
基于结构优化的择伐对锐齿槲栎天然林结构与生长的影响
2023-01-12 02:37张保全段丰沛郭小龙刘文桢
西北林学院学报 2023年1期
张保全,段丰沛,郭小龙,刘文桢*,张 婷
(1.甘肃省小陇山林业科学研究所 甘肃省次生林培育重点实验室 甘肃小陇山森林生态系统国家定位观测研究站,甘肃 天水 741022; 2.甘肃农业职业技术学院,甘肃 兰州 730030;3.甘肃林业职业技术学院,甘肃 天水 741022)
采伐是森林经营的重要手段,也是对森林的一种十分重要的人为干扰[1]。随着森林经营目标的转变,传统的皆伐作业因对森林生态系统产生显著的负面影响而逐渐被摒弃,而择伐被认为是一种有效的经营方式。研究表明,适度择伐能够改善群落内的光照条件,加快林分更新和生长,对于林分的结构多样性和特殊物种的保护具有积极作用[2-3]。择伐是异龄混交林经营的主要采伐方式,但由于异龄混交林树种组成复杂,林木年龄差异较大,择伐经营方式应用仍存在一些难点,主要表现在应伐木的确定和采伐强度的控制2个方面。应伐木选择的一般原则为“采坏留好、采老留壮、采大留小、采密留均”,但缺乏相应的标准,对于相同的林分类型可能会因为技术工人的认识不同而出现不同的标记结果。国内外针对各种林分类型开展了大量的不同择伐强度对林分结构和生长影响的对比试验,得出的结论也不尽相同[4-9],在培育健康稳定、优质高效的森林生态系统的经营目标下,如何确定应伐木及适当地采伐强度仍然一个重要的问题。
基于结构优化的森林择伐方法是在森林可持续经营原则下提出的经营方法[10-11],该方法在森林经营中采用择伐的方式进行林分结构调整,应伐木选择及采伐强度控制方面与传统择伐方法显著不同,即经营前并不预设采伐强度,在优先考虑林木个体健康的前提下,以林分中的顶极树种或乡土树种的中、大径木为经营对象,运用林分空间结构参数来指导应伐木的选择[10-12],即综合考虑培育对象与最近相邻木组成的结构单元中林木分布格局、树种隔离程度和竞争状态来确定应伐木,具有相应的指标和量化方法。此外,该方法在确定经营方向和经营效果评价时均以林分现状分析为基础[10-11,13],因此在标记采伐后即可预知经营效果,从而可以及时、合理地调整经营措施,避免造成不必要的损失。
秦岭西段的小陇山林区是中国西北地区重要的天然林区,是中国西北地区物种多样性最丰富的地区之一,在水源涵养、保持水土、维护地区生态平衡等方面发挥着不可替代的作用。锐齿槲栎(Quercusalienavar.acuteserrata)天然林是该地区的主要森林类型,在锐齿槲栎天然林的群落结构、种间关系、生物多样性等方面开展了一些研究[15-18],但很少有锐齿槲栎天然林经营效果监测的报道。本研究对甘肃小陇山林区锐齿槲栎天然林内建立的基于结构优化的森林择伐地和未择伐样地3次调查数据进行对比分析,了解基于结构优化的择伐对锐齿槲栎天然林组成结构、径级结构、林分空间结构以及生长的影响,为小陇山林区锐齿槲栎天然林经营提供依据。
1 研究区概况
