对细菌群落结构具有极大的影响。经蒙特卡洛检验,全钾和有机质对细菌群落结构影响显著。变形菌门的相对丰度与全氮、有机质呈显著正相关(P<0.05)、全磷呈极显著正相关(P<0.01)、全钾呈显著负相关(P<0.05);拟核杆菌与全钾呈显著负相关(P<0.05),与全氮、全磷、有机质呈不显著正相关;放线菌门的丰度与全氮呈显著负相关(P<0.05),与有机质呈极显著负相关(P<0.01);酸杆菌门与全钾呈显著正相关(P<0.05),与其他3种因子呈不显著正相关;绿弯菌门与全氮和有机质呈显著负相关(P<0.05);疣微菌门与全氮和有机质呈显著正相关(P<0.05),与全磷呈极显著正相关(P<0.01);蓝细菌、浮霉菌门、芽单胞菌门与土壤因子均呈不显著相关性。

图5 秦岭石生苔藓结皮细菌的相对丰度与土壤养分的RDA分析
2.3.2 真菌群落与养分的关系通过冗余分析,得到不同采样点的苔藓结皮层门水平上相对丰度占前十的真菌类群与土壤因子的RDA图(图6)。由图可知,第一轴与第二轴分别解释68.26%和30.23%的真菌群落组成。经蒙特卡洛检验,全磷、有机质对结皮层真菌的群落组成有显著影响。
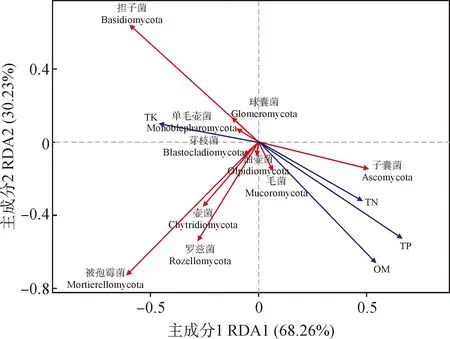
图6 秦岭石生苔藓结皮真菌的相对丰度与土壤养分的RDA分析
子囊菌门的相对丰度与全氮和有机质含量呈显著相关性(P<0.05)、与全钾呈极显著负相关(P<0.01)、担子菌门的丰度与全钾和有机质呈极显著相关性(P<0.01)、与全氮呈显著相关(P<0.05)。其余真菌种类与土壤养分均无显著相关性。
3 讨 论
3.1 秦岭石生苔藓结皮层的养分状况
根据全国第二次土壤养分分级标准,采样点结皮层土壤有机质含量均达到了极丰富级别;全氮含量均显著高于2 g/kg,达到了极丰富级别。研究区内丰富的植被资源和土壤微生物对固定大气中的氮起着关键作用,能够提高土壤中氮含量。除CY-1的全磷含量达到中上级外,其余采样点全磷量都位于0.88~1.19 g/kg范围内,属于中下级别。土壤全磷含量取决于土壤母质、成土过程和施肥作用[30],由于采样点人迹罕至,因此磷的含量较低主要是受到母质与成土作用的影响。此外,有机质含量越丰富,土壤全磷的含量也越高[31],因此土壤有机质较高的车峪-1的磷素含量高于其他4点达到中上级。除骆峪全钾为中上外,其余均为中下。
3.2 秦岭石生苔藓结皮层的微生物特征
对本次调查的5个采样点的结皮层土壤进行高通量分析共得到3界、42门、82纲、212目、403科、771属。其中在细菌群落结构分析中,共有32门、45纲、97目、176科、347属,变形菌门、拟杆菌门、放线菌门、酸杆菌门为优势细菌菌门。腾格里沙漠苔藓结皮层的优势细菌门为变形菌门(29.94%)、蓝细菌门(16.38%)和放线菌门(11.29%)[32],毛乌素沙地苔藓结皮层优势细菌门为放线菌门(38.52%)和变形菌门(26.01%)[33],表明苔藓结皮层微生物优势群落与生境有关。未知蓝细菌属、深铁杆菌属、人参地黄色土源菌和未知酸杆菌属是优势菌属,此外马赛菌属、苔藓杆菌属的含量也相当丰富。土壤细菌主要养分来源为根系分泌物和凋落物,旱区与湿润区的营养差异导致土壤细菌群落组成有所不同[34]。
与旱区结皮微生物的研究结果不同[22,33],研究区内拟杆菌门的丰度高于放线菌和酸杆菌,拟杆菌门是一种富营养型的异养细菌[35],研究区湿润的气候、丰富的枯落物和有机质为其提供了适宜的生长条件。而放线菌和酸杆菌都是贫营养型细菌[35-36],能分解难降解的碳库,能更好地适应干旱贫瘠的生态系统。因此在多数研究中都是旱区丰度最高的优势菌门。蓝细菌是生物结皮早期演替的主要固氮微生物,Steven等[37]的研究表明,气温升高和降水量的增加会显著降低蓝细菌门的相对丰度,因此相较于腾格里沙漠和毛乌素沙地,研究区内蓝细菌的含量较低。
