
基于部分NCII交配设计的陆地棉产量和纤维品质性状遗传分析
2022-11-18 06:46冯常辉焦春海张友昌秦鸿德王琼珊张教海王孝刚夏松波蓝家样陈全求
作物杂志 2022年5期
冯常辉 焦春海 张友昌 别 墅 秦鸿德 王琼珊张教海 王孝刚 夏松波 蓝家样 陈全求
(1湖北省农业科学院经济作物研究所/农业农村部长江中游棉花生物学与遗传育种重点实验室,430064,湖北武汉;2湖北省农业科学院,430064,湖北武汉)
我国现有各类棉花种质资源有8000余份[1],为育种提供了丰富的遗传资源。了解和掌握陆地棉资源材料育种目标性状的遗传特点,对育种实践具有重要的指导意义。皮棉产量和纤维品质是棉花育种的主要目标性状,其遗传特点一直是育种家关注的重点。对于衣分和纤维品质性状的遗传效应,已有研究结果较为一致,多数认为以加性效应为主[2-5]。但其他产量性状的遗传特点,目前仍缺少统一的认识。对于皮棉产量,不少研究[6-9]认为其遗传效应以显性效应为主,也有研究[3,10]认为以加性效应为主,还有研究[4-5,11-12]认为兼有加性和显性效应。对于铃数和铃重,多数研究[2,6,8]表明,二者均以显性效应为主,但王学德等[10]认为,铃数和铃重以加性效应为主。至于皮棉产量与纤维品质性状的相关性,一致认为二者之间存在负相关性(主要是皮棉产量与纤维上半部平均长度或者断裂比强度之间),但是相关程度差异较大[5,13-15]。有研究[14]认为,皮棉产量与纤维品质之间相关程度较低,二者之间的关系可以协调;也有研究[13]认为,二者之间负相关程度较高,协调二者的关系较为困难。以上结果相互矛盾的原因,一般认为是由于不同研究中所选择的材料不同所致。
在已有的陆地棉产量与品质性状的遗传研究中,所用交配设计主要为双列杂交或双因素交叉式遗传(North Carolina II,NCII)。当使用的亲本数量增加时,这2种交配设计所需杂交组合数量接近指数增长。因试验田面积和性状调查所耗时间等科研条件的限制,多选用不超过12个亲本的交配群体进行遗传分析[16],因此材料选择不同而造成的结果差异难以避免。为在资源有限的条件下评价更多的亲本,Kempthorne等[17]和 Miranda等[16]分别提出了部分循环双列杂交和群体间的部分循环双列杂交。对于群体间的部分循环双列杂交,因其属于NCII设计的特殊类型,因此也被称作部分NCII交配设计[18]。从交配方式上看,部分NCII交配设计也属于不完全双列杂交的一种,可用混合线性模型分析方法进行遗传效应分析[19-21]。
为避免亲本数量少而造成遗传分析结果对材料选择的严重依赖,本研究以60份不同地域来源的陆地棉材料为亲本,采用部分NCII交配设计配制杂交组合,利用加性-显性(AD)遗传模型剖析陆地棉产量和品质性状的遗传效应,并分析各性状之间的遗传相关性,进一步了解陆地棉主要的产量和纤维品质育种目标性状的遗传特点;同时对60个陆地棉亲本材料进行评价,为后续育种程序的制定提供理论依据。
