
根区温度胁迫对番茄幼苗根系生长及蔗糖代谢的影响
2023-01-31 03:13刘冰珠孟宪敏张梦夏董春娟尚庆茂
中国蔬菜 2023年1期
刘冰珠 张 锋 雷 蕾 孟宪敏 张梦夏 董春娟 尚庆茂
(中国农业科学院蔬菜花卉研究所,农业农村部园艺作物生物学与种质创制重点实验室,北京 100081)
番茄(Solanum lycopersicum)是重要的蔬菜作物,以育苗移栽为主。培育壮苗是提高番茄产量的先决条件(Dezhabad &Haghighi,2020)。强大的根系是壮苗的重要标志(Nagel et al.,2009)。根系是吸收水分和矿质养分的主要器官,其感受并传递基质/土壤中各种胁迫信号,并且通过改变自身的形态来应对各种逆境胁迫(Malamy,2005;Lamers et al.,2020)。根区温度胁迫已成为制约番茄幼苗生长的主要逆境(傅国海 等,2016;张耀丰,2019;Dezhabad &Haghighi,2020)。探究番茄幼苗对温度胁迫的响应机制有助于提高其抗逆性。
糖及糖代谢在植物响应逆境胁迫中起着重要的作 用(Bhattacharya &Kundu,2020;Yoon et al.,2021;陈宏艳 等,2022)。在胁迫条件下,黄瓜(Du et al.,2010)和番茄(Zhou et al.,2011)根系中糖酵解酶活性上调,促进能量代谢,产生更多能量来适应胁迫。可溶性糖可作为渗透调节物来提高植株抗逆性,常作为判断品种抗逆性强弱的筛选指标。糖还可以作为信号分子在逆境胁迫中起作用。糖信号转导有4 条途径,其中己糖激酶(HXK)信号转导途径中的HXK 和磷酸戊糖(PPP)途径中的关键酶葡萄糖-6-磷酸脱氢酶(G-6-PDH)在响应低温胁迫、干旱胁迫和盐胁迫中均有积极作用(Yang et al.,2014;Li et al.,2017);糖酵解(EMP)信号转导途径在响应逆境胁迫有间接调控作用(Melvin et al.,2017)。
根区温度胁迫对番茄幼苗地上部的影响已有研究报道(张耀丰,2019;Hmiz &Ithbayyib,2021),但对根系的研究鲜有报道。本试验以耐高低温品种久绿787 和不耐高低温品种硬粉8 号为试材,研究根区10 ℃低温和34 ℃高温对番茄幼苗根系生长及蔗糖代谢的影响,以期为培育番茄壮苗提供理论依据。
1 材料与方法
1.1 材料培养
供试番茄材料为耐高低温品种久绿787,购于北京育正泰种子有限公司;不耐高低温品种硬粉8号,由北京市农林科学院蔬菜研究中心提供。
