
温带草原7种针茅植物根系特征及其对环境因子变化的适应
2022-11-26 03:00袁野梅柳隽瑶高秀丽王仁忠
生态学报 2022年21期
袁野梅,柳隽瑶,高秀丽,薛 璟,4,王仁忠,4,*
1 广东培正学院统计学系,广州 510830
2 陕西省林业科学院黄土高原水土保持与生态修复国家林草局重点实验室,西安 710082
3 中国科学院植物研究所,北京 100093
4 中国科学院大学,北京 100049
在陆地生态系统中,根系不仅是植物获取水分、养分及同化物分配的重要器官,也是植物适应环境变化的重要构件,因此受到广泛关注[1—2]。根系作为水分吸收器官对干旱极为敏感,不仅根系外部形态性状因干旱而产生变化[3—4],根系解剖结构[5—6]和生理调节物积累也表现出一定的变化[7—9],从而形成应对环境变化的综合适应机制。但以往的研究多集中在生态学、解剖学或生理生态学等某一特定层次上,缺乏多学科多层次的系统性研究[10],因此在根系生物量、解剖结构和生理调节等多层次上系统研究植物对环境变化的综合适应机制对探讨未来气候变化下植物适应策略及分布区域变化等有着重要的理论和实践意义[11]。
针茅属植物广泛分布于我国西部干旱和半干旱区的天然草地,是荒漠草原区主要牧草资源。关于针茅属植物的研究报道很多[12—13],主要集中在特定区域或某一种针茅植物的特征[14—17]、草地管理方式对针茅植物根系的影响[18—19]等方面,关于大尺度梯度下多种针茅植物根系性状的比较、与区域环境因子的关系及未来气候变化下适应策略和分布区域变化等尚不明确。本研究以温带草原8个针茅草地的7种针茅植物根系为研究对象,系统研究了针茅属植物根系生物量、根冠比(R/S)、解剖结构和生理调节物积累等性状的变化规律,并分析了根系性状与环境变化的关系,旨在为探究未来气候变化下针茅植物适应策略及不同针茅植物分布区更替趋势等提供科学依据。
1 材料和方法
1.1 研究区自然概况
研究区域涵盖了我国温带主要针茅草原(106°88′—120°12′E,38°02′—49°35′N),从东北部的海拉尔到西南部的盐池高沙窝镇(约1600 km),东西贯穿经度约14°,南北跨越纬度约10°(表1)。海拔高度呈现由东北至西南增加的趋势,位于东北端的海拉尔(HL)平均海拔为679 m,锡林郭勒白音锡勒牧场(BY)和多伦(DL)海拔高度逐渐升高至1000 m以上,达茂旗召河(ZH)和乌拉特后旗(UR)地势隆升,海拔高于1600 m,乌海四合木保护区(WH)、鄂尔多斯草原(OR)及宁夏盐池高沙窝镇(GS)海拔在1300—1400 m之间。
该区域属温带大陆性季风气候,夏季风弱、冬季风强,气候干燥,日照丰富。近50年(1970—2018年)气象数据统计表明区域年平均气温在-0.8—8.5℃之间,最低温多出现在1月,最高温均出现在7月,年均气温总体呈现由东向西及由北至南逐次递增的趋势(表1)。降水量少而不均是限制该区域草原植物生长的首要因子,年降水量分布东多西少,北高南低,西南部乌拉特后旗和乌海年降水量小于150 mm,东北部海拉尔、多伦、白音锡勒牧场年降水量高于350 mm,是西南部样点年降水量的2倍多。春季和初夏(4—6月)会经历较为明显的季节性干旱(1—6月降水量约占全年的30%),降水量峰值出现在7—8月(降水量约占全年的50%)。降水和气温的梯度变化导致植被分布呈现出明显地带性格局,由东北到西南依次为草甸草原、典型草原和荒漠草原或荒漠,土壤主要为钙土或暗栗钙土、栗钙土、风沙土、灰钙土、漠钙土和棕漠土等,主要针茅植物依次为贝加尔针茅、大针茅、克氏针茅、本氏针茅、短花针茅、戈壁针茅和沙生针茅。