1 材料和方法
1.1 试验材料
选用60个陆地棉亲本材料(父、母本各30份)为试验对象(表1)。父本编号范围为1~30,母本编号范围为31~60。所选亲本来源于不同地区,育成时间有较大差别,在熟性、产量和纤维品质等性状上具有不同特点,其中有11个为生产推广应用优良杂交品种的亲本。

表1 亲本名称及其来源Table 1 The name and origin of parents
1.2 交配设计
采用部分NCII交配设计[18],按每个父本分别与6个母本杂交,每个母本分别与6个父本杂交的循环方式产生180个F1组合(表2)。
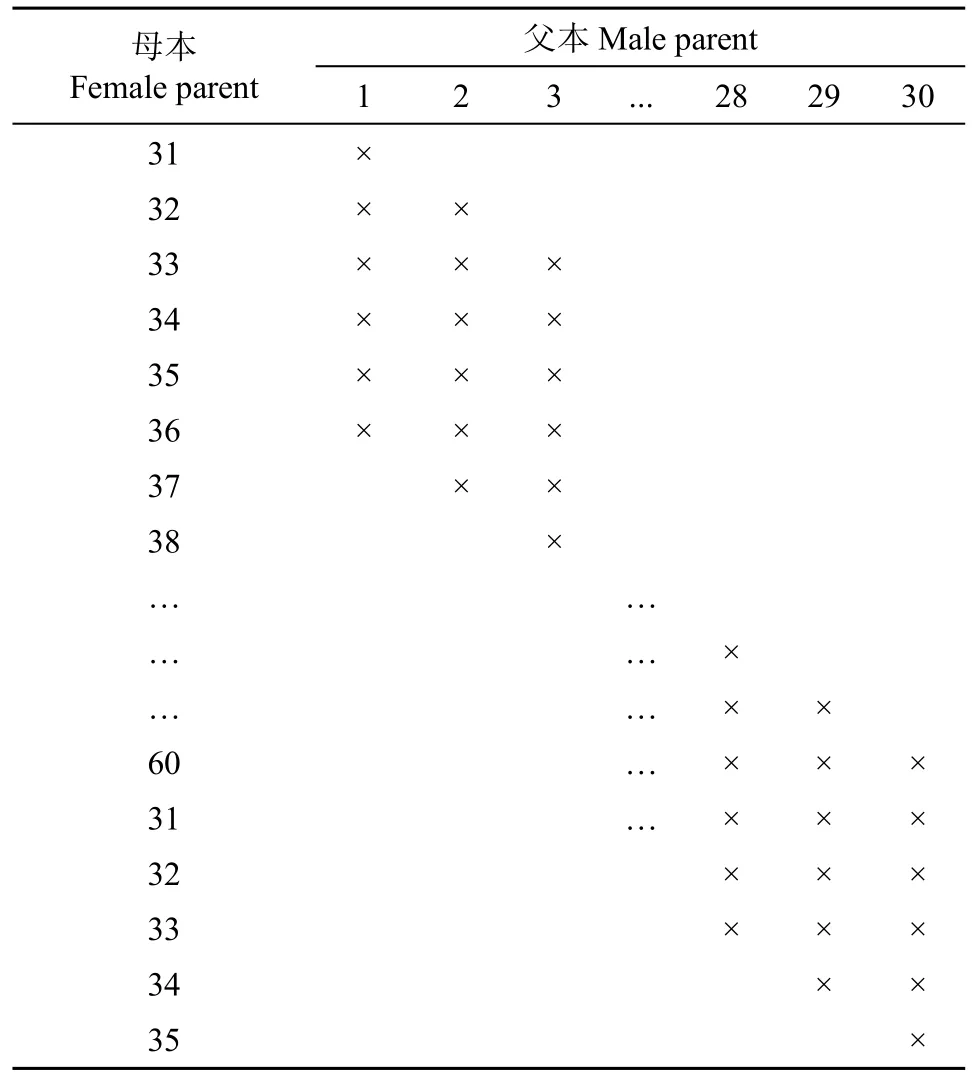
表2 部分NCII交配设计的交配方式Table 2 Mating patterns in partial NCII mating design
1.3 田间试验
2017年在海南配制杂交组合。60个亲本和180个F1杂交组合,将240个材料种植于3个环境:2018年种植于湖北省农业科学院经济作物研究所武汉试验基地,2019年种植于湖北省农业科学院经济作物研究所武汉试验基地和鄂州试验基地。3个环境的田间试验均采用随机区组设计,单行区,行长4.40m,行距0.76m,3次重复。其中,武汉试验点在2018和2019年均于4月17日左右进行营养钵育苗,5月11日前后移栽,种植密度3.30万株/hm2;鄂州试验点于4月25日直播,种植密度3.90万株/hm2。栽培管理同一般大田。
1.4 性状调查
在9月20日前后,调查每小区总铃数。每行采收棉株中部正常吐絮的棉铃50个,考察铃重(50个铃重的平均值)、衣分(%,50个棉铃去掉棉籽后的皮棉/50个棉铃重量×100)以及纤维品质。小区皮棉产量按“(铃重×小区总铃数×衣分)/小区面积”计算。采用HFT 9000大容量纤维检测仪测定纤维上半部平均长度、断裂比强度和马克隆值。
1.5 数据处理
运用AD遗传模型对240个试验材料在3个环境下的表型数据进行联合分析,参照文献[21-24],利用QGAStation 2.0软件进行遗传方差分量估算、遗传效应及基因型相关性分析。运用MINQUE(1)法估算各性状各项遗传效应的方差分量,运用Jackknife抽样方法计算各遗传参数的估计值及其标准误,采用t检验法对遗传参数做统计检验。采用SPSS 17.0完成描述性统计分析、方差分析和简单相关系数计算。
2 结果与分析
2.1 亲本和杂交组合的产量与纤维品质表现