试验于2021 年11 月至2022 年1 月在中国农业科学院蔬菜花卉研究所南区玻璃温室内进行。育苗基质为蛭石,粒径2~4 mm。挑选籽粒饱满的种子于室温下浸种6 h 后,使用5% NaClO 消毒10 min,用蒸馏水反复冲洗,然后在28 ℃条件下恒温催芽。选择发芽一致的种子,播于装有蛭石的纸杯(高13.7 cm,上口径9 cm,底部直径5.8 cm,底部打孔)中,覆盖1.5 cm 厚的蛭石,置于玻璃温室中培养。
1.2 试验设计
于播种后30 d,进行根区10 ℃低温和34 ℃高温处理,以温室中正常栽培的幼苗作为对照(根区平均温度为20 ℃),共胁迫处理6 d;每处理每个品种30 株,3 次重复。根区温度胁迫通过图1 所示的装置实现,挑选生长一致的幼苗,用塑料包裹纸杯置于固定架上,冷暖机设定温度,通过连接水泵,有效控制并保持水槽中水的温度,同时通过隔热棉的设置,增强了水槽的保温效果,减少水槽中温度和水分的散失。利用冷暖机将水槽水温控制在10 ℃实现根区低温胁迫,将水槽水温控制在34 ℃实现根区高温胁迫。根区温度测定方式:将温室娃娃(型号GIS-4-CE-3V)测定地温的探针插入基质内,每隔5 min 记录1 次根区温度,纸杯中基质温度变化范围可以控制在±1 ℃以内,能够满足试验要求(图2)。
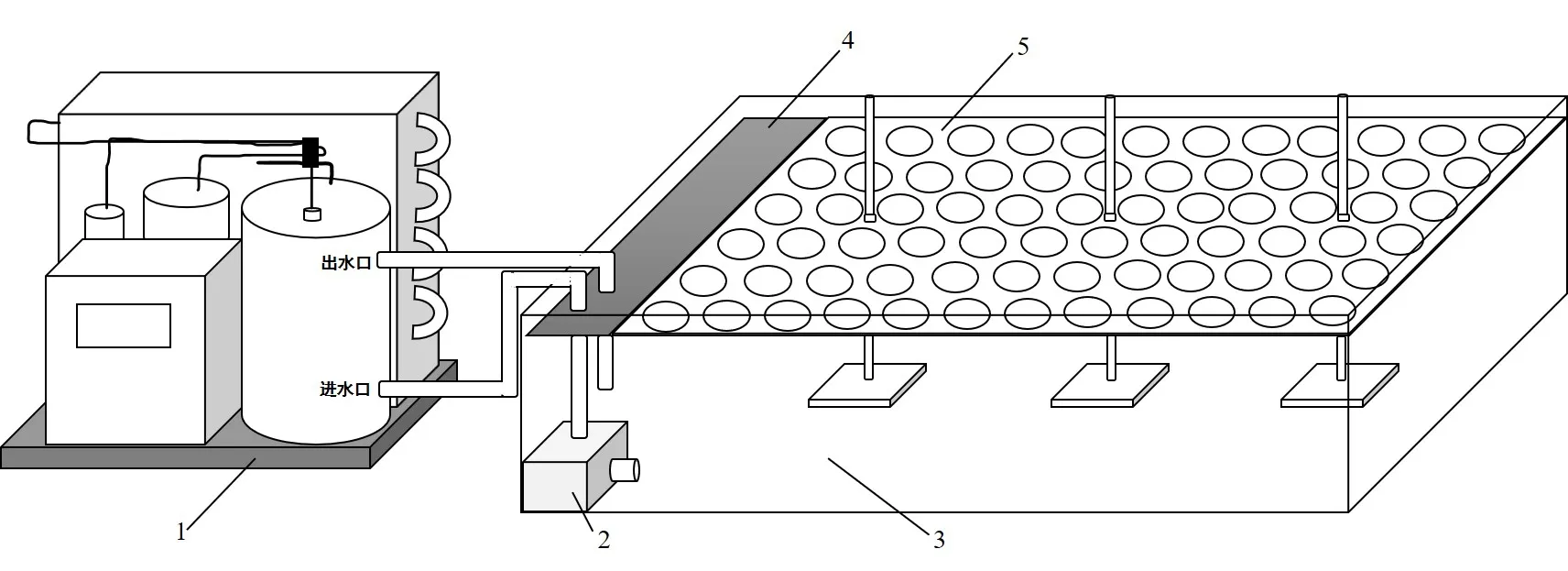
图1 控温装置
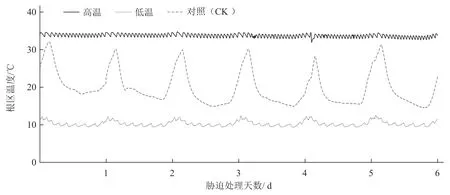
图2 各处理番茄幼苗根区温度变化
1.3 测定指标与方法
1.3.1 根系形态指标测定 胁迫第6 天取样,每处理4 株。将番茄幼苗根系用清水清洗干净,用剪刀在茎基部剪断,将完整根系置于根系扫描仪(LA-S型,杭州万深检测科技有限公司)中扫描拍照观察,并测定根系总长度、根系表面积、根系体积及不同径级根长和表面积等形态学参数。
1.3.2 根系干鲜质量测定 胁迫第3、6 天取样,每处理3 株。将番茄幼苗根系用清水清洗干净,用剪刀在茎基部剪断,用吸水纸将根系擦干,用电子天平称量根系鲜质量;然后放入烘箱中,105 ℃下杀青15 min 后,80 ℃烘干至恒重,用电子天平称量根系干质量。