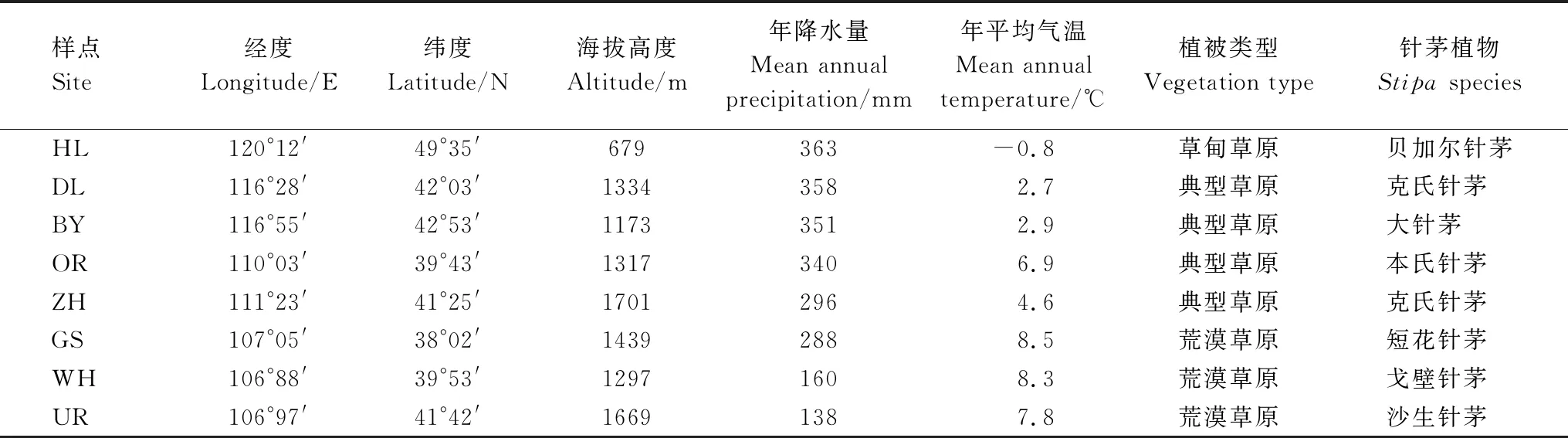
表1 大尺度梯度下实验样点地理位置、气候、地带性植被和主要针茅植物种类.
1.2 实验方法
在研究区内选择8个优势种为针茅属植物且人为干扰较少的天然草地为固定采样点(表1),2018年4月26日—6月11日(旱季)开花期和7月28日—9月5日(雨季)果后期分别进行野外采样。为减少不同样点植物生长节律差异的影响,旱季由南至北(西南部返青早于东北部),雨季由北至南进行调查取样(北部枯死期早于南部)。在各样点选取3个天然针茅草地作为采样区(面积大于1 hm2,且土壤条件基本一致),每个采样区随机设置5个1 m×1 m样方,在样方内取1个包含中等大小针茅株丛的20 cm×20 cm×20 cm土柱,将土柱分成0—10 cm和10—20 cm2层(涵盖了针茅植物90%以上根系生物量),分别装入塑料袋带回实验室。地上部分分成针茅和其他植物,分别装入纸袋,烘干称重。
用1 mm孔径筛子筛出每层土样中的土粒,将植物根装入90目尼龙网袋,用清水冲洗干净。去除杂物和其他植物根系,得到针茅根系样品,用吸水纸将植物根表面的水分吸干。剪下针茅根蘖,称根系样品鲜重。依据颜色、弹性、韧性等将其分为活根和死根,活根再分为吸收根(主要是当年新生1—3序级细根)和运输根(主要是大于3序级的往年细根)[20—21],用信封分装,放入烘箱,105℃杀青1 h,65℃烘48 h,称干重。根冠比(R/S)为地下根系生物量与地上生物量之比。
在每个土柱中取2—3段针茅植物活根(从根尖开始5 cm长,主要是当年生的吸收根)放入FAA (冰醋酸∶甲醛∶70%乙醇=1∶1∶18)固定液固定。采用石蜡切片法[5],经过软化、抽真空、脱水、透明、浸蜡以及石蜡包埋,用切片机(Leica,RM2235,Germany)切成8—10 μm切片,最后染色制片。利用NIS-Elements(Nikon,Japan)图像分析软件测定根切片的导管(Vea)和中柱(Stea)直径,表皮厚度(Ept)、皮层厚度(Cort)、内皮层厚度(Ent)、内皮层径向壁厚度(Rawt)及根直径(Rd)等[10]。
烘干称量后的各层活根样品经球磨仪粉碎过100目筛,粉末样品用于测定抗旱生理指标脯氨酸(Pro)和可溶性糖(SS)含量。取0.1—0.5 g样品粉末加入磺基水杨酸(10 mL,3%)进行提取,经滤纸过滤后取上清液2 mL,茚三酮法测定Pro含量[10]。取50 mg样品粉末置于80%乙醇萃取液,85℃静置1 h。然后高速离心(3000转)10 min,如此反复用80%乙醇萃取三次,合并上清液,加入活性炭脱色,在真空干燥器里干燥,最后用蒸馏水定容,蒽酮比色法测定SS含量[10]。
