
稻纵卷叶螟幼虫驯化产生的热适应能力的继代效应
2022-11-28 08:31李明珠刘向东
昆虫学报 2022年10期
李明珠,刘向东
(南京农业大学植物保护学院昆虫学系,南京 210095)
全球气温呈明显上升趋势,与1850-1900年间相比,全球地表温度在2001-2020年的20年间升高0.99℃,在2011-2020年的10年间升高1.09℃,预计2021-2040年间将上升1.5℃(IPCC,2021)。全球变暖影响生物种群的分布、数量和发生时间等;相应地,生物也在快速进化以适应温度的变化(Parmesan and Yohe,2003;Bradshaw and Holzapfel,2006;Geertsetal.,2015)。多世代经历高温可提高昆虫的耐热能力(Cavicchietal.,1995;Guetal.,2019;Quanetal.,2020)。在连续多年的高温胁迫下,昆虫会发生相对快速的耐受性进化和种群间分化(Cavicchietal.,1995)。用高于环境温度4℃的高温连续选择处理水蚤Daphniamagna,2年后水蚤的耐高温能力显著提高;测定1955-1965年间沉积于湖底的水蚤的临界高温值CTMax也显著低于1995-2005年间沉积的水蚤,表明自然水蚤种群也随全球变暖发生了耐热能力提高的进化(Geertsetal.,2015)。烟粉虱Bemisiatabaci对热选择也有很强的反应,其热耐受性的狭义遗传力在0.47~0.51之间(Maetal.,2014)。在全球变暖背景下,英国两种灌木蟋蟀Conocephalusdiscolor和Metriopteraroeselii的分布区域在不断由南向北和由海边向内地扩张,并且种群中长翅或超长翅型个体比率显著提高,从而有利于种群采用快速扩散方式应对气候变暖的胁迫(Thomasetal.,2001)。这些研究结果表明部分生物在遗传进化上发生了应对温度升高的明显变化。但是也有物种不能在极端环境中获得适应力并将其遗传给后代。例如,高海拔地区的果蝇Drosophilanigrosparsa不能通过产生进化上的遗传变化来适应热胁迫,种群受全球变暖的影响大(Kinzneretal.,2019)。热带地区的一些果蝇物种的遗传变异也较低,体内缺少与热适应相关的遗传变异,锻炼后提高的热适应能力仅是一种表型可塑性,没有传代能力(Kellermannetal.,2009)。昆虫能否产生热适应性与昆虫种类和所处地理环境有关(Maetal.,2014;Kinzneretal.,2019)。水稻是亚洲热带地区广泛种植的粮食作物,危害水稻的害虫常年会受到高温的胁迫,但是水稻害虫,特别是迁飞性种类,是否具有耐热能力提高的遗传与进化,这还不完全清楚。
稻纵卷叶螟Cnaphalocrocismedinalis广泛分布于东亚地区,是我国水稻上的重要迁飞性害虫,在我国华南、华中、华北及云贵高原都有发生记载。稻纵卷叶螟主要以幼虫纵向卷起水稻叶片取食的方式影响水稻叶片叶绿素、光系统Ⅱ活性以及改变剑叶水分,从而造成水稻产量的损失(Padmavathietal.,2013)。21世纪以来,稻纵卷叶螟在我国频频暴发,严重威胁我国水稻生产。温度对稻纵卷叶螟种群影响较大(陈永年等,1981;吴进才和张孝羲,1984)。稻纵卷叶螟成虫期受到高温胁迫后,其繁殖力降低;蛹期经历高温,历期显著缩短,羽化率降低;幼虫经高温处理后存活率显著下降;卵受到极端高温后干瘪,不能孵化(方源松等,2013;廖怀建等,2013;Liaoetal.,2014;Qianetal.,2016)。虽然稻纵卷叶螟受高温的影响较大,但是已有的研究也表明稻纵卷叶螟成虫可通过迁飞和选择产卵场的方式应对高温(吴进才,1985;Bodlahetal.,2016,2017),幼虫可改变叶苞结构和建苞速度、产生热适应性来应对高温(Bodlahetal.,2019;Guetal.,2019)。稻纵卷叶螟具有多种热应对策略,但是幼虫的热适应能力的遗传特性如何还不清楚。因此,本研究以经锻炼后产生的稻纵卷叶螟热锻炼品系为对象,通过杂交试验、耐热能力测定,阐明幼虫热适应的继代效应。研究结果将充实昆虫热适应能力的遗传进化理论,同时为全球变暖形势下依据温度指标进行稻纵卷叶螟发生趋势预测提供指导。