对180个F1和60个亲本的皮棉产量及其构成性状、纤维品质表现进行描述性统计分析(表3)。7个性状的所有偏度绝对值介于0和1之间,接近正态分布;从峰度上看,皮棉产量、铃重和铃数的分布峰较陡峭,数据值较为集中。皮棉产量(164.21~3051.49kg/hm2)、铃数(18.66×104~118.78×104/hm2)、衣分(24.48%~48.72%)、铃重(2.94~7.58g)、上半部平均长度(24.00~33.60mm)、断裂比强度(23.10~38.70cN/tex)和马克隆值(3.50~6.60)等性状的变化范围较大。

表3 部分NCII群体的产量和纤维品质性状描述性统计分析Table 3 Descriptive statistic characteristics of the yield and fiber quality traits in the partial NCII mating design population
从方差分析结果(表4)可知,基因型、环境以及基因型×环境互作均对性状产生显著或极显著影响。
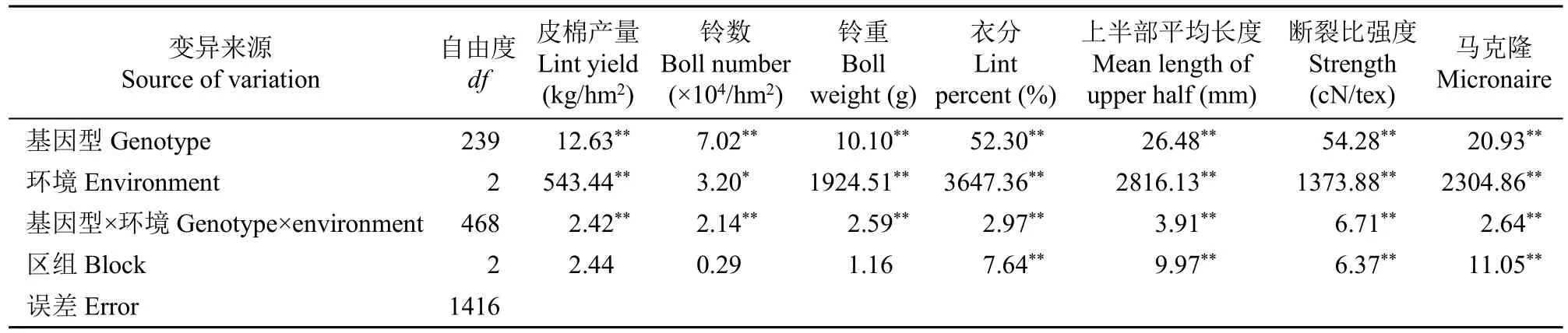
表4 部分NCII群体中目标性状方差分析Table 4 Variance analysis on the breeding target traits in the partial NCII mating design population
2.2 产量和纤维品质目标性状的遗传方差构成
对180个F1和60个亲本的皮棉产量及其构成因素和纤维品质性状的方差分量进行估计(表5)。7个性状中,所有加性方差比例均达极显著水平;除断裂比强度外,其余性状的显性方差比率均达显著或极显著水平。说明既可在常规育种上通过世代选择,使加性效应稳定遗传,也可以在杂种优势利用中发挥部分性状的显性效应。

表5 目标性状的方差分量比例Table 5 Estimated proportions of variance components for the breeding target traits
皮棉产量的4种遗传方差分量中,加性方差分量(VA/VP)占比最大(0.25),其次为显性方差分量(VD/VP,0.17)和显性×环境互作方差分量(VDE/VP,0.15),加性×环境互作方差分量比例(VAE/VP,0.05)最小。铃数的遗传方差分量中,VA/VP(0.20)也占主要地位,约为VD/VP(0.07)的3倍,但VDE/VP(0.14)却显著高于VAE/VP(0.08),约为后者的1.7倍。铃重的遗传方差分量中,VD/VP(0.21)占主要比例,其次为VDE/VP(0.17)和 VA/VP(0.14),VAE/VP(0.05)所占比例较小。衣分的VA/VP(0.62)在所有方差中最大,约为VD/VP(0.13)的4.8倍。