在各样方中采集0—20 cm土样,风干后研磨粉碎过0.15 mm筛。取0.1—1.0 g土样,重铬酸钾滴定法测土壤碳含量。取0.1—1.0 g样品,凯氏定氮仪(FOSS2200,Denmark)测定土壤全氮含量。
1.3 数据处理
采用Excel 2016和R 3.4.1软件对各样点旱季和雨季针茅植物地上地下生物量、R/S、Vea、Stea、Ept、Cort、Ent、Rawt、Pro和SS含量等数据进行统计分析,方差分析比较各样点间及同一样点旱雨季间针茅根系地上地下生物量、R/S、Vea、Stea、Ept、Cort、Ent、Rawt,Pro和SS含量的差异。利用一元线性回归分析了针茅根系生物量、R/S、Ept/Rd、Cort/Rd、Ent/Rawt、Vea/Stea、Pro和SS含量与旱季和雨季降水量和气温间的关系(旱季和雨季降水量和气温分别指1—6月和1—8月降水量和平均气温,涵盖了所有时段降水和气温对植物性状的影响)。采用独立性较高的旱季和雨季平均气温、降水量、海拔作为环境变量对针茅植物上述性状进行偏相关分析(控制其中两个因子,分析各指标与第3个因子的关系)。本研究气象数据来源于中国气象数据网(http://www.worldclim.org) 1970—2018年地面常规观测资料。
2 结果
2.1 根系生物量和根冠比
无论是旱季还是雨季,除BY样点的大针茅具有较高的根系生物量外,在由东北到西南的大尺度区域内针茅植物根系生物量基本呈现增加的趋势(图1)。除了旱季OR与ZH、雨季DL、OR与ZH相邻样点间外,其他各样点0—10 cm土层根系生物量差异均达到显著水平(P<0.05),旱季DL和ZH样点的克氏针茅0—10 cm土层根系生物量差异也达到了显著水平。旱季和雨季,HL和DL样点0—10 cm土层根系生物量仅为UR和BY样点的7%和17%。8个样点10—20 cm土层针茅植物根系生物量变化基本与表层一致,BY样点根系生物量最大,显著高于其他样点(P<0.01),旱季约为最低样点(HL)的6.1倍、雨季约为最低样点(DL) 5.3倍。旱季OR与UR、ZH与GS间及雨季GS与WH间10—20 cm土层根系生物量差异不显著(P>0.05)。旱季和雨季各样点0—10 cm土层针茅根系生物量均显著大于10—20 cm土层(P<0.05),且表层平均根系生物量为总根系生物量的85%。在研究区域内,针茅植物R/S差异显著,HL和DL样点R/S最低,旱季由BY至WH逐渐降低,最低值(DL)仅为BY和UR样点的20%和13%,UR样点针茅R/S最大。雨季由东北到西南R/S基本呈增加的趋势,UR样点的R/S约为HL样点的8倍。
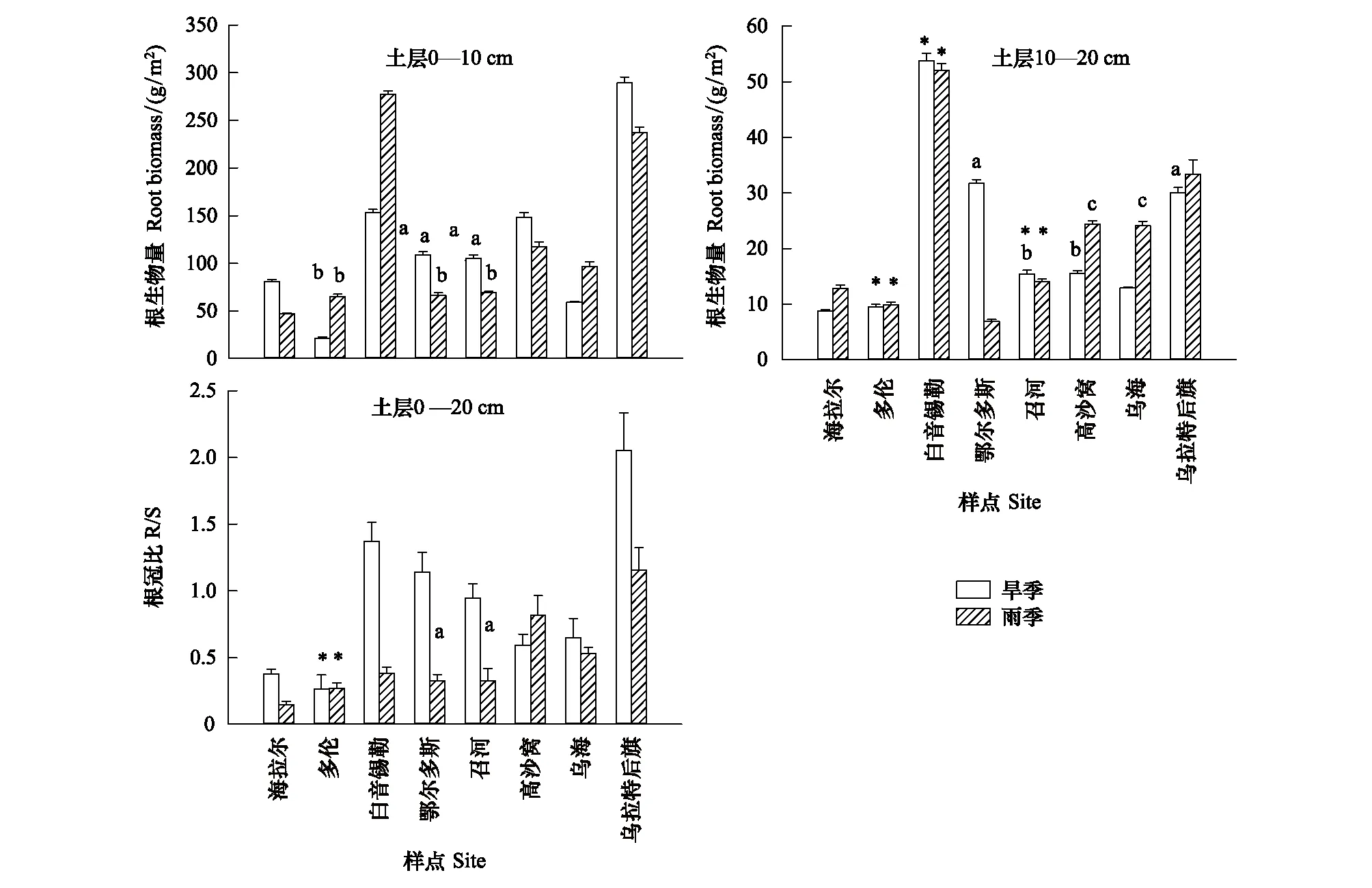
图1 大尺度梯度下旱季(DS)和雨季(RS)各取样点0—10 cm和10—20 cm土层针茅植物根系生物量和根冠比的比较(平均值±标准误)
2.2 运输根和吸收根