1 材料与方法
1.1 供试虫源
稻纵卷叶螟种群采自江苏南京稻田,并在室内采用小麦苗继代饲养(朱阿秀等,2015)。饲养温度为27±1℃,相对湿度为60%±10%,光周期为14L∶10D。饲养用小麦为常规品种苏麦188。
1.2 幼虫热锻炼
南京7-8月历史最高温为39~40℃,IPPC(2021)预计未来20年全球地表气温将升高1.5℃,因此本研究选用39℃和41℃进行稻纵卷叶螟幼虫的热锻炼。室内27℃饲养的稻纵卷叶螟3龄幼虫连苗带虫转移至27℃光照培养箱中,然后每天上午9:00时开始对培养箱以每30 min升高3℃的方式进行升温操作,分别升到39℃或41℃,保持3 h;然后按每30 min降低3℃的方式,将箱内温度降回至27℃并保持。连续3 d处理后,在27℃条件下饲养。当下一代幼虫发育到3龄时,进行与上一代相同的热锻炼处理。经39℃或41℃连续锻炼30代后,获得热锻炼品系HA39和HA41。两品系3龄幼虫经39℃或41℃每天3 h、连续3 d热击后的存活率与其在27℃条件下无显著差异(Guetal.,2019;Quanetal.,2020)。以一直饲养在27℃下的同样来源种群HA27为对照。每品系幼虫均在2 000头以上。HA39和HA41品系经30代以上热锻炼后用于实验。
1.3 热锻炼品系幼虫的耐热能力测定
将HA27,HA39和HA41三品系的1-5龄幼虫饲养在36℃或41℃条件下,依据各龄期的耐热能力高低测定经历1~144 h不等时长后的存活率,以27℃下HA27的存活率为对照,确定热锻炼品系的耐热能力。剪取30株约25 cm高的小麦苗,用湿润的脱脂棉将其茎基部包裹保鲜,然后置于塑料杯(上口直径8.5 cm,底部直径5.5 cm,高17.5 cm)中,将幼虫接于杯中麦苗上,每杯20头,然后用保鲜膜将杯口密封,并用在膜上戳洞以保持透气。接虫后苗杯转移至36℃或41℃培养箱。每处理重复5次。处理后的前6 h中每小时调查一次存活率,以后每天查一次存活率直到全部死亡或7 d,以相同温度下处理相同时长后的存活率高低表示幼虫的耐热能力大小。
1.4 热锻炼品系与非锻炼品系杂交后的繁殖力及后代幼虫耐热能力测定
将HA27和39℃下经35代热锻炼品系HA39进行正反交和自交,设4种组合:HA27♀×HA27♂,HA39♀×HA39♂,HA27♀×HA39♂和HA39♀×HA27♂。每种组合15对,每对成虫饲养在一个产卵杯中。产卵杯上口径7 cm、下口径5 cm、高10.5 cm,杯底垫有一层约0.5 cm厚的脱脂棉,并滴加0.5%的蜂蜜水湿润,为成虫补充营养,杯口用塑料薄膜封口并戳孔。产卵杯放在温度为27±1℃、相对湿度为60%±10%、光周期为14L∶10D的温室内。每天更换薄膜并统计产卵量,直至雌蛾死亡,计算总产卵量。在每个组合所产卵中随机取50粒,置于27,33和36℃下孵化,计算各配对组合所产卵在不同温度下的孵化率,以明确杂交后代卵的耐热性。重复5次。
将不做孵化率测定的卵连薄膜放于小麦苗上,自然孵化。孵化后幼虫的饲养参照朱阿秀等(2015)的方法。当幼虫发育至3龄时,测定其在36℃和41℃高温下处理3~120 h不等时长后的存活率,以确定杂交后代幼虫的耐热能力。测定方法如1.3节,每处理20头幼虫,重复5次。
1.5 热锻炼停止后幼虫的耐热能力测定
将39℃热锻炼38代后的HA39品系饲养在27℃下,不再进行热锻炼,然后按1.3节的方法测定停止锻炼2代后3龄幼虫在36,39和41℃下暴露1~168 h不同时长后的存活率,并以HA27在同样温度处理相同时长下的存活率为对照。每处理测定20头幼虫,重复5次。
1.6 数据分析