小陇山林区位于甘肃省东南部(104°22′-106°43′E,33°30′-34°49′N),年均气温7~12 ℃,年积温3 563.9 ℃;年日照时数2 098.7 h。年均降水量460~800 mm,50%~60%的降水集中在7-9月,无霜期185 d,海拔750~3 200 m。兼有我国南北气候特点,生物的地理成分、区系成分复杂多样,是甘肃生物种质资源最丰富的地区之一。小陇山林区海拔2 200 m以下是以锐齿槲栎和蒙古栎(Q.mongolica)为主的天然林,栎类林主要伴生树种有华山松(Pinusarmandii)、漆(Toxicodendronvernicifluum)、冬瓜杨(Populuspurdomii)、白桦(Betulaplatyphylla)、山杨(Populusdavidiana) 、千金榆(Carpinuscordata)、甘肃山楂(Crataeguskansuensis)、刺楸(Kalopanaxseptemlobus)等乔木树种。
2 研究方法
2.1 样地设置与数据调查
样地位于甘肃小陇山林区百花林场墁坪营林区小阳沟营林区71号林班内,小班面积35.6 hm2,海拔1 720 m,坡向东北,平均坡度13°。2008年9月,运用全站仪设置了2块70 m×70 m的全面调查样地,一块为经营样地,一块为对照样地。对样地内胸径大于5 cm的林木全部进行定位、挂牌/编号,并进行胸径测量,记载每株树木的树种、胸径,同时调查林分的郁闭度、坡度、林分平均高、林层数和枯立木情况等;分别在样地的中心、四角设置10 m×10 m的小样方5个,调查更新幼树、幼苗,灌木、草本的种类、数量、高度、盖度等内容。样地每5 a进行1次复测,调查内容与首次调查时完全相同,并对于达到起测胸径(DBH≥5 cm)的林木记录胸径、树高和冠幅等,并测量与其最近3株林木的距离,运用2点间距离公式推算其坐标。样地林分因子特征及复测数据见表1。
2.2 经营样地应伐木确定
在对经营样地的林分状态特征进行分析后,确定了该林分的经营方向,即降低林分密度,提高培育目标的竞争态势和伴生树种的比例,增加林木的混交程度,择伐、利用部分成熟林木。在采伐木确定时,首先将林分中的不健康林木作为采伐对象,然后选择干形通直、生长旺盛的锐齿槲栎、华山松、漆等中、大径木作为培育对象,利用林分空间结构参数,重点调整培育对象的混交和竞争。此次经营共标记45株林木作为采伐对象,其中,有4株林木由于健康原因、8株为达到目标直径而择伐利用,其余33株主要是调整培育对象和避免与相邻木的竞争和混交;采伐木涉及12 个树种,包括锐齿槲栎18株,华山松、白桦、杜梨(Pyrusbetulifolia)、多毛樱桃(Cerasuspolytricha)、湖北海棠(Malushupehensis)、桑(Morusalba)、胡桃楸(Juglansmandshurica)、甘肃山楂和深灰槭(Acercaesium)各1株,春榆(Ulmusdavidianavar.japonica)、茶条槭(Acerginnala)各2株,于2008年12 月进行了采伐作业,采伐株数强度为10.1%,断面积强度为22.6%,属于中度干扰。2013年12月第1次对2块样地进行复测,2018年12月第2次对2块样地进行复测。
2.3 数据分析
2.3.1 林分特征 2块样地的树种组成结构包括林分密度、断面积、平均胸径、树高、郁闭度、树种多样性等内容,树种多样性计算指标包括Shannon-Wiener指数[19]和Pielou均匀度指数[20]。
