
稳定同位素标记结合超高效液相色谱-质谱技术分析镉胁迫下水稻中的羧基化合物
2022-11-24 07:57曾宸蔡文静叶甜甜冯钰锜
分析科学学报 2022年5期
曾宸, 蔡文静, 叶甜甜*, 冯钰锜
(武汉大学化学与分子科学学院,湖北武汉 430072)
随着工业化进程的加快,重金属污染问题日益严重。镉是目前毒性最大、分布最广的重金属污染物之一[1]。镉是植物生长非必需金属元素,它通过阻碍植物水分和养分的吸收引起一系列代谢紊乱,如光合强度下降和酶活性的改变,蛋白质、糖和叶绿体合成受阻,使植物叶片萎黄,植株矮化,最终导致作物品质下降和产量减少,甚至死亡[2 - 4]。此外,重金属镉易被植物吸收并在可食用部分大量富集,从而进入食物链,严重影响人体的健康[3,5]。羧基化合物作为生物体内重要的小分子化合物,不仅以底物或者产物的形式直接参与中心碳代谢,而且也是脂类、蛋白质代谢路径中重要的中间代谢物。此外,羧基化合物与植物的生长发育和胁迫应答等生理过程密切相关[ 6- 8]。当植物受到一定量的镉胁迫后,会引起一系列羧基代谢物的含量变化来应对镉胁迫[3,9,10]。例如,在镉胁迫下,水稻会首先通过提高三羧酸循环中的部分前体和相关代谢产物的含量来抵御毒害[3,11]。另外,Zhang等[12]报道了镉胁迫会促进有机酸的合成,而有机酸可与Cd2+发生螯合作用,从而抵御Cd2+的毒害。由此可见,羧基化合物在植物响应镉胁迫中发挥了至关重要的作用[13 - 15]。因此,建立高灵敏度、高覆盖度、高准确性的羧基化合物分析方法,将有助于了解植物对镉胁迫的响应机理,从而减轻镉胁迫造成的危害。
目前,在植物中检测到的羧基化合物数量十分有限。Navarro-Reig等[15]用高效液相色谱-质谱检测植物体内的化合物,发现在镉胁迫条件下,水稻中的74个代谢物含量发生显著变化,其中包括氨基酸、有机酸等22个羧基化合物。Schäfer等[14]建立了固相萃取结合超高效液相色谱-串联质谱法,实现了对100多种初级代谢物、次级代谢物,以及植物激素的高通量定量分析,其中仅包含60多种羧基化合物[14]。然而,这些方法都是在负离子模式下对羧基代谢物进行检测,检测灵敏度通常较低。因此,高灵敏度方法的开发将有助于植物中低丰度羧基化合物的发现和鉴定,从而丰富植物中羧基化合物的种类。
为了全面研究水稻中羧基化合物的组成,本文建立了基于稳定同位素标记技术的羧基化合物的超高效液相色谱-质谱(UPLC-MS)全分析方法,并对镉胁迫下水稻根部和地上部分的含羧基化合物进行了定性和相对定量研究。利用该方法在水稻样品中一共发现了1 444个可能的含羧基化合物,其中648个化合物可以被定性(包括72个标准品确认,13个多级质谱解析和563个数据库检索获得)。同时,相对定量结果表明,镉胁迫条件下,水稻根部中的大部分羧基代谢物显著降低,而地上部分中的95%以上的羧基代谢物含量都显著升高,这表明水稻根部和地上部分可能在响应镉胁迫时,在羧酸代谢水平采用了不同的应对策略。最后,代谢网络分析表明,这些差异代谢物主要富集在苯甲酸族化合物代谢、苯丙氨酸代谢、苯基丙酸生物合成、脂肪酸生物合成、脂肪酸降解、不饱和脂肪酸生物合成、亚麻酸代谢和花生四烯酸代谢等8条路径中。