数据统计分析利用SPSS V20.0软件进行。高温处理后各品系间存活率的差异采用ANOVA单因素方差分析,并用Tukey氏法进行平均值间差异的多重比较。幼虫品系与热击时长对存活率的影响采用双因素方差分析。比率数据经反正弦平方根转换后进行方差分析。
2 结果
2.1 热锻炼品系各龄幼虫的耐热能力
2.1.11龄幼虫:热锻炼可提高稻纵卷叶螟1龄幼虫的耐热能力(图1:A,B)。1龄幼虫的耐热能力较差,在36和41℃下处理4 h后全部死亡。在36℃下处理1~3 h时HA41 1龄幼虫的存活率均显著高于HA27 1龄幼虫的(P<0.05),处理1 h时HA39 1龄幼虫的存活率显著高于HA27 1龄幼虫的(P<0.05)(图1:A);在41℃下处理1~3 h时HA41 1龄幼虫的存活率显著高于HA27 1龄幼虫的(P<0.05),处理3 h时HA39 1龄幼虫的存活率显著高于HA27 1龄幼虫的(P<0.05)(图1:B)。
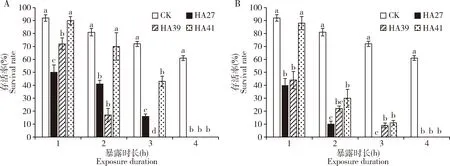
图1 稻纵卷叶螟热锻炼品系HA39和HA41和非锻炼品系HA27的1龄幼虫在36℃(A)和41℃(B)下处理不同时长的存活率Fig.1 Survival rates of the 1st instar larvae of Cnaphalocrocis medinalis from the heat-acclimated strains HA39 and HA41 and the unacclimated strain HA27 exposed to 36℃ (A) and 41℃ (B) for different time.CK:27℃下饲养的HA27品系HA27 strain reared at 27℃;HA39:39℃连续热锻炼30代获得的热锻炼品系Heat-acclimated strain after acclimation to 39℃ for 30 generations;HA41:41℃连续热锻炼30代获得的热锻炼品系Heat-acclimated strain after acclimation to 41℃ for 30 generations.39和41℃热锻炼在3龄幼虫期进行,每天9:00时温度从27℃以3℃/30 min速率逐渐升至39或41℃后保持3 h,然后以同样速率降至27℃并保持,第2和3天以同样方式处理后该世代的热锻炼结束,下一代以同样方式进行。图中数据为平均值±标准误;柱上不同字母表示经Tukey氏HSD检验比较各品系或交配组合在同一暴露时长差异显著(P<0.05)。Heat acclimation to 39℃ and 41℃ was performed at the 3rd instar larval stage.Temperature increased to 39℃ or 41℃ from 27℃ at 9:00 by 3℃ per 30 min and maintained for 3 h,and then decreased to 27℃ with the same rate and maintained till the treatment next day.The same treatment was performed in 3 d.The same heat treatment was carried out in the next generation.Data in the figure are means±SE.Different lowercases above bars mean significant differences between strains or mating combinations for the same exposure duration at P<0.05 using Tukey’s HSD post hoc test.图2-5和图7同The same for Figs.2-5 and Fig.7.