对于3个纤维品质性状,纤维上半部平均长度、断裂比强度和马克隆值的VA/VP在4类遗传方差中均为最大,分别为0.48、0.60和0.48,而VD/VP均接近零,VDE/VP均较小(0.08~0.22)。
所有育种目标性状的VAE/VP在总遗传方差中均占比最低(0.03~0.08),而VDE/VP均不低于VAE/VP,表现为断裂比强度(0.22)>上半部平均长度(0.18)>铃重(0.17)>皮棉产量(0.15)>铃数(0.14)>马克隆值(0.08)>衣分(0.07)。
2.3 亲本的加性遗传效应
对60个亲本的产量及其构成因素的加性遗传效应进行估计(表6)。皮棉产量性状上,亲本的加性效应负向最小(-37.19),正向最大(23.65)。在加性效应值排序前10的亲本中,58(JA501)和56(荆3372)的衣分和铃重加性效应较大,60(鲁棉398)和8(宜棉3号)的铃数和铃重加性效应较强,38(D5M)的衣分和铃数加性效应较强,而亲本29(华惠15)的铃数加性效应较强(25.28)。亲本33(苏棉12号)的衣分加性效应突出(2.58)。

表6 亲本产量及构成性状的加性效应估算值Table 6 Estimated additive effects of yield and its components for parents
皮棉产量加性效应较高的亲本多为优良杂交品种,如 30(泗抗 1 号)、33(苏棉 12 号)、38(D5M)和56(荆3372)等亲本加性效应居于前10位,4(D5F)居11位,这5个亲本同时也是新创12号、苏杂26、南抗3号、鄂杂棉10号和鄂杂棉17号等优良杂交品种的亲本之一[25-28],亲本4和38是鄂杂棉10号的2个亲本,亲本33是苏杂26和南抗3号的共同亲本。皮棉产量加性效应较低的亲本中,多为来自于生态环境差异较大的品种,如4个国外种质资源(9/美G-82、21/美棉2、27/SD1和49/SD2)、3个新疆来源的亲本(13/新陆早15、24/新陆中10号和34/JA532)以及适于新疆种植的品种(19/中36),其皮棉产量加性效应均在-13.0以下。
为选育出强优势组合,利用皮棉产量加性效应较高的材料作亲本是重要途径之一。皮棉产量的加性效应是其相关性状加性效应的综合反映,发挥各皮棉产量构成性状的正向加性效应也可间接提高杂交组合的产量。
纤维上半部平均长度、断裂比强度和马克隆值的加性效应估计值正向最大分别达1.42、3.93和0.36;负向最大分别达-1.19、-2.14和-0.47(表7)。其中,亲本6(鄂棉9号)、9(美G-82)、14(U37-8)、34(JA532)、36(SJB006-2)、46(S6043)、49(SD2)和52(MBC31776)等8个亲本的纤维上半部平均长度和断裂比强度加性效应分别达到0.46和0.13以上,且马克隆值具有负向加性效应。马克隆值是纤维细度和成熟度的综合反映,一般认为马克隆值在3.70以上时,数值越低纤维品质越好。本研究中亲本和组合的马克隆值在3.50~6.60。因此,优质育种中可以利用这些亲本提高纤维上半部平均长度和断裂比强度,并适度利用马克隆值的负向加性效应将其控制在优质范围内。