1 实验部分
1.1 实验试剂
所有羧酸标准品购买自美国Sigma-Aldrich公司、北京百灵威公司、美国Cayman Chemical公司和上海阿拉丁公司。分析纯的甲酸(88%)、2-氯-1-甲基碘化物(CMPI)、三乙胺(TEA)和N,N-二甲基乙二胺(DMED)购买自上海国药集团;同位素标记试剂DMED-d4依照实验室报道过的方法合成[16];色谱纯乙腈购于德国默克公司。TEA(10 μmol/mL)、CMPI(20 μmol/mL)、DMED(20 μmol/mL)、DMED-d4(20 μmol/mL)和有机酸(1 mg/mL)的储备溶液均以色谱纯ACN配制,-20 ℃冰箱中储存。强阴离子交换-固相萃取柱(SAX-SPE,3 mL,200 mg)购自维泰克科技(武汉)有限公司;实验中所用水均为Milli-Q超纯水(美国Millipore公司纯水仪制备)。
1.2 植物样品的收集
籼稻品种(Oryza sativa ssp.indica cv.Zhenshan 97B)由中国水稻研究所-水稻生物学国家重点实验室提供。种子在水培条件下萌发,然后移栽于Hoagland营养液中,于相对湿度70%~80%,光周期为16 h光照(28 ℃)、8 h黑暗(25 ℃)生长10天后,将一半植株用100 μmol/L Cd(NO3)2对大小均匀的幼苗进行镉胁迫。正常组和胁迫组各做4组生物学平行和3组技术平行,胁迫7 d后分别收获茎和根,称重,液氮冷冻后,立即于-80 ℃贮藏。
1.3 样品前处理和稳定同位素标记
称取水稻样品100 mg,在液氮中研磨,并加入1 mL乙腈,然后置于黑暗条件下4 ℃提取过夜(12 h),离心(10 000 g,20 min,4 ℃),取上清液。样品经SAX-SPE小柱纯化[17]:用3 mL乙腈活化SPE柱后,上样,然后分别用3 mL体积比1∶9 乙腈/水和3 mL体积比9∶1 乙腈/水清洗,最后加入3 mL解吸液(含1%甲酸的乙腈溶液),收集解吸液,在40 ℃水浴加热下,氮气流吹干。
将纯化后的样品用200 μL乙腈复溶后,均分成两份,向每份溶液中依次加入20 μL TEA(10 μmol/mL)和CMPI(20 μmol/mL),涡旋混匀。然后在一份加入20 μL DMED(20 μmol/mL),另一份加入DMED-d4(20 μmol/mL),于40 ℃、1 500 r/min振荡60 min,衍生化反应完成,在40 ℃水浴加热下,氮气流吹干。将轻重标记的样品分别用100 μL体积比1∶9 乙腈/水复溶后合并,进行后续UPLC-MS分析。
1.4 实验仪器和分析条件