旱季和雨季,各针茅植物0—10 cm土层根系中运输根占比最大,平均为64.4%和65.4%,且由东北至西南呈逐渐增大的趋势(图2),吸收根平均占比分别为20.7%和17.4%,其变化趋势与运输根相反。在10—20 cm土层根系中,运输根平均占比下降(图2),旱季和雨季分别为31.0%和38.3%,二者差异显著(P<0.05),吸收根比例分别增大到49.0%和36.2%,且旱季显著大于雨季(P<0.01)。雨季下层土壤中死根比例有所增加,达到25.4%,是旱季的3倍。旱季和雨季,BY、ZH、OR和WH样点针茅植物0—10 cm的吸收根与运输根比(Ra/Rt)差异显著(P<0.05),BY样点大针茅旱季Ra/Rt约是雨季的2倍。GS和HL样点10—20 cm土层中Ra/Rt旱季和雨季差异显著,GS样点短花针茅旱季是雨季近3倍。
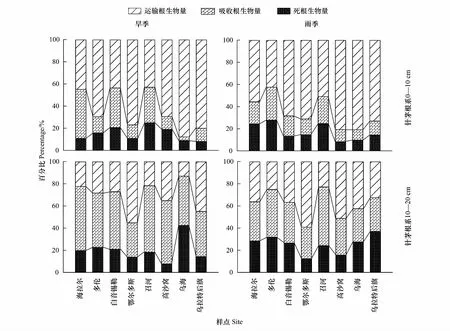
图2 旱季(DS)和雨季(RS)针茅根系0—10 cm和10—20 cm土层运输根(Rt)、吸收根(Ra)和死根(Rd)生物量百分比堆叠图
2.3 根解剖结构
在实验区域内,旱季和雨季针茅植物根直径(Rd)、表皮厚度(Ept)和皮层厚度(Cort)表现出不同的变化规律(表2)。旱季针茅Rd、Ept和Cort基本表现为由东北至西南呈下降趋势,但在最西部的UR各指标均出现明显上升,而雨季总体呈由东北到西南的上升趋势。旱季HL样点Rd比雨季高45% (P<0.01),而其他样点均为雨季高于旱季。旱季东北部3个样点的Ept和Cort均显著高于雨季(P<0.05),而西南部5个样点则低于雨季。

表2 旱季和雨季针茅根直径、表皮厚度及皮层厚度 (平均值±标准误)
各针茅植物根系的内皮层均有不同程度的马蹄形加厚,旱季针茅根系内皮层(Ent)和径向壁厚度(Rawt)由东北至西南有下降趋势,但在UR样点显著增加(P<0.01),而雨季无明显趋势(表3)。旱季东北部3个样点的Ent和Rawt均显著高于雨季(P<0.05),其余5个样点的Ent则是雨季高于旱季。8个样点中,针茅植物中柱(维管束)直径(Stea)的种间差异显著,但无明显规律。导管直径(Vea)的变化与Ent相似,但雨季大体由东北至西南逐渐上升(表3)。除东北部3个样点外,其余5个样点的Stea和Vea基本为雨季高于旱季。
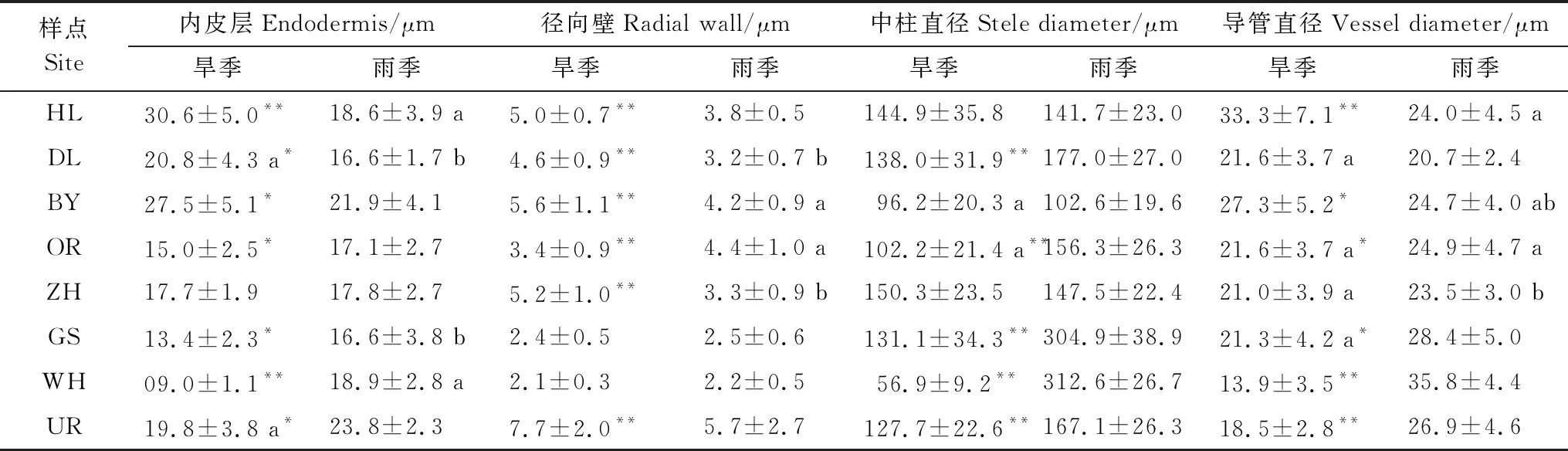
表3 旱季和雨季针茅根内皮层、径向壁厚度、中柱和导管直径变化(平均值±标准误)
根表皮厚度和皮层厚度与根直径比例(Ept/Rd、Cort/Rd)可以直观地反映植物根对表皮的物质投入比例及其适应功能。各样点针茅植物Ept/Rd和Cort/Rd基本为旱季大于雨季(ZH样点Cort/Rd除外),WH样点戈壁针茅旱雨季差异最大,旱季Ept/Rd约为雨季的3倍(P<0.01),贝加尔针茅(HL)、克氏针茅(DL,ZH)和本氏针茅(OR)季节差异不显著(P>0.05)(表4)。旱季各植物Ept/Rd处于波动状态,种间差异显著。雨季Ept/Rd总体呈现由东北向西南逐渐下降的趋势,最大值为HL样点贝加尔针茅,是最小值WH样点戈壁针茅的1.6倍(P<0.05)。