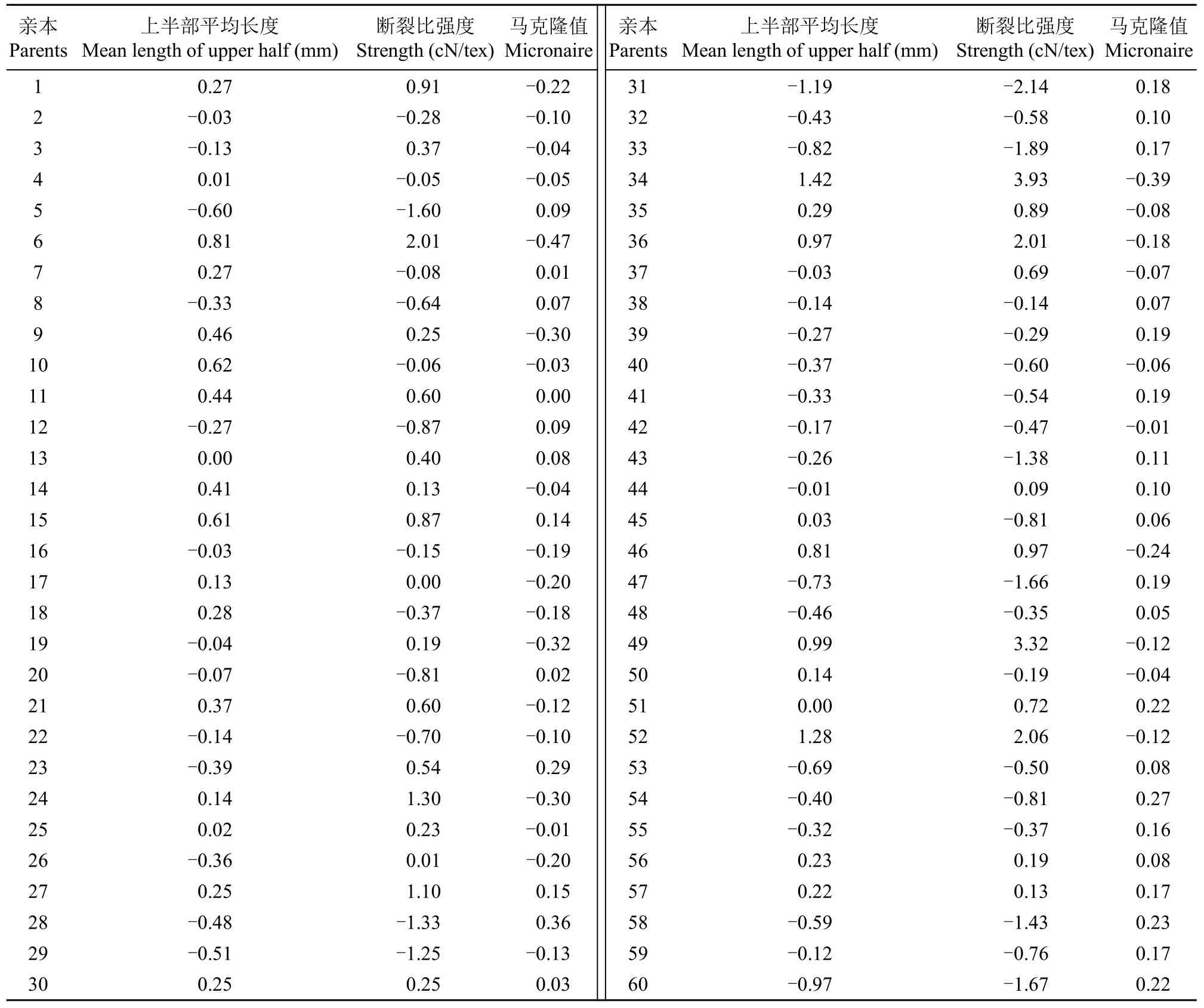
表7 亲本纤维品质性状的加性效应估计值Table 7 Estimated additive effects of fiber quality traits for parents
2.4 性状之间的基因型相关性
性状之间的基因型相关剔除了环境的影响,代表相关性中可遗传的部分[5]。因此,本研究采用基因型相关系数比较成对性状间的相关性。由表8可知,皮棉产量与铃数、衣分和铃重之间呈较强的正相关,相关系数分别为0.83、0.73和0.49。衣分与铃数存在较高相关性,相关系数达0.45,而衣分与铃重、铃重与铃数之间相关性较弱,分别为0.22和0.10。
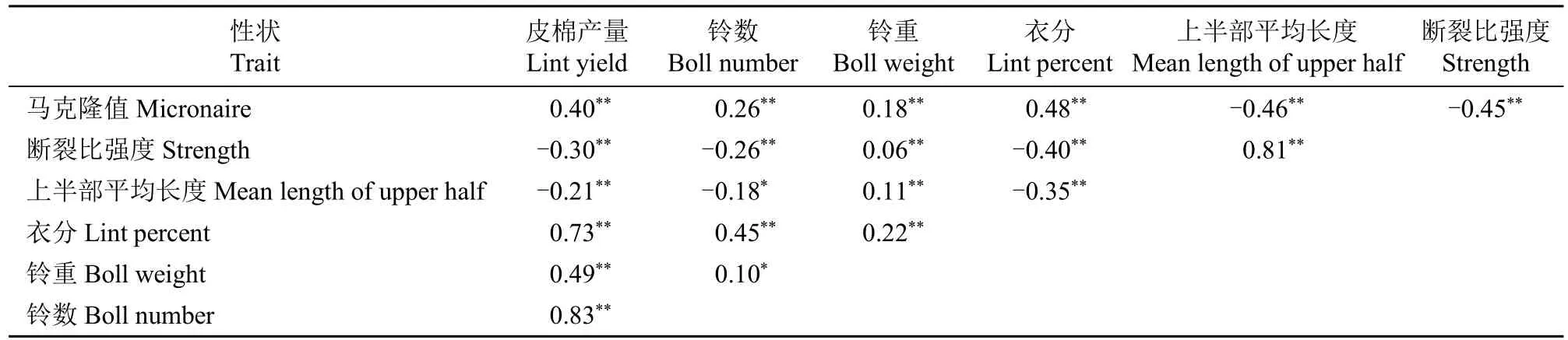
表8 目标性状间的基因型相关系数(r)估计Table 8 Estimated genotypic correlation coefficient(r)among the breeding target traits
纤维上半部平均长度与断裂比强度之间呈极显著正相关(0.81),但它们与马克隆值之间均呈极显著负相关(-0.46和-0.45)。纤维上半部平均长度与断裂比强度同马克隆值之间的负相关实际上有利于纤维上半部平均长度、断裂比强度和马克隆值的同步改良。
产量构成因素与纤维品质性状间,衣分性状与纤维上半部平均长度和断裂比强度间的负相关程度最高,铃数与上半部平均长度和断裂比强度之间的负相关次之,铃重与纤维上半部平均长度和断裂比强度则有一定程度的正相关。但衣分、铃数和铃重均与马克隆值呈正相关,且以衣分的相关性最强。皮棉产量与纤维上半部平均长度和断裂比强度存在较小负相关性(-0.21和-0.30),而与马克隆值呈中等程度正相关(0.40)。
3 讨论
3.1 部分NCII交配设计
部分NCII交配方式属于NCII交配设计的一种抽样[17],在一定程度上可以代表NCII交配设计,其主要优点为能够对较大容量的亲本进行遗传分析和评价,但其结果与NCII的一致性如何取决于抽样的代表性[29-30]。为平衡部分NCII设计容纳样本的数量和抽样的代表性,本研究将亲本以接近随机的方式进行位置排序,而且以代表性较好的平衡交配方式配制杂交组合[30],使每个亲本能与不同来源或类型的亲本进行杂交。本研究利用部分NCII群体和AD遗传模型对遗传方差比例和亲本遗传效应进行分析,在衣分和纤维品质性状上得到的结果与此前的相关研究呈现较高的一致性。目前已有研究[2-5]结果认为,衣分和纤维品质性状的遗传以加性效应为主,本研究的结果也显示,衣分和纤维品质性状的加性效应方差比例为0.48~0.62,在遗传方差中占绝对优势。在铃重性状上,本研究与较多的研究[7,9,31-32]一致认为其主要受显性遗传效应控制。