质谱为Orbitrap Fusion Tribrid mass spectrometer(美国,赛默飞公司)。液相系统为UltiMate 3000 UHPLC System,(美国,赛默飞公司)。色谱条件:AccucoreTMC18 Columns柱(150×2.1 mm,2.6 μm,美国赛默飞公司),柱温设置为40 ℃。0.1%脂肪酸水溶液作为流动相A,乙腈作为流动相B,梯度洗脱程序:0~5 min,5%B;5~45 min,5%~90%B;45~50 min,90%B;50~52 min,90%~5%B;52~60 min,5%B。流速:0.4 mL/min,进样体积:10 μL。
质谱条件:电喷雾离子源,正离子模式,全扫描模式,扫描范围为m/z180~600,仪器分辨率设置为120 000。离子传输毛细管温度350 ℃,加热块温度300 ℃,喷针电压3.5 kV,鞘流气35 psi,辅助气15 psi。使用数据依赖采集模式(DDA)获取多级质谱信息。通过CID碎裂模式对所筛查到的羧基化合物进行二级质谱分析,将MS3依赖于MS2中最强子离子进行三级分析。隔离宽度m/z2.0,活化时间为10 ms,扫描范围为m/z50~600。MS2信息采集采用30%的归一化碰撞能量,MS3设置为30%的归一化碰撞能量。使用Mass Frontier Server Manager 7.0(美国赛默飞公司)辅助阐明羧基化合物结构。
1.5 数据处理
利用Thermo Compound Discoverer 2.1 Software(美国赛默飞公司)软件对采集的初始数据抽提,得到保留时间(RT)、m/z和Intensity信息。通过计算保留指数(RI)来校正RT[18]。运用Matlab软件对m/z相差4.025 Da、RI一致(轻重标记产物RI相差不超过1%)和峰强度一致(IntensityDMED-labeled/IntensityDMEDd4-labeled:0.76~1.50)的峰对进行匹配以筛选出候选化合物。利用Xcalibur 2.2.0(Thermo Fisher Scientific,USA)软件,根据峰匹配得到的精确分子量信息,推测出标记产物的分子式,质量偏差为10 ppm,元素组成为C、H、O、N、S、Cl。通过本实验室所建立的化学标记代谢物库(Chemically Labeled Metabolite Database,CLMD,http://www.clmdb.cn/)对水稻中的羧基化合物进行比对确认,同时将模拟的分子式在Metlin(https://metlin.scripps.edu/landing page.php)和Massbank(https://massbank.eu/MassBank/)等数据库中检索相关羧基化合物,得到可能的候选物。通过镉处理组与正常组的相对含量比(即Cd/normal)计算差异倍数(Fold Change,FC),使用统计软件IBM SPSS statistics 19.0,先对数据进行单因素方差分析得到p值,再通过综合变异倍数分析(FC≥2 or ≤0.5)和p值(p<0.05)筛选出具有显著性差异的羧基化合物。采用Microsoft Excel绘制热图。