内皮层与径向壁厚度比(Ent/Rawt)在一定程度上反映了根保水能力的强弱。各样点针茅根Ent/Rawt基本是雨季大于旱季(表4),且差异显著(P<0.01)(GS样点除外)。无论旱季还是雨季GS和WH样点的Ent/Rawt均显著高于其他样点。导管面积与中柱面积比(Vea/Stea)体现了根系的输水效率,该值越大说明根输水能力越强。旱季和雨季间各种针茅植物Vea/Stea差异较小,且东北部3个样点针茅植物间差异均不显著(P>0.05),最西部的UR样点Vea/Stea显著高于其他样点(P<0.01)。

表4 旱季和雨季针茅根表皮厚度、皮层厚度与根直径比、根内皮层与径向壁厚度比和导管与中柱面积比
2.4 根系生理调节物
各样点针茅根系脯氨酸含量(Pro)波动很大,由东北到西南没有明显的变化规律(表5),最西部的UR样点Pro最高。旱季和雨季仅有不到一半样点各土层根系Pro差异显著,然而,无论是旱季还是雨季,各样点10—20 cm土层根系Pro含量均高于0—10 cm土层(除最西部的UR样点外)。3/4样点10—20 cm土层根系Pro旱季高于雨季。与Pro变化相似,由东北到西南针茅植物根系可溶性糖含量(SS)没有呈现明显的增减趋势(表5)。除OR样点外,各样点根系SS呈现显著的季节性差异(P<0.05),西南部3个样点旱季0—10 cm土层根系SS含量显著高于雨季。除HL样点外,10—20 cm土层根系SS含量均为旱季高于雨季。

表5 旱季和雨季0—10 cm和10—20 cm土层针茅根系脯氨酸和可溶性糖含量(平均值±标准误)
2.5 根系性状与环境因子的相关分析
偏相关分析显示大尺度梯度上针茅植物根系性状与环境因子(降水量、平均气温和海拔高度(Alt))间存在显著的相关性(表6)。0—10 cm土层针茅根系生物量与P1—6、P1—8、T1—6、T1—8和Alt均呈极显著负相关关系(P<0.01); 10—20 cm土层根系生物量与P1—8和Alt呈显著负相关关系(P<0.01),但与T1—6呈显著正相关关系(P<0.05)。旱季针茅植物R/S仅与P1—6显著负相关(P<0.01),但雨季R/S与P1—8和Alt均达到了极显著负相关水平(P<0.01),且与T1—8呈正相关关系(P<0.01)。降水量、平均气温和海拔高度对各土层针茅运输根生物量均呈负效应,与旱季0—10 cm土层运输根生物量呈极显著负相关关系(P<0.01)。旱季针茅植物吸收根生物量受到降水量和海拔高度的极显著负效应(P<0.01),T1—6仅对表层吸收根生物量产生极显著的正效应(P<0.01),雨季环境因子对针茅植物吸收根生物量的影响较旱季明显减弱。

表6 大尺度梯度上根系生物量、根冠比、吸收根和运输根生物量与气候和海拔高度(Alt)等因子的偏相关分析
环境因子对针茅植物根系解剖性状有显著影响(表7),P1—6对Ept/Rd、Cort/Rd及P1—8对Cort/Rd、Ent/Rawt、Vea/Stea均有显著负效应(P<0.01),温度(T1—6、T1—8)对上述各指标的影响不尽相同。降水量显著影响旱季各土层和雨季10—20 cm土层Pro含量(P<0.05),温度对各土层根系SS有极显著负效应(P<0.01)。海拔高度对表层根系Pro和SS含量有显著正效应(P<0.01)。