2.1.22龄幼虫:稻纵卷叶螟热锻炼品系2龄幼虫的耐热能力明显提高,在36℃下处理72 h以上稻纵卷叶螟2龄幼虫才全部死亡,并且41℃下处理5 h时仍有幼虫存活(图2:A,B)。热锻炼提高了2龄幼虫对特定高温的耐受力。在36℃下处理1~3 h时HA41 2龄幼虫的存活率与HA27 2龄幼虫的在27℃(CK)下的相同,并且显著高于HA27 2龄幼虫经历热处理1~3 h后的存活率(P<0.05),但经历4 h及以上时长热处理后HA41与HA27 2龄幼虫的存活率差异不显著(P>0.05)。HA39 2龄幼虫在36℃下处理24和48 h的存活率显著高于HA27 2龄幼虫的(P<0.05);36℃下处理72 h时,各品系2龄幼虫的存活率均无显著差异(P>0.05)(图2:A);在41℃下处理1~3 h时各品系2龄幼虫的存活率无显著差异(P>0.05),但处理4 h时HA39和HA41 2龄幼虫的存活率显著高于HA27 2龄幼虫的(P<0.05),处理5 h时HA39 2龄幼虫的存活率显著高于HA27 2龄幼虫的(P<0.05)(图2:B)。
2.1.33龄幼虫:稻纵卷叶螟热锻炼品系3龄幼虫的耐热能力显著增强,在36℃下处理5 d各品系3龄幼虫的存活率仍在50%左右;41℃下处理6 h时3龄幼虫存活率接近50%,但24 h时全部死亡(图3:A,B)。36℃处理2 d时HA41 3龄幼虫的存活率显著高于HA27 3龄幼虫的(P<0.05),其他处理时长下HA39和HA41 3龄幼虫的存活率与HA27 3龄幼虫的相似(图3:A)。在41℃下处理3~5 h时HA41 3龄幼虫的存活率显著高于HA27 3龄幼虫的(P<0.05),但其他处理时长下锻炼品系与非锻炼品系3龄幼虫的存活率差异不显著(P>0.05)(图3:B)。
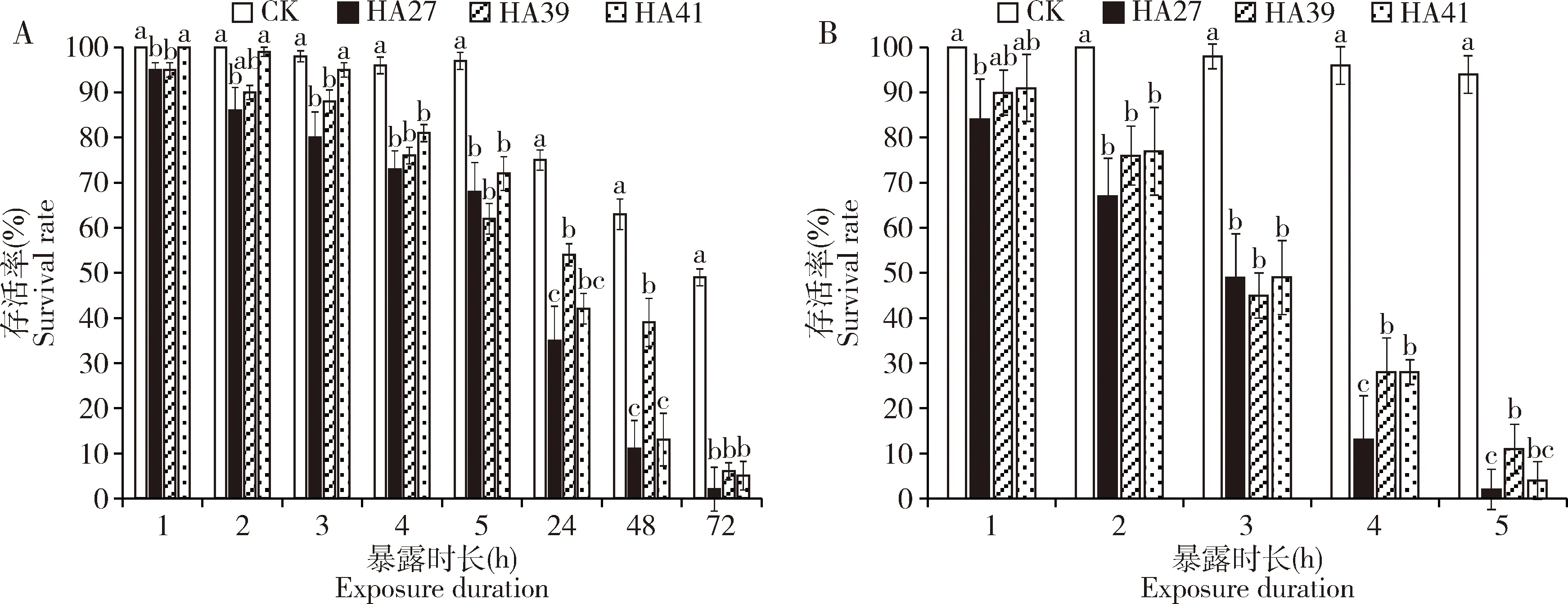
图2 稻纵卷叶螟热锻炼品系HA39和HA41和非锻炼品系HA27的2龄幼虫在36℃(A)和41℃(B)下处理不同时长的存活率Fig.2 Survival rates of the 2nd instar larvae of Cnaphalocrocis medinalis from the heat-acclimated strains HA39 and HA41 and the unacclimated strain HA27 exposed to 36℃ (A) and 41℃ (B) for different time
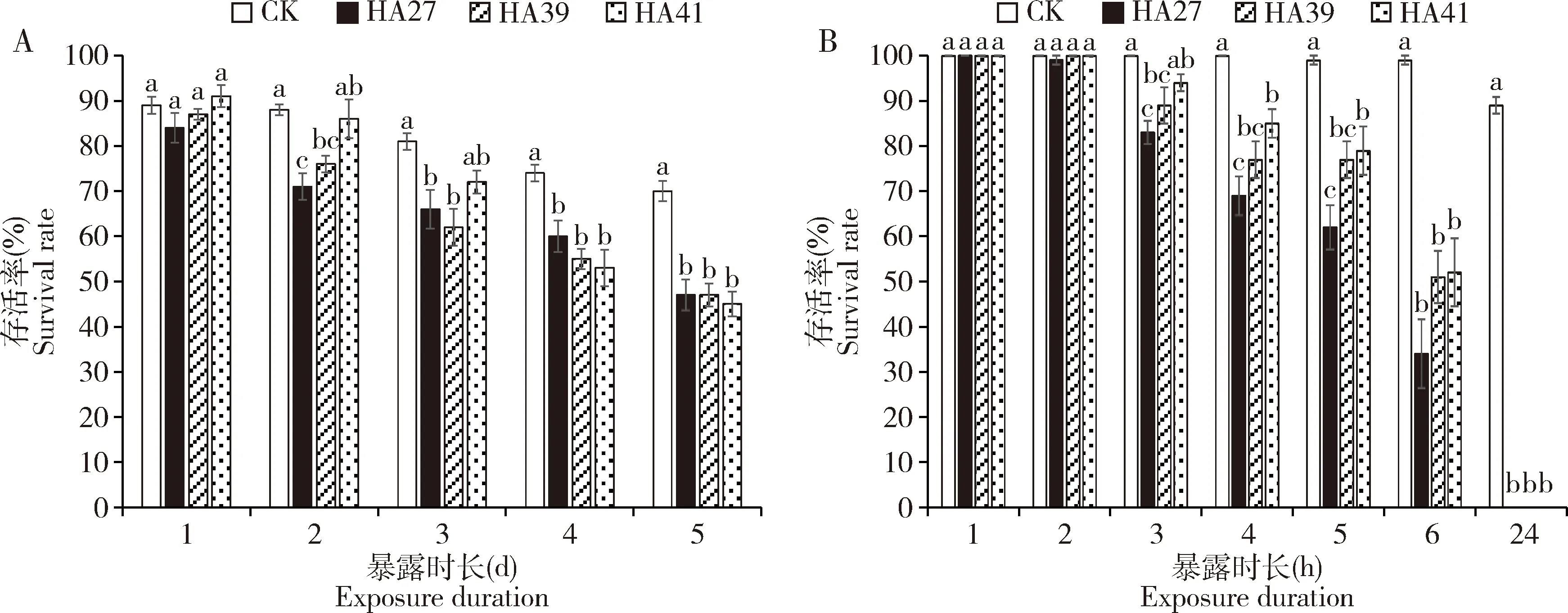