2 结果与讨论
2.1 水稻中羧基化合物的筛查
水稻是地球一半以上人类赖以生存的粮食作物,也是一种重要的模式植物,以水稻为研究对象,通过分析它在镉胁迫条件下的代谢物种类和含量变化情况,有助于探索镉胁迫相关的代谢途径,研究植物中镉胁迫的调控机制。本研究使用了一对轻重标记试剂(DMED和DMED-d4)来标记羧基化合物,通过引入易电离的叔胺基团,由负离子模式转变成正离子模式检测,提高了羧基化合物的离子化效率,增强其质谱检测灵敏度,从而实现水稻中低丰度羧基化合物的分析筛查。具体工作流程如图1所示。
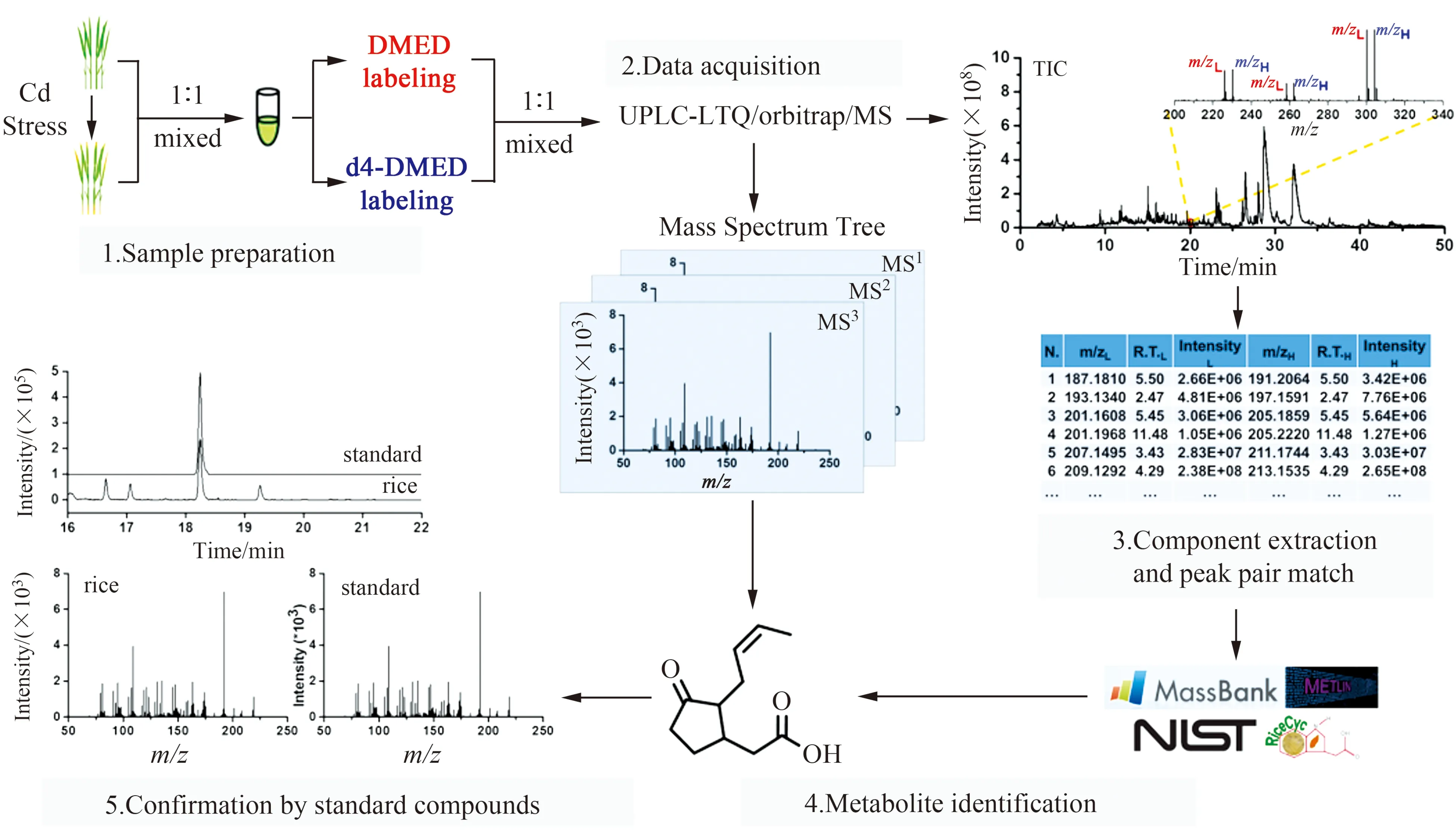
图1 水稻中羧基化合物筛查和定性的流程图Fig.1 Workflow for screening and identification of carboxylic compounds in rice
首先,我们将正常生长和镉胁迫处理后的样品提取液等比例混合,再等分成两份,分别用DMED和DMED-d4进行标记。然后,将轻标和重标标记的样品按照1∶1(V/V)进行混合,进行LC-Orbitrap MS分析。图2A为DMED/DMED-d4标记的水稻样品的总离子流图。在全扫描模式下,当抽提出的峰对符合以下三个条件:“轻”“重”标记的质量数相差4.025 Da、保留指数(RI)[18]一致和峰强度比一致时,认为是潜在的含羧基代谢物。如图2B中的提取离子色谱图所示,m/z为187.1799和191.2053的峰强度和保留指数一致,这类化合物被认为是潜在的羧基化合物。同时,轻重标记样品的MS2信息和相应碎裂片段(图2C)进一步展示了化合物的结构信息。

图2 经DMED/DMED-d4标记的水稻样品在LC-Orbitrap MS全扫模式下分析。(A)总离子流色谱图;(B)m/z187.1799和m/z191.2053的己酸提取离子色谱图;(C)标记的己酸二级质谱图。Fig.2 DMED and DMEDd4 labeled rice analyzed by LC-Orbitrap MS under full scan mode.(A) Total ion current chromatogram of rice sample labeled by DMED and DMED-d4;(B) Extracted ion chromatograms of caproic acid at m/z187.1799 and m/z191.2053 from DMED/DMEDd4-labeled rice;(C) MS2 spectra of DMED/DMED-d4-labeled caproic acid.