表7 旱季和雨季针茅根系解剖和生理调节特征与降水、温度和海拔的偏相关分析
3 讨论
3.1 针茅植物根系性状差异及其适应策略变化
植物应对干旱的适应策略虽然复杂多样,但主要包括以下两种途径,一是通过改变根系性状(如增加R/S、减小内皮层厚度等)最大程度增加吸水和保水能力,二是改变植物叶片性状(如减少气孔密度、增加比叶重等)减少水分散失[9]。针茅植物根系对干旱极为敏感,研究其耐旱性状变化对揭示针茅植物的适应策略和机制具有重要的理论和实践意义。在本研究的8个样点中,由东北至西南针茅根系生物量呈现波动增长趋势(图1),即体现了针茅植物的种间差异,也反映了它们对干旱的趋同适应性。这一趋势与其他报道基本一致,Gao等对分布于不同沙丘部位两种沙蒿根系的比较发现生长在沙丘顶部(干旱贫瘠)的白沙蒿(Artemisiasphaerocephala)比生长在沙丘底部(水分和养分相对较好)的褐沙蒿(A.intramongolica)具有更发达的根系,不仅分布较深(140 cm以上),而且生物量也高[4]。Wang等的研究显示在典型草原和荒漠草原区随干旱增加羊草根系生物量呈现增加的趋势[22]。干旱区生态系统中发达的根系增强了植物吸水和保水能力,进而增强了植物适应干旱的能力[3—4]。多数样点针茅植物在旱季维持较高的表层(0—10 cm)根系生物量,这主要是由于表层运输根占比高(约65%,图2),其木质素含量高周转率低[2]。针茅植物深层(10—20 cm)根系生物量旱季和雨季差异不显著,其原因是深层根系中周转率较高的吸收根(或细根)占比大(图2),相对稳定的土壤环境(主要是温度和水分)导致吸收根新生或死亡的波动较小[4]。深层根系对降水的响应较弱表明针茅有着较强的耐旱性,其深层根系的相对稳定维持了群落生物量的稳定[23]。
Ma等的研究显示内蒙古草原群落R/S与降水量及温度没有显著相关性[13],但在本研究区域内,各针茅植物R/S普遍是旱季大于雨季,且旱季和雨季R/S与降水量均呈极显著负相关,这与Schenk等[3]、Mokany等[24]和Wang等[22]的结果基本一致。旱季较高的R/S增强了植物吸水、保水及碳水化合物贮藏能力,有助于植物度过干旱逆境[25]。与以往报道不同的是在由东北至西南的样带上,旱季针茅植物R/S并没有随年均降水量的减少而增加(图1),而是由BY样点至WH样点呈逐渐下降的趋势,这可能与该样带温度变化有关,BY样点至WH样点年均温逐渐增加,升温导致根系呼吸增强,降低生物量积累,同时升温有助于植物生长和地上生物量增加,结果导致这些样点针茅R/S下降。升温与干旱胁迫对植物根系交互影响比较复杂,今后需要增加控制实验才能准确甄别。
3.2 针茅植物根系功能和解剖结构及其对环境的适应