本研究使用的亲本数量较以往相关研究有较大幅度的增加,在产量性状的遗传特点上也得到了与以往研究存在一定差异的结果。已有的陆地棉产量性状遗传分析结果[2,4,12,31]认为,产量性状除衣分外,均以非加性效应为主,加性效应的方差比例低于显性效应或环境互作效应。而本研究结果显示,皮棉产量和铃数的加性效应在各类遗传效应中占比最高,显性及其环境互作效应的比例低于加性效应。出现这种差异的原因除与不同研究材料的选择有关外,还可能与样本容量有关。因为样本容量对遗传参数估计有一定的影响,交配亲本或组合较多时估计值更可靠[33]。在已有相关研究[2,5-9]中,使用的亲本数量多在12个以下。但在王学德等[10]和Jenkins等[3]的研究中,其亲本数量分别为15和20个,结果也显示皮棉产量的加性效应比例超过显性效应。
3.2 环境对陆地棉产量和品质性状的影响
棉花植株个体较大,具有无限生长习性,产量和品质性状,尤其是除衣分外的产量性状易受环境影响。设置重复和增加环境因素可有效降低环境对分析结果的影响。在本研究中,3个环境因素对产量和纤维品质性状产生显著或极显著影响。Wu等[32]通过分析12个亲本及其37个杂交组合的遗传方差,表明皮棉产量、衣分和铃重的显性与环境互作效应显著,高于加性与环境互作效应。Campbell等[34]在育种群体中估算遗传方差,表明皮棉产量、衣分和纤维品质性状的显性与环境互作效应显著,高于加性与环境互作效应。本研究表明,3个纤维品质性状与产量性状一样,具有较高的环境互作方差,其加性×环境和显性×环境方差分量之和分别达到0.23、0.25和0.16(表5),产量性状和纤维品质育种7个育种目标性状的遗传与环境互作效应主要通过显性×环境体现。
3.3 陆地棉产量和纤维品质性状遗传特性与育种策略
育种目标性状的遗传特性决定了育种策略的选择。已有研究[32]认为,皮棉产量和铃数的加性效应比例偏低,因此,在利用杂交育种方式进行常规品种选育时,皮棉产量和铃数等性状选择一般在高世代进行。而本研究结果显示,尽管皮棉产量和铃数的误差方差比例较高,但其加性效应比例在4类遗传效应中仍然最高。这表明在不同亲本材料组成的群体中,皮棉产量和铃数可能存在不同的遗传特点。利用以加性效应为主的亲本群体进行杂交育种时,在后代选择过程中,在控制好试验误差的情况下,对皮棉产量和铃数性状在低世代选择也可以取得较好的收益。