运用该方法,实验一共在水稻样品中发现了1 444个可能的羧基化合物,表明水稻中存在着丰富的羧基化合物。其中,根部和地上部分中的代谢物种类有很大差异,分别测到771和1 044个羧基化合物,有371个化合物在根部和地上部分中均被检测到(图3A)。之前的报道在水稻中检测到的羧基化合物种类不超过100种,本文实验结果极大扩展了水稻中羧基化合物的种类。

图3 韦恩图显示了镉胁迫下根部(黄色)和地上部分(紫色)及二者共同含有的羧基化合物数量。(A)CIL-UPLC-MS分析中检测到的羧基化合物数量;(B)根部和地上部分中含量有显著差异性变化的羧基化合物数量;(C)根部和地上部分中含量显著上调或下调的羧基化合物数量。Fig.3 Venn diagram showing the number of metabolites selected for Cd treatment in root(yellow),in shoot(purple) and commonly for both tissues.(A) Metabolites detected in CIL UPLC-MS analysis;(B) Metabolites showing a significant change;(C) Significant up-regulated and down-regulated metabolites in root and shoot under Cd stress.
2.2 水稻中羧基化合物的定性分析
本文对筛查到的羧酸化合物进行了定性分析。首先,将潜在羧基化合物的精确分子量、RI、MS2等信息与CLMD中的标准品信息进行比对,有72个羧基化合物可以与Chemically Labeled Metabolite Database(CLMD)(http://www.clmdb.com)中对应的标准品相匹配,同时通过标准品进一步验证(level 1,No.1-72);根据MS2等信息,推测出13个可能的羧基化合物结构(level 2,No.73-85);将化合物的模拟分子式在Metlin(https://metlin.scripps.edu/)等数据库中进一步搜索,可以获得563个潜在的羧基化合物信息(level 3,No.86-648);通过Xcalibur软件,对剩余的796个化合物进行搜索,其中有691个化合物可以与精确的分子式匹配(level 4,No.649-1339);另外105个化合物没有得到精确的分子式(level 5,No.1340-1444)。level 4和level 5中的化合物,可能是水稻中存在的一些新的羧基化合物,将在以后对其做进一步的确认和研究。
2.3 水稻中羧基化合物的相对定量分析
进一步探索了镉胁迫条件下水稻中羧基化合物含量的差异,对筛查到的1 444个潜在羧基化合物进行相对定量分析(图4)。运用多元统计方法处理数据,结果表明,在镉胁迫下水稻内检测到486种羧基化合物含量具有显著性差异,根部和地上部分中分别有159和365种羧基化合物含量表现出显著差异,其中有38种在二者间均表现出显著变化(图3B)。

图4 水稻中含羧基化合物的相对定量工作流程Fig.4 Overview of the procedure ford relative quantitation of carboxylic metabolites in rice
实验发现根部中检测到的大部分的羧基化合物含量显著降低,而地上部分中绝大多数的羧基化合物含量都显著升高。图5A、5B显示了镉胁迫条件下根部和地上部分中羧基化合物的相对含量变化。在根部中,有38%(61个)的羧基化合物含量显著升高,而62%(98个)的羧基化合物含量显著降低。然而,在地上部分中,有95%(344个)羧基化合物含量都是显著升高,只有5%(15个)的羧基化合物含量显著降低(图3C)。这表明水稻根部和地上部分在镉胁迫时可能在羧酸代谢水平采用了不同的应对策略。

图5 镉胁迫条件下根部(A)和地上部分(B)中羧基化合物含量变化的火山图。通过t检验计算各代谢物的p值,通过镉处理组与正常组的相对含量比(即Cd/normal)计算fold change(FC)。以p<0.05,FC≥2或≤0.5为临界值,红色点表示镉处理组中含量升高的羧基代谢物,蓝色点表示镉处理组中含量降低的羧基代谢物。黑点表示含量没有明显变化的羧基代谢物。Fig.5 Volcano plots of the carboxylic acids in root(A) and shoot(B) under Cd stress.The p -value of each metabolite was calculated from t-test,and the fold change(FC) was calculated from the peak ratios of Cd treated group and normal group(i.e.,Cd/normal).Using a cut-off value of p<0.05 and FC≥2 or ≤0.5,the red points represent the metabolites with higher concentrations in the Cd treated group,and the blue points represent the metabolites with lower concentrations in the Cd treated group.The black points represent the metabolites with no significant change.