干旱不仅改变植物R/S,也改变根系的功能结构。本研究根据文献[20—21]、根序级及解剖结构将针茅根系分为吸收根和运输根两部分,划分虽然简单,却能较好地区分不同等级根系在适应干旱过程中的功能作用。表层根系中运输根占比较大,深层吸收根比例较高,且旱季大于雨季(图2),表明二者在应对干旱逆境时其结构和功能发生了分异。旱季吸收根生物量与降水量呈显著负相关关系也证明了植物根系功能组成和适应策略的差异(表6)。Wang等关于干旱胁迫增加深层羊草新生根(即吸收根)生物量的报道也支持上述结论[25]。但Gao等报道在本实验区域内两种沙蒿植物细根生物量分布与土壤水分没有显著的相关性[4],因为沙蒿根系很深,主要利用深层地下水,受土壤干旱的影响较小。目前,关于植物粗根和细根的研究主要集中在碳周转率方面[2,21],而关于根系性状与水分关系的报道也多集中于作物研究[1,7],因而关于草原和荒漠等干旱区不同植物类群根系功能分异的系统研究仍需加强。
解剖结构的变化是植物长期生活在特定环境下而形成的稳定性适应策略,在本研究区域内针茅植物根表皮厚度(Ept)和皮层厚度(Cort)的变化与根直径一致(表2),但是,旱季和雨季变化趋势不同。旱季由东北至西南随降水量的减少Ept和Cort变薄,Chimungu等研究显示Ept和Cort变薄、细胞增大等有利吸收土壤水分,并横向输送至导管[26]。在最干旱的UR样点针茅根系Ept和Cort均维持较高水平,Ept和Cort增厚虽然降低水分横向输导,但防止了根内水分由中柱倒流回皮层,增强了保水能力[27]。雨季由东北至西南Ept和Cort则呈增加趋势,Ept和Cort增加,防止根内水分和养分倒流[27]。以往研究主要涉及特定时期(或一次取样)植物解剖结构,关于解剖结构的季节性差异报道甚少[10]。本研究发现旱季和雨季不同气候区域针茅植物根系Ept和Cort呈现不同的变化规律,东北部3个样点Ept和Cort旱季大于雨季(表2),因为在这些样点区域春季和夏初具有明显干旱胁迫现象,增加Ept和Cort厚度可以起到保水作用,雨季这些区域降水量充沛,Ept和Cort厚度减少可以增加吸水的能力。西北部5个样点Ept和Cort雨季大于旱季,这些区域常年处于干旱缺水状态,即使雨季降水量显著增加,水分仍然匮乏,雨季增加Ept和Cort厚度可以增强根系保水能力,有效应对干旱。旱季针茅根Ept/Rd和Cort/Rd与降水量呈显著负相关也证明了这一点。
内皮层(Ent)和径向壁(Rawt)调控皮层和中柱间水分和养分的运输,控制水分和养分进入中柱,也防止中柱内溶质倒流回皮层[27]。在由东北至西南的样带上,针茅植物根系内皮层(Ent)和径向壁(Rawt)厚度变化与Ept和Cort基本相似(表3),其功能作用也是类似的,这里不再赘述。WH样点戈壁针茅和GS样点短花针茅的Ent/Rawt显著高于其他针茅植物(表4),体现出了较强的耐旱保水能力。水分通过横向运输到达中柱导管进行轴向运输,Vea/Stea的大小在一定程度上反映了根系轴向水分运输能力[5—6]。本研究中,针茅植物中柱直径和维管束直径与内皮层具有相似的变化趋势,旱季由东北向西南逐渐下降,而雨季逐渐增加(表3),且多数样点,旱季中柱(Stea)和导管(Vea)直径小于雨季,干旱条件下针茅根导管直径减小有助于根内水分贮存,为地上部分生长提供保障。Richards等[28]的研究支持这个观点,该研究表明春小麦种子根部轴向水分运输阻力由于导管直径减小而增大,节约了小麦生长早期对水分的消耗,有利于其之后的生长并实现丰产。偏相关分析(表7)表明雨季针茅根Vea/Stea与降水呈负相关关系(P<0.05),这也证明在干旱区随水分条件的改善针茅植物会选择增大导管直径的方式来提高导管占中柱的相对面积,增强根系水分运输效率。中柱和维管束直径大小与植物耐旱性关系存在争议,其在不同植物类群和不同生长期的功能作用也不同[10],因而今后对不同植物类群及不同生长期的Stea和Vea的比较研究是必要的。