2.3.2 林分直径结构 将样地中胸径>5 cm的林木按照4 cm径级进行统计,并计算每个径级林木的数量占全部林木总数的比例,绘制林分胸高直径分布图,并用K-S检验方法[21-22]对林分径级结构进行差异显著性检验。
2.3.3 林分空间结构 林分空间结构采用基于4株最近相邻木林分空间关系的结构参数进行分析,主要参数包括角尺度和混交度[23-24],并用K-S检验方法对林分角尺度分布和混交度分布进行差异显著性检验。
2.3.4 林分生长 林木生长采用胸径(胸高断面积)绝对生长率(AGR)和相对生长率(RGR)计算,计算公式如下:
AGR=(dbht-dbh0)/Δt
(1)
RGR=(ln(dbht)-ln(dbh0))/Δt
(2)
式中:dbht和dbh0分别为调查时间t和t0时林木的胸径(胸高断面积),Δt为调查间隔期。采用无重复双因素方差分析方法对不同调查间隔期及处理进行相对生长率差异分析[25]。
林分林木死亡率(m)和进阶率(r)分别计算如下[26]。
m=1-(Nt/N0)1/t
(3)
(4)
式中:Nt和N0分别代表t时和初始调查时(t0)时林分中林木的数量,Ns代表t时的林分中林木存活数量。
3 结果与分析
3.1 择伐对林分树种﹑密度﹑生长及多样性的影响
2008年,对照样地和择伐样地择伐前具有相似的林分因子特征(表1)。择伐样地进行择伐后,林分密度、林木平均胸径、平均树高、平均冠幅及胸高断面积均有所下降,其中,平均胸径和断面积下降幅度较大。2013年和2018年数据表明,择伐样地的林木密度和树种数呈现先下降后增加的趋势,其他因子则呈现持续增加的趋势,其中,2013-2018年林分断面积增长幅度较大。未择伐样地的林木由于自然稀疏,密度呈现下降的趋势;样地内的树种也有所下降,其他林分因子指标均呈现增加的趋势(表1)。
2008年,择伐样地的Shannon-Wiener指数值和Pielou均匀度指数分别为2.595和0.742,择伐后,分别为2.615和0.755;择伐5 a和10 a的Shannon-Wiener指数值和Pielou均匀度指数均有所上升;未择伐样地Shannon-Wiener指数值和Pielou均匀度指数均较择伐样地高,但随着时间的推移树种多样性和均匀性总体上表现出持续下降的趋势。

表1 择伐样地和未择伐样地林分因子Table 1 Stand feature of the management plot and control plot in Xiaoyanggou
3.2 择伐对林分径级结构的影响

图1 2块样地的径级结构Fig.1 Diameter distribution of plots in different survey time
3.3 择伐对林分空间结构的影响