图6呈现了不同定性标准下的Cd胁迫后同一羧基化合物在根部和地上部分中的相对含量变化情况。5个level中显著差异的羧基代谢物个数分别为:level 1(24个)、level 2(3个)、level 3(219个)、level4(206个)、level 5(34个)。可以看出,同一化合物在根部和地上部分中变化情况各异,如苯甲酸仅在地上部分中含量上升,而肉桂酸仅在根部中含量上升。值得注意的是,差异倍数增大或减小明显的代谢物多在level 3和level 4水平,例如变化10倍以上的代谢物共有70个,level 3和level 4的占比分别是21%(15个)和69%(48个)。我们推测level 4中含有新的羧基化合物,且可能在镉胁迫中发挥重要作用,对于将来缓解镉胁迫的危害有重要意义。

图6 镉胁迫下显著差异性含羧基代谢物的热图。以p<0.05,FC≥2或≤0.5为临界值,红色表示含量升高,蓝色表示含量降低,白色表示含量没有明显变化,灰色表示未检测到。Fig.6 Heat map of metabolites with significant changes under Cd stress.Using a cut-off value of p<0.05 and FC≥2 or ≤0.5,the red points represent the metabolites with higher concentrations in the Cd treated group,and the blue points represent the metabolites with lower concentrations in the Cd treated group.The white points represent the metabolites with no significant change.The gray points represent the undetected metabolites.
2.4 显著性差异化合物的代谢网络分析
为了进一步揭示镉胁迫对水稻根部和地上部分中代谢稳态的调节,对鉴定出的代谢物进行了代谢网络分析。将筛查到的72个可以和标准品比对确认的羧基化合物(level 1)导入KEGG数据库(http://www.genome.jp/kegg/pathway.html)中,获得了这些化合物主要参与的代谢途径。结果表明,其中37种羧基代谢物主要富集在8条代谢通路,分别为苯甲酸族化合物代谢、苯丙氨酸代谢、苯基丙酸生物合成、脂肪酸生物合成、脂肪酸降解、不饱和脂肪酸生物合成、亚麻酸代谢和花生四烯酸代谢。在这8条路径中,镉胁迫条件下根部中绝大多数可以检测到的羧酸代谢物含量是无显著变化或显著降低的,而在地上部分中,含量发生显著变化的代谢物的变化趋势都是显著升高的。这些结果进一步说明水稻根部和地上部分在镉胁迫时可能在羧酸代谢水平采用了不同的应对策略。

图7 镉胁迫下水稻内部分含羧基化合物的代谢网络图。以p<0.05,FC≥2或≤0.5为临界值,红色表示含量升高,蓝色表示含量降低,白色表示含量没有明显变化,灰色表示未检测到。Fig.7 Metabolic network map of metabolites with significant changes under Cd stress.Using a cut-off value of p<0.05 and FC≥2 or ≤0.5,the red points represent the metabolites with higher concentrations in the Cd treated group,and the blue points represent the metabolites with lower concentrations in the Cd treated group.The white points represent the metabolites with no significant change.The gray points represent the undetected metabolites.