3.3 针茅植物生理调节机制差异
脯氨酸(Pro)是理想的渗透调节物质,相较于可溶性糖(SS)更加灵活,可以在胁迫发生时迅速累积[8]。本研究中,各针茅植物间Pro和SS含量均处于较大的波动状态,最干旱的UR样点具有相对较高的Pro和SS含量(表5),表明各针茅物种根系渗透调节物质及其功能存在明显的种间差异。Guo等的研究显示大尺度水分梯度上植物Pro和SS含量与干旱指数(SPEI)显著相关,且不同植物功能群间差异显著[10]。本研究发现针茅植物深层根系Pro和SS含量显著高于表层(表5),这主要是深层根系基本为新生吸收根,生理活性强,是渗透调节的主要参与者。与Munns的报道相反,本研究中多数样点针茅根系Pro旱雨季差异不显著,而SS差异显著(表5),说明SS在针茅植物应对干旱过程中的功能作用可能比Pro更加活跃或二者对不同环境因子的响应存在差异。偏相关分析结果显示针茅植物根系Pro含量与降水量呈显著负相关,而SS含量却与均温呈显著的负相关关系(表7),表明针茅植物对各环境因子的响应机制不尽相同。Guo等研究表明在干旱草原区乔木Pro和SS存在着权衡关系,即在应对干旱时二者的功能是不一致的[10]。Pro和SS作为快速响应逆境胁迫的重要调节物对环境因子(干旱、升温等)变化反应迅速,这也增加了其不稳定性,因此,要明确自然条件下他们的功能作用及相互关系需要做严格的控制实验。
4 结论
在由东北至西南1600 km的研究区域内,针茅属植物呈现明显的更替分布(间或镶嵌分布)状态,总体表现为随降水量的逐渐下降针茅植物的抗旱特征增强或适应策略趋于复杂(采用多种抗旱策略),旱季和雨季不同针茅植物根系有着不同的适应策略。针茅根系各性状与环境因子的偏相关分析表明贝加尔针茅、大针茅及克氏针茅受干旱制约显著,克氏针茅和大针茅对降水高度敏感,雨季降水能够促进其快速生长。沙生针茅、短花针茅、戈壁针茅、本氏针茅通过增大根冠比、加大根系保水和吸水能力、增加渗透调节物质累积等途径适应干旱,这些变化可能为预测未来气候变化[29]下针茅植物分布区域变化及防风固沙能力等提供参考依据[30—31]。
猜你喜欢
中国交通信息化(2022年7期)2022-10-27
成都信息工程大学学报(2022年3期)2022-07-21
山西农业大学学报(自然科学版)(2021年4期)2021-09-23
土壤(2021年1期)2021-03-23
福建林业科技(2020年4期)2020-12-31
今日农业(2020年19期)2020-12-14
安徽农学通报(2020年7期)2020-05-26
农家致富顾问·下半月(2020年2期)2020-05-11
现代农业科技(2019年22期)2019-12-25
现代农业科技(2017年16期)2017-09-22