图3 稻纵卷叶螟热锻炼品系HA39和HA41和非锻炼品系HA27的3龄幼虫在36℃(A)和41℃(B)下处理不同时长的存活率Fig.3 Survival rates of the 3rd instar larvae of Cnaphalocrocis medinalis from the heat-acclimated strains HA39 and HA41 and the unacclimated strain HA27 exposed to 36℃ (A) and 41℃ (B) for different time
2.1.44龄幼虫:热锻炼提高了稻纵卷叶螟4龄幼虫的耐热能力。在36℃下处理2和4 d时,HA39的4龄幼虫的存活率显著高于HA27 4龄幼虫的存活率(P<0.05),但HA41 4龄幼虫的存活率与HA27 4龄幼虫的无显著差异(P>0.05),41℃热锻炼不能提高4龄幼虫对36℃高温的耐受性(图4:A);在41℃下处理3~6 h,HA39和HA41 4龄幼虫的存活率均显著高于HA27 4龄幼虫的(P<0.05)(图4:B)。

图4 稻纵卷叶螟热锻炼品系HA39和HA41和非锻炼品系HA27的4龄幼虫在36℃(A)和41℃(B)下处理不同时长的存活率Fig.4 Survival rates of the 4th instar larvae of Cnaphalocrocis medinalis from the heat-acclimated strains HA39 and HA41 and the unacclimated strain HA27 exposed to 36℃ (A) and 41℃ (B) for different time
2.1.55龄幼虫:热锻炼明显提高了稻纵卷叶螟5龄幼虫的耐热能力。在36℃下处理2~5 d HA41 5龄幼虫的存活率显著高于HA27 5龄幼虫的(P<0.05);在36℃下暴露3和4 d时HA39 5龄幼虫的存活率也显著高于HA27 5龄幼虫的(P<0.05)(图5:A)。在41℃下处理5和6 h时HA39 5龄幼虫的存活率显著高于HA27 5龄幼虫的(P<0.05),并且与不受热处理的CK相同,但是HA41 5龄幼虫的存活率与HA27 5龄幼虫的无显著差异(P>0.05)(图5:B)。3龄幼虫期在39℃下的热锻炼可提高其5龄幼虫对41℃高温的耐受性,但在41℃下的锻炼却不能(图5:B)。
2.2 热锻炼与不锻炼品系杂交后代的耐热能力