在重金属胁迫条件下,有机酸能够通过与金属离子螯合来解毒,是重要的植物初级代谢产物。镉胁迫条件下,在苯甲酸族化合物和苯基丙酸的代谢路径中共检测到6种有机酸。其中肉桂酸在根部中的含量升高,其下游代谢通路产物,苯甲酸和对香豆酸的含量在地上部分中均升高,进一步促进次生代谢,诱导大量酚类物质合成。酚类物质是植物体中的重要次生代谢产物,可与金属离子发生络合反应,或者通过去除自由基直接减弱氧化损伤,从而增强植物体自身防御系统[10]。在苯基丙酸生物合成路径中,存在一系列酚酸,我们推测镉可能诱导一系列中间代谢酚酸产物(咖啡酸和阿魏酸)的合成来响应胁迫。
酸性植物激素在镉胁迫毒害下调节植物适应和存活起着关键作用。实验中检测到两种羧基类植物激素生长素(IAA)和脱落酸(ABA)在镉胁迫水稻地上部分中的含量均升高。ABA与水稻的镉耐受性密切相关,水稻受到镉胁迫后,为了阻止镉在体内的快速转运,会分泌大量ABA,从而抵抗胁迫。同时ABA的含量改变也会诱导IAA的含量变化,两者共同协调,帮助植物抵御胁迫[1,9,10]。
脂肪酸(FAs)的生物合成主要发生在叶绿体基质中,饱和FAs经过去饱和作用可形成不饱和FAs。在植物细胞膜中,不饱和FAs含量的高低直接决定膜的流动性,影响植物对非生物胁迫的抗性。Chaffai等[20]发现脂类化合物与镉胁迫条件下植物的生长调节密切相关。8条代谢路径中有27种FAs化合物,其中地上部分中有4个FAs代谢物含量显著上升,根部内有1个FAs代谢物含量上升,4个含量下降。脂氧合酶(LOX)可催化脂质过氧化形成丙二醛(MDA)等,在胁迫条件下,FAs降解和不饱和脂肪酸的代谢活跃,LOX活性明显升高[21]。实验中观察到根部中不饱和FAs代谢通路上仅有花生四烯酸的含量上升,同时它的下游产物12-HETE和Leukotriene均能监测到,表明镉胁迫条件下,LOX可能催化了FAs的氧化裂解,导致膜脂过氧化程度升高,细胞膜的透性增加,造成氧化损伤。
3 结论
本文运用稳定同位素标记结合UPLC-MS联用技术,全面研究了水稻中羧基化合物的组成。最终发现了1 444个潜在羧基化合物,可以定性出其中的648个化合物(包括72个标准品确认,13个多级质谱解析和563个数据库检索获得)。同时,通过统计学分析得到镉胁迫下含量发生显著性变化的化合物。结果表明,镉胁迫条件下与正常条件下生长的水稻相比,水稻根部中大部分(62%)的羧基化合物含量显著降低,而地上部分中绝大多数(95%)的羧基化合物含量显著升高。进一步对这些化合物进行代谢网络分析,发现化合物涉及苯甲酸代谢、脂肪酸的生物合成与降解等8条代谢路径。这8条路径中的羧基代谢也呈现了不同的变化趋势。因此,推测水稻根部和地上部分在响应镉胁迫时,可能在羧酸代谢水平上采用了不同的应对策略。该研究结果为从代谢物水平上揭示植物对镉胁迫的适应机制提供了数据基础,为进一步揭示植物应答镉胁迫的响应机制提供了有力工具。
猜你喜欢
现代临床医学(2022年4期)2022-09-29
理化检验-化学分册(2020年5期)2020-06-15
金属加工(热加工)(2020年12期)2020-02-06
农药科学与管理(2019年5期)2019-08-13
植物营养与肥料学报(2019年12期)2019-03-07
腹腔镜外科杂志(2016年11期)2016-06-01
橡胶工业(2016年2期)2016-02-23
磁共振成像(2015年9期)2015-12-26
民用飞机设计与研究(2015年3期)2015-12-07
中国医学创新(2015年33期)2015-04-27