从图2可以看出,择伐样地择伐后,林分中角尺度W=0.5和W=1的结构单元频率明显下降,而W=0.25和W=0.75的结构单元频率有所上升;在2013年和2018年调查时,W=0.25和W=1的结构单元频率有所下降,W=0.75的结构单元频率仍在上升。择伐样地不同的调查时间平均混交度值分别为0.494、0.484、0.479和0.492,始终变动在[0.475,0.517]之间,属于随机分布的范畴。未择伐样地中W=0.5的结构单元频率呈现增加的趋势,W=0.75和W=1的结构单元频率有所下降,而W=0.25结构单元频率呈现先增加后下降的趋势;未择伐样地3次调查的平均角尺度分别为0.534、0.525和0.521,属于轻度团状分布,但总体上有向随机分布转变的趋势,可认为是自然演替的结果,这也符合天然林林木分布的空间格局从聚集到随机的演替规律[27-28]。
择伐样地的混交度分布频率可以看出(图3),择伐后,林分中混交度M=1的结构单元频率有所上升;2013 年与2018年调查表明,林分中M=0的结构单元频率为0,M=0.75的结构单元频率不断下降,而M=1的结构单元频率持续上升。择伐样地不同调查时间的混交度均值分别为0.816、0.831、0.852和0.846,择伐后林分的平均混交度呈现增加的趋势。未择伐样地不同调查时间的林分混交度变化较小。未择伐样地3次调查的平均混交度分别为0.771、0.768和0.768,林分平均混交度呈现略有下降的趋势。择伐样地和未择伐样地不同调查时间的混交度分布差异性不显著。

图2 择伐样地与未择伐样地不同调查时间的角尺度频率分布Fig.2 Uniform angle index distribution frequency of plots in different survey time
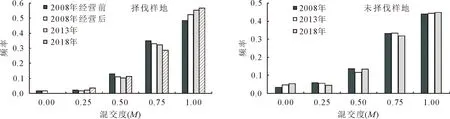
图3 择伐样地与未择伐样地不同调查时间的混交度频率分布Fig.3 Mingling distribution frequency of plots in different survey time
3.4 择伐对林分生长的影响
由表2可以看出,2008-2013年择伐样地和未择伐样地的胸径平均绝对生长率均为0.21 cm·a-1;在2013-2018年择伐样地的胸径平均绝对生长率较未择伐样地的高。在2个调查间隔期内,择伐样地的胸径平均相对生长率分别较未择伐样地高18.4%和55.2%。从断面积平均绝对生长率可以看出,在2008-2013年择伐样地较未择伐样地低,而在2013-2018年择伐样地较未择伐样地高。2个调查期间内,择伐样地的胸高断面积相对生长率分别较未择伐样地高18.2%和54.4%。在2008-2013年择伐样地的进阶率和死亡率分别为3.269%和4.257%,均高于未择伐样地(3.157%和3.703%),而在2013-2018年择伐样地进阶率略有下降,但死亡率下降幅度较大,仅为1.810%;未择伐样地的进阶率有所提高,而死亡率有所下降,分别为2.288%和2.782%。
择伐样地和未择伐样地2次调查间隔期内各径阶的相对生长率可以看出(图4),各径阶的相对生长率均呈现随着径阶的增加而下降。择伐样地2次调查间隔期内,除<9 cm径阶林木的平均相对生长率略有下降外,其他径阶林木的相对生长率均有所提高;未择伐样地<17 cm径阶林木的平均相对生长率略有下降,41~45 cm和51~55 cm略有上升外,其他径阶胸径相对生长率变化很小。