常规棉品种选育过程中,皮棉产量与纤维品质存在较高程度的负相关,同步改良难度较大。本研究结果显示,纤维上半部平均长度和断裂比强度性状与皮棉产量的负相关程度(-0.21和-0.30)低于杨六六等[5]和Campbell等[13]的研究结果,表明利用不同来源的亲本材料进行杂交,降低负相关程度具一定的可能性。最近研究[35]也显示,控制衣分与纤维强度的染色体片段之间存在紧密连锁的证据,但同时发现大量控制纤维品质性状与控制产量性状的染色体片段之间存在不连锁的现象,为打破产量与纤维品质的负相关提供了依据。
本研究中多数目标性状的遗传效应以加性为主的结果也提示,在杂交棉育种中,重视亲本选配的同时需要重视亲本的性状改良,尤其是打破产量性状与品质性状间的不利相关,选育产量性状和品质性状协调的亲本,使不同亲本的组合聚集更多有利基因,从而培育优良杂交种。
3.4 亲本材料的遗传评价
了解亲本的遗传效应,对于育种实践中基础材料的选择利用具有重要的意义。因为在育种实践中,常有亲本本身产量表现较好而加性效应不高的现象,因此难以利用它们配制出强优势杂交组合;而有些亲本本身表现一般,但加性效应较大,则可作为常规育种和杂种优势育种中的优良亲本。本研究筛选出的一批具有较高产量加性效应的亲本,同时也是优良杂交品种的亲本[25-28],表明部分NCII交配对亲本的评价具有较好的效果。
此外,鉴定出亲本8(宜棉3号)等多个亲本在产量性状上具有较强的正向加性效应,亲本6(鄂棉9号)等多个亲本具有较高纤维上半部平均长度和断裂比强度的加性效应。为进一步的常规品种选育和优势杂交组合的测配提供了较好的基础。
4 结论
部分NCII交配设计可用于对育种目标性状的遗传特点进行分析,并可同时对多个亲本进行有效评价。通过对陆地棉产量和纤维品质育种目标性状加性效应、显性效应以及其与环境的互作效应进行剖析,表明陆地棉的皮棉产量、衣分和铃数性状的遗传呈现出以加性效应为主的特点,而铃重具有较大的显性效应;纤维上半部平均长度、断裂比强度和马克隆值3个品质性状的遗传以加性效应为主。基因型与环境相互作用主要通过显性效应与环境的互作来体现。相关性分析结果表明,产量性状中,衣分与纤维上半部平均长度、断裂比强度和马克隆值的相关性均最大。
猜你喜欢
中国糖料(2021年3期)2021-07-13
棉花科学(2020年2期)2020-06-01
中国纤检(2019年5期)2019-12-09
中国棉花加工(2018年5期)2019-01-18
热带农业科技(2019年1期)2019-01-14
中国果业信息(2019年11期)2019-01-05
中国棉花(2019年4期)2019-01-04
中国棉花(2018年7期)2018-08-29
安徽农学通报(2016年7期)2016-05-03
中国棉花加工(2015年6期)2015-10-28