2.2.1不同交配组合的产卵量和卵孵化率:稻纵卷叶螟热锻炼品系的产卵量和卵孵化率与非锻炼品系的相同,两品系杂交后代的繁殖力与自交后代的一致(图6:A)。HA39与HA27杂交后雌虫产卵量均在200~250粒/雌之间,各交配组合的繁殖力无显著差异(F3,56=1.997,P=0.125)(图6:A)。各交配组合27℃下卵孵化率为97%~99%(图6:B),33℃下为90%~95%(图6:C),各组合间无显著差异(27℃:F3,16=0.705,P=0.563;33℃:F3,16=1.657,P=0.216)。在36℃,所有交配组合所产的卵均不能孵化。

图6 稻纵卷叶螟高温锻炼品系HA39和非锻炼品系HA27各交配组合的产卵量(A)以及在27℃(B)和33℃(C)下的卵孵化率Fig.6 Numbers of eggs laid by different mating combinations of Cnaphalocrocis medinalis between the heat-acclimated strain HA39 and the unacclimated strain HA27 (A),and egg hatching rates at 27℃ (B) and 33℃ (C)图中数据为平均值±标准误;柱上相同小写字母示差异不显著(P>0.05,Turkey氏HSD检验)。Data in the figure are means±SE.The same lowercase letter above bars means no significant difference (P>0.05,Tukey’s HSD test).
2.2.2不同交配组合后代3龄幼虫的耐热能力:稻纵卷叶螟各交配组合后代的3龄幼虫在36℃持续热胁迫1~5 d的存活率没有显著差异(1 d:F3,16=1.385,P=0.284;2 d:F3,16=2.568,P=0.091;3 d:F3,16=1.667,P=0.214;4 d:F3,16=0.920,P=0.454;5 d:F3,16=0.052,P=0.984)(图7:A)。但是在41℃热胁迫5 h时,热锻炼品系及其与非锻炼品系杂交后代的3龄幼虫的存活率显著高于非锻炼品系自交后代的(F3,16=5.742,P=0.007);41℃处理6 h时热锻炼品系雌虫与非锻炼品系雄虫交配所产后代3龄幼虫的存活率也显著高于非锻炼品系自交后代的(F3,16=7.333,P=0.003)(图7:B)。结果说明稻纵卷叶螟幼虫经热锻炼产生的耐热能力可传给后代。
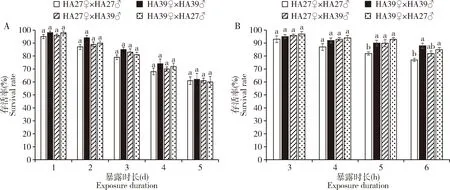