对2块样地调查间隔期间的胸径相对生长率进行无重复双因素方差分析结果表明,同一样地内胸径相对平均生长率差异并不显著(表3),但样地内各径阶林木的胸径相对平均生长率差异显著(分别为F=17.031 68≥Fcrit(F临界值)=3.905 204和F=12.185 21>Fcrit=3.241 93)。择伐样地和未择伐样地间在第1个调查间隔期内(2008-2013年),林分整体胸径相对平均生长率差异并不显著,但各径阶林木的胸径相对平均生长率差异显著(F=13.617 44>Fcrit=3.241 93)。在第2个调查期内(2013-2018年),林分整体胸径相对平均生长率差异(F=12.876 7>Fcrit=8.399 74)和各径阶林木的胸径相对平均生长率差异均显著(F=12.400 79>Fcrit=3.241 93),结合表2和图4可以看出,择伐样地在2013-2018年期间的相对生率明显高于未择伐样地。
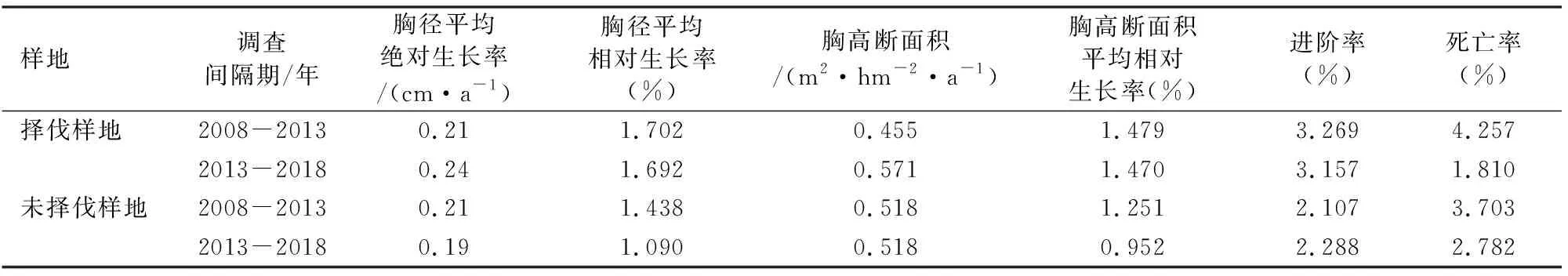
表2 择伐样地与未择伐样地的生长Table 2 The growth comparison of management plot and control plot in different survey intervals
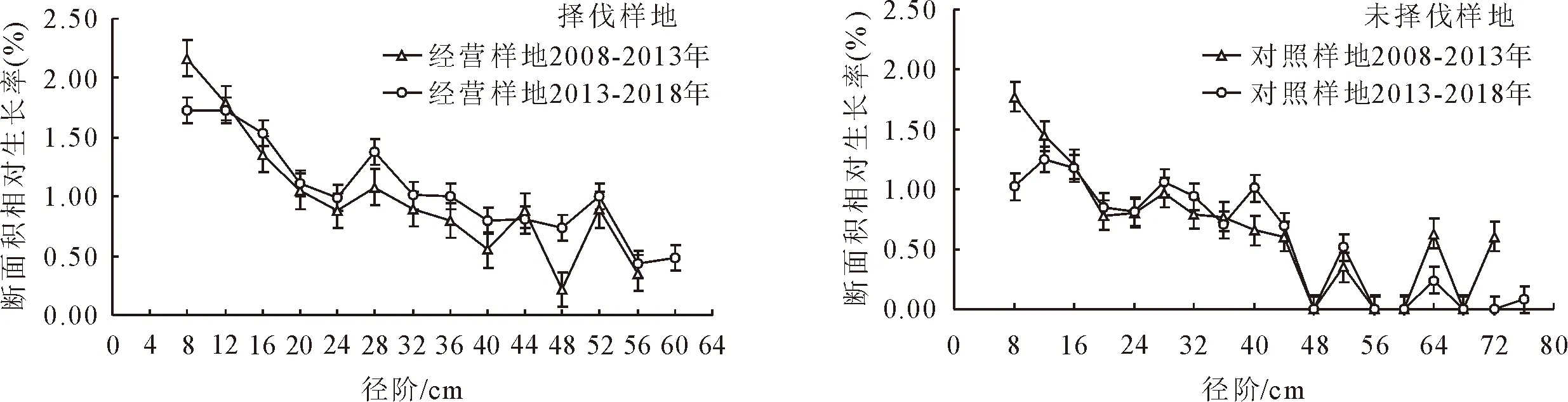
图4 择伐样和未择伐样地各径阶林木相对生长率Fig.4 Relative growth rate of dimeter classes of plots in different survey intervals
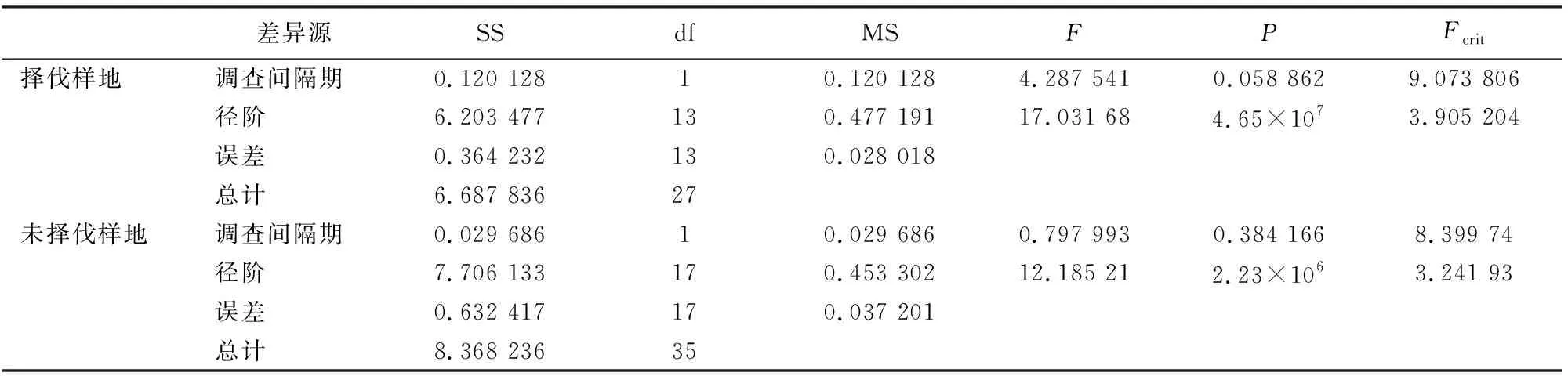
表3 相同样地2次调查间隔期胸径平均相对生长率方差分析Table 3 Variance analysis of average relative growth rate of diameter class in same plot at two survey intervals
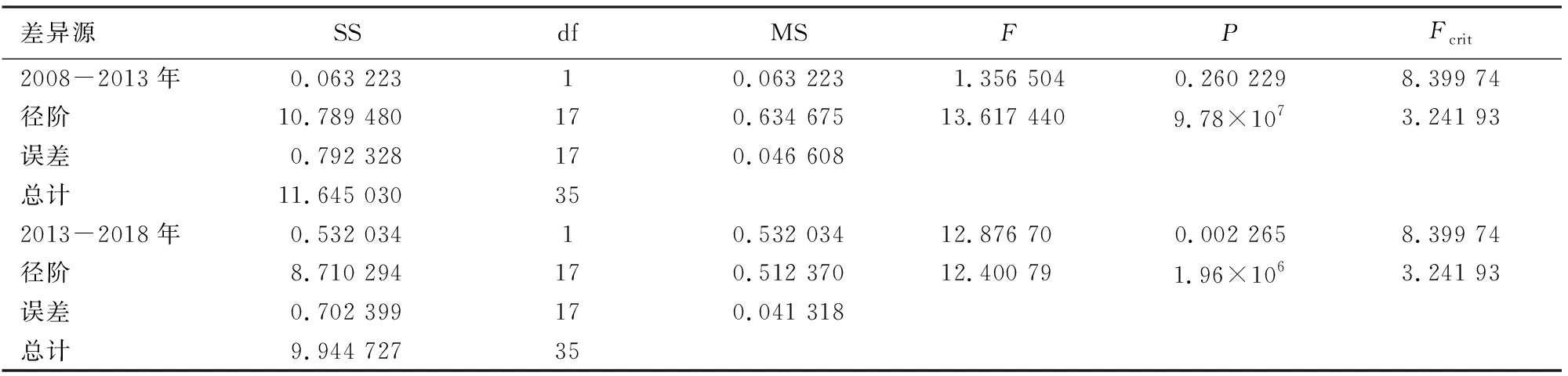
表4 择伐样地与未择伐样地的胸径平均相对生长率方差分析Table 4 Vaiance analysis of average relative growth rate of diameter class of different plot at two survey intervals
4 结论与讨论
4.1 结论
锐齿槲栎天然林采用基于结构优化的择伐方式择伐后,林木的分布格局始终变动在随机分布的范围内,林分的径级结构并未发生明显的改变,仍为典型异龄混交林倒“J”型分布;择伐后林分树种多样性和林木混交程度的持续提高,未择伐样地林分树种多样性和林木混交程度有所下降。
择伐能够促进林木的生长和更新,胸径平均相对生长率和胸高断面积相对生长率分别较未择伐样地提高18.4%~55.2%和18.2%~54.4%。
综上,建议在锐齿槲栎天然林实施管理中应采用基于结构优化的择伐方式,主要由于该择伐方式能够保持天然林林分结构的连续性和稳定性,促进林木生长和更新,利用部分成熟林木。
4.2 讨论