土壤真菌是土壤微生物的重要组成部分,其物种多样性在维持生态系统的平衡中起着关键作用[38]。对于真菌群落结构分析中,共有10门、37纲、105目、227科、424属,子囊菌门和被孢霉门为优势菌门。子囊菌门在研究区是绝对的优势真菌,这与旱区的研究一致,尽管他的相对丰度远小于在旱区的研究中的含量,仅为46%,这或许是因为其他真菌如被孢菌门等的含量增加。事实上子囊菌群在全球土壤真菌中占主导地位[39],其群落主要由腐生菌构成,可以通过分泌多种酶降解复杂有机质,为苔藓提供养分[40-41]。在属水平上,Willeya、被孢霉属、健孔菌属、球孢白僵菌、小球腔属、亚脐菇属、疣状苔藓属、Neosulcatispora、Agonimia、粉褶覃菌属是主要的优势菌属。Willeya是子囊菌门下的优势属,与疣状苔藓属同属于Verrucariaceae科,广泛存在于岩石上,可以通过分泌酸来帮助分解石灰岩。疣状苔藓属和亚脐菇属都是担子菌门下的优势属,通常与苔藓呈现出某种互惠关系。
3.3 土壤养分和微生物群落组成的关系
微生物是生物结皮形成的参与者,又受到生物结皮发育程度的影响[42]。微生物群落结构特征是土壤整体状况的综合反映,土壤养分对土壤微生物群落结构特征具有重要影响[43]。
生物结皮能够改善土壤理化性质[44],从而显著影响土壤微生物的群落结构[33, 45]。冗余分析显示全钾及有机质是影响结皮下土壤细菌群落组成变化的主要动力,其中全钾是最主要的影响因子。
研究区的全氮和有机质均保持在较高水平,这或许是因为变形菌的存在。变形菌是氮循坏中发挥主要作用的物种[46],也是参与有机质分解转化的主要功能细菌[47-48],能够在植物根际快速地富集,在湿度大[49]且营养丰富的土壤中具有更高的存在比例[38, 50]。马赛菌属是变形菌门下含量最高的菌属,属于β——Proteobateria,对营养状态的改变有很高的敏感性,有研究发现马赛菌属的相对丰度与土壤活性有机碳含量呈正相关关系[51],这与本研究结果一致。同时,马赛菌属具有溶磷的作用,在马赛菌属分布较多的地方,磷的含量也较丰富。人参地黄色土源菌属、深铁杆菌属、黄杆菌属都是拟杆菌门下的菌属,作为根际促生菌,能够促进石生苔藓的生长。放线菌丰度与全氮含量呈负相关,这与樊瑾等[52]在毛乌素的研究一致,但周虹等[53]的研究表明,放线菌与全氮呈正相关。这是由于放线菌发育缓慢,当环境提供的营养物质较多时,竞争能力弱;而当营养物质较多时,放线菌数量反而占优势[54]。
研究区苔藓结皮层中的真菌以子囊菌门和被孢霉门为主,子囊菌门含量与全氮、有机质含量呈现出显著正相关,子囊菌门以腐生菌为主,土壤中难以分解的木质素、纤维素等复杂的有机质都可以被它分解成植物可直接利用的有机质。而丰富的有机质可以增加子囊菌门的丰度[55]。不同于赵宇龙等[50]在毛乌素沙地的研究,被孢霉门的丰度占到10%左右,是该区的优势真菌菌门之一。被孢霉菌是一类有益的腐生真菌,可补充氮素、有机碳,在碳氮丰富的土壤中,其丰度显著增加[56]。被孢霉能够分解难利用的磷,在低磷水平的土壤中有较好的适应性,这或许是采样点结皮微生物中被孢霉含量高的原因之一。
4 结 论
本研究发现秦岭石生苔藓结皮层的优势细菌主要是变形菌、拟核杆菌、放线菌和酸杆菌,其中放线菌和酸杆菌的含量相对于旱区而言有所下降,而拟杆菌含量显著增加;子囊菌门、被孢霉菌门和担子菌门的丰度占了真菌的70%左右,但与旱区相比,子囊菌门和担子菌门的相对丰度明显减少,而被孢菌门丰度显著增加,甚至成为了优势菌门。有机质、全氮、全磷和全钾对群落组成都有一定的影响,其中有机质是影响结皮层微生物群落组成最主要的养分因子。研究结果为阐明苔藓结皮在湿润地区的发育形成机制及功能发挥机理奠定基础,可为裸露岩石创面的生态修复提供借鉴。
猜你喜欢
科学技术与工程(2022年26期)2022-11-01
现代园艺(2022年19期)2022-09-21
中国农学通报(2022年14期)2022-06-01
野生动物学报(2022年2期)2022-05-16
小读者(2021年19期)2021-11-07
湖北农业科学(2019年22期)2019-12-23
作文评点报·中考版(2019年4期)2019-02-18
课外语文·中(2018年10期)2018-11-16
农业与技术(2017年12期)2017-07-11
江苏农业科学(2016年6期)2016-07-25