图7 稻纵卷叶螟热锻炼品系HA39与非锻炼品系HA27交配组合后代3龄幼虫在36℃(A)和41℃(B)下处理不同时间的存活率Fig.7 Survival rates of the 3rd instar larvae of Cnaphalocrocis medinalis from the mating combinations between the heat-acclimated strain HA39 and the unacclimated strain HA27 after exposure to 36℃ (A) and 41℃ (B) for different time
2.3 热锻炼停止后幼虫的耐热能力
在36℃和41℃下,经2代恢复的稻纵卷叶螟热锻炼品系(HA39-R2)的存活率与HA27的无显著差异(36℃:F1,56=0.929,P=0.339;41℃:F1,48=0.868,P=0.356)(图8:A,C),但经39℃处理后,HA39-R2幼虫的存活率仍高于非锻炼品系(F1,56=9.058,P=0.004)(图8:B),并且在处理4 h时差异显著(t=2.395,df=8,P=0.044)。由此说明,稻纵卷叶螟幼虫经多世代热锻炼产生的耐热能力在停止锻炼2代后仍能部分保持。
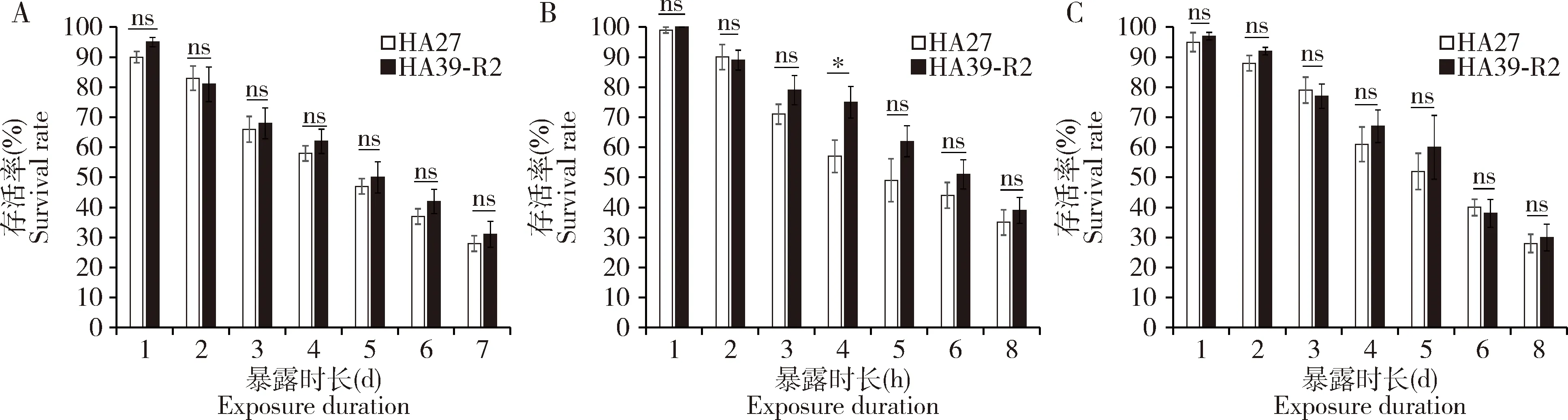
图8 稻纵卷叶螟HA39品系停止高温锻炼2代(HA39-R2)和HA27的3龄幼虫经36℃(A),39℃(B)和41℃(C)处理不同时长的存活率Fig.8 Survival rates of the 3rd instar larvae of Cnaphalocrocis medinalis from HA39 after stopping high temperature acclimation for two generations (HA39-R2) and HA27 when exposed to 36℃ (A),39℃ (B) and 41℃ (C) for different time图中数据为平均值±标准误;柱上星号表示经Student氏t测验差异显著(P<0.05),ns表示差异不显著(P>0.05)。Data in the figure are means±SE.The asterisk above bars in the figure means significant difference (P<0.05),and ns means no significant difference (P>0.05) by Student’s t-test.