4.2.1 结构优化择伐方式对林分结构的影响 择伐作为重要的人为干扰方式,能够对林分结构产生重要的影响,会因为强度的不同而产生不同的影响,但大多研究仅关注择伐强度对林分径级结构分布的影响,如胡云云等[4]对云、冷杉林不同强度择伐林分研究结果表明,随着初始择伐时间的增加,云、冷杉林的径级结构由反“J”形径级分布向不对称单峰或多峰山状曲线变化,需要控制合理的采伐强度和择伐周期,以保证径级结构不发生明显变化,合理的采伐强度应保持在20%左右,择伐周期为8 a左右。雷相东等[29]研究表明,20%~30%的择伐强度并不能明显改变落叶松、云、冷杉林的多样性和结构。近年来,随着林分空间结构日益受到重视,也有一些研究开始关注择伐对林分空间结构的影响。陈辉荣等[30]对择伐后恢复的杉木、马尾松和阔叶混交林空间结构进行了分析,发现择伐对林木的分布格局影响不显著,弱度和中度择伐可以增加林分的混交度,而强度择伐会降低林分的混交度。吴蒙蒙等[31]对带岭林业实验局东方红林场的红松阔叶林不同强度择伐后林分空间结构变化进行了分析,得出了类似的结论,但这些择伐试验在进行采伐木选择时并没有考虑林分的空间结构。与传统研究相比,本试验是对锐齿槲栎天然林择伐样地林分状态特征进行分析后,确定了择伐方向,用林分空间结构参数作为指导,针对锐齿槲栎、华山松、漆树等主要树种的竞争态势、混交程度进行结构调整,并采伐留用了部分成熟林木。择伐样地和未择伐样地的径级结构和林分空间结构多次调查数据对比表明,基于结构优化的择伐并未显著改变林分的径级结构和林木的分布格局,尽管择伐样地的径级宽幅减小,这也体现了择伐利用部分成熟林木的择伐目的;此外,择伐样地林分的树种多样性和林分混交度呈现增加的趋势,尽管这一变化并不十分显著,但在一定程度上说明,择伐促进了林分树种多样性的提高和树种分配的均匀性,增加了林木的混交程度。
4.2.2 结构优化择伐方式对林分生长的影响 适度的择伐会促进林分的生长,但仅根据林分生长和蓄积量的增长来确定择伐强度的方法存在一定的局限性[32],因为这一方法始终是模糊且受到多种标准影响[33],特别对于具体林分而言,不同的林分会因为面临问题不同而选择不同的采伐强度。本研究在择伐前并未设置采伐强度,而是根据择伐目标选择采伐后得到了采伐强度为22.6%,也属于中度干扰。以上结果虽然无法直接排除由于择伐强度适当而促进了林分生长和多样性的提高,但从林分的结构变化来看,基于结构优化的择伐方法可以保持林分的连续性和稳定性。择伐对林分的影响涉及许多方面,本研究仅对采用结构优化的方法择伐后林分整体的径级结构、林木分布格局、多样性及径向生长等方面的影响进行了分析,今后的研究还需要进一步对林分中主要种群的动态变化、培育目标林木的生长进行系统的评价和长期的监测。
猜你喜欢
广西林业科学(2022年4期)2022-11-07
林业调查规划(2022年3期)2022-06-13
防护林科技(2020年9期)2020-11-09
防护林科技(2020年6期)2020-08-12
绿色科技(2019年6期)2019-04-12
绿色科技(2019年6期)2019-04-12
中南林业调查规划(2018年3期)2018-11-24
中南林业调查规划(2015年3期)2015-12-20
西北植物学报(2015年9期)2015-07-05
中南林业科技大学学报(2015年8期)2015-05-22