3 讨论
昆虫可以通过温度锻炼来提高其应对极端温度胁迫的能力(Angilletta,2009;Colinet and Hoffmann,2012)。赤拟谷盗Triboliumcastaueum成虫、蛹、幼虫和卵经亚致死温度36℃和42℃锻炼后,其对50℃致死高温的敏感性明显降低,高温锻炼显著提高了各虫态在极端温度下的耐受性,且42℃亚致死高温锻炼对赤拟谷盗的耐热性影响更大(Lü ang Liu,2017)。黑腹果蝇Drosophilamelanogaster的25℃和29℃驯化品系在38℃高温条件下的存活率显著高于16.5℃驯化品系(Gilchristetal.,1997)。稻纵卷叶螟每代的3龄幼虫经3 d热处理,39℃驯化5代后即可适应该高温(Guetal.,2019)。在多变环境中,快速进化是多数物种长期存在的重要策略(Hoffmann and Sgrò,2011;Meesteretal.,2018)。多世代的高温锻炼可推动昆虫种群发生适应性进化。在拟果蝇Drosophilasimulans中发现保持有大量的适应性变异,以便于对新环境产生快速进化反应,形成新的适应性表型(Barghietal.,2019)。稻纵卷叶螟的高温锻炼品系与非锻炼品系杂交后代幼虫的耐热能力显著高于非锻炼品系自交后代的(图1~5),表明稻纵卷叶螟幼虫期热锻炼产生的耐热能力能传递给后代,耐热能力具有遗传性。这种能被筛选且有遗传能力的耐热表型很可能是其种群发生快速进化性适应的结果。同时,稻纵卷叶螟热锻炼品系的繁殖力与非锻炼品系的无显著差异(图6)。幼虫通过热锻炼筛选后表现出的高耐热能力不会影响其后续成虫的繁殖力,这种耐热性的产生对种群在高温条件下的发展是有利的。生物经历环境变化获得的适应性可以通过母系传递给子代(Mousseau and Fox,1998)。广聚萤叶甲Ophraellacommuna母代经历低温锻炼后,其子代的耐寒性得到显著提高(Zhouetal.,2013)。稻纵卷叶螟经热锻炼筛选获得的耐热性不仅可通过杂交方式传递给后代,也可通过母代传递给子代,表现为停止热锻炼后的幼虫热适应能力仍能部分保持(图8)。这种热适应性继代效应的存在,意味着其种群在不断经历热胁迫后能逐渐适应高温,全球气候变暖对种群的影响将较小,这很可能是20世纪70年代以来虽然全球变暖在加剧,但该虫暴发成灾更为频繁的原因之一。
全球温度升高对昆虫的影响是系统性的,不仅涉及昆虫,还涉及其寄主植物等。高温影响寄主植物后会间接影响昆虫。水稻幼苗期和抽穗开花期长期受到高温胁迫后,其体内与水稻抗逆性相关的脱落酸含量呈先增加后下降趋势,最终降低了水稻的抗逆性(刘晓龙等,2021)。高温处理显著提高了水稻叶片中N和P的含量(韩笑等,2019)。随着温度从25℃升高到34℃,水稻IR26品种中可溶性糖含量升高、草酸含量下降,对褐飞虱的抗性减弱(王保菊等,2010)。温度影响水稻的抗逆性和叶片中营养物质的含量后,水稻上害虫种群的适合度会发生改变,从而导致害虫种群数量和危害程度的变化。在不同水稻品种上稻纵卷叶螟的生长发育和繁殖力存在差异(王亭等,2018),稻纵卷叶螟成虫对不同水稻品种的选择性不同(常晓丽等,2019)。如果高温改变了水稻的抗虫性,无疑也会导致稻纵卷叶螟种群发生数量的变化。因此,在全球变暖形势下,各水稻品种对稻纵卷叶螟的抗性变化还值得研究。
稻纵卷叶螟热锻炼品系与非锻炼品系在繁殖力上无显著差异,幼虫耐热能力的提高没有繁殖代价,这很可能与本研究中的热锻炼只是在3龄幼虫期进行有关。幼虫期的热胁迫对成虫的繁殖力影响较小(Qianetal.,2017)。热锻炼品系耐热能力的提高是否存在其他方面的代价,如药剂敏感性等,这还需要研究。另外,稻纵卷叶螟幼虫驯化后得到的热适应品系具有遗传性,并且热锻炼与非锻炼品系受热后会引起热激蛋白、表皮蛋白和氧化还原酶活性等相关基因的表达差异(Quanetal.,2020)。稻纵卷叶螟的耐热性有其遗传基础,耐热品系很可能发生了基因型的改变,因此不存在适合度代价,但是热适应相关的基因型或突变还需要继续研究。
猜你喜欢
上海农业科技(2022年5期)2022-10-19
长江蔬菜(2022年11期)2022-07-29
建材发展导向(2021年11期)2021-07-28
当代水产(2020年10期)2020-03-17
园林科技(2020年2期)2020-01-18
当代水产(2019年8期)2019-10-12
上海农业科技(2019年3期)2019-06-25
山东林业科技(2019年2期)2019-06-03
云南农业科技(2019年2期)2019-04-04
浙江农业学报(2017年1期)2017-05-17