1.3.3 根系中糖含量及蔗糖代谢相关酶活性测定 胁迫第0、3、6 天取样,每处理2 株。番茄幼苗根系用清水清洗,水温为各自胁迫温度和室温,洗净后用剪刀在茎基部剪断,用吸水纸擦干根系后放入液氮中,存于-80 ℃冰箱保存。葡萄糖、果糖和蔗糖含量采用离子色谱仪(ICS-3000,Dionex,美国)测定(Dong et al.,2011)。蔗糖磷酸合成酶(SPS)、蔗糖合成酶(SS)分解方向(SS-I)、细胞壁转化酶(CWIN)、可溶性酸性转化酶(SAI)和中性转化酶(NI)活性测定分别参照酶活性测定试剂盒(北京索莱宝科技有限公司)说明书进行。
1.4 数据处理
利用Microsoft Excel 2019 和Origin 2018 软件处理数据和作图,利用SPPS 26 软件进行差异显著性分析。
2 结果与分析
2.1 根区温度胁迫对番茄幼苗根系表型的影响
如图3 所示,与对照(CK)相比,根区10 ℃低温和34 ℃高温对久绿787 和硬粉8 号根系生长均产生了不同程度的抑制,其中硬粉8 号受抑制程度大于久绿787。根区10 ℃低温对2 个番茄品种幼苗根系总长度、表面积和体积均产生了显著抑制,胁迫第6 天久绿787 根系总长度、表面积和体积分别较对照降低了31.1%、28.1%、26.5%,硬粉8 号较对照分别降低了40.0%、36.6%、37.8%(图4)。

图3 根区温度胁迫6 d 后番茄幼苗根系形态观测结果
根区34 ℃高温使2 个品种幼苗根系总长度、表面积和体积较对照均明显降低,胁迫第6 天久绿787 根系总长度和表面积均显著低于对照,较对照分别降低了23.2%、23.6%,根系体积较对照降低了22.2%,但未与对照有显著差异;而硬粉8 号根系总长度、表面积和体积均显著低于对照,较对照分别降低了30.3%、29.2%和29.0%(图4)。
2.2 根区温度胁迫对番茄幼苗不同径级根系形态的影响
如图5 所示,2 个番茄品种幼苗根系长度占比最大的径级范围基本一致,主要集中在0~0.5 mm,且此区间根系长度占总长度的比例相差不大,均达93.0%~95.0%。随着径级的增大,根系长度占比逐渐减少。根区10 ℃低温胁迫使久绿787 在径级0~0.2 mm 和0.2~0.5 mm 中的根系长度较对照分别降低了37.6%、24.4%,硬粉8 号较对照分别降低了50.7%、29.7%。根区34 ℃高温胁迫使久绿787 在径级0~0.2 mm 和0.2~0.5 mm 中的根系长度较对照分别降低了20.6%、26.3%,硬粉8 号较对照分别降低了33.1%、27.9%。对照、根区10℃低温和根区34 ℃高温处理的久绿787细根(≤0.5 mm)总长度分别是相应处理硬粉8 号的1.52、1.75、1.68 倍。表明,久绿787 细根根长在根区温度胁迫中受抑制程度低于硬粉8 号。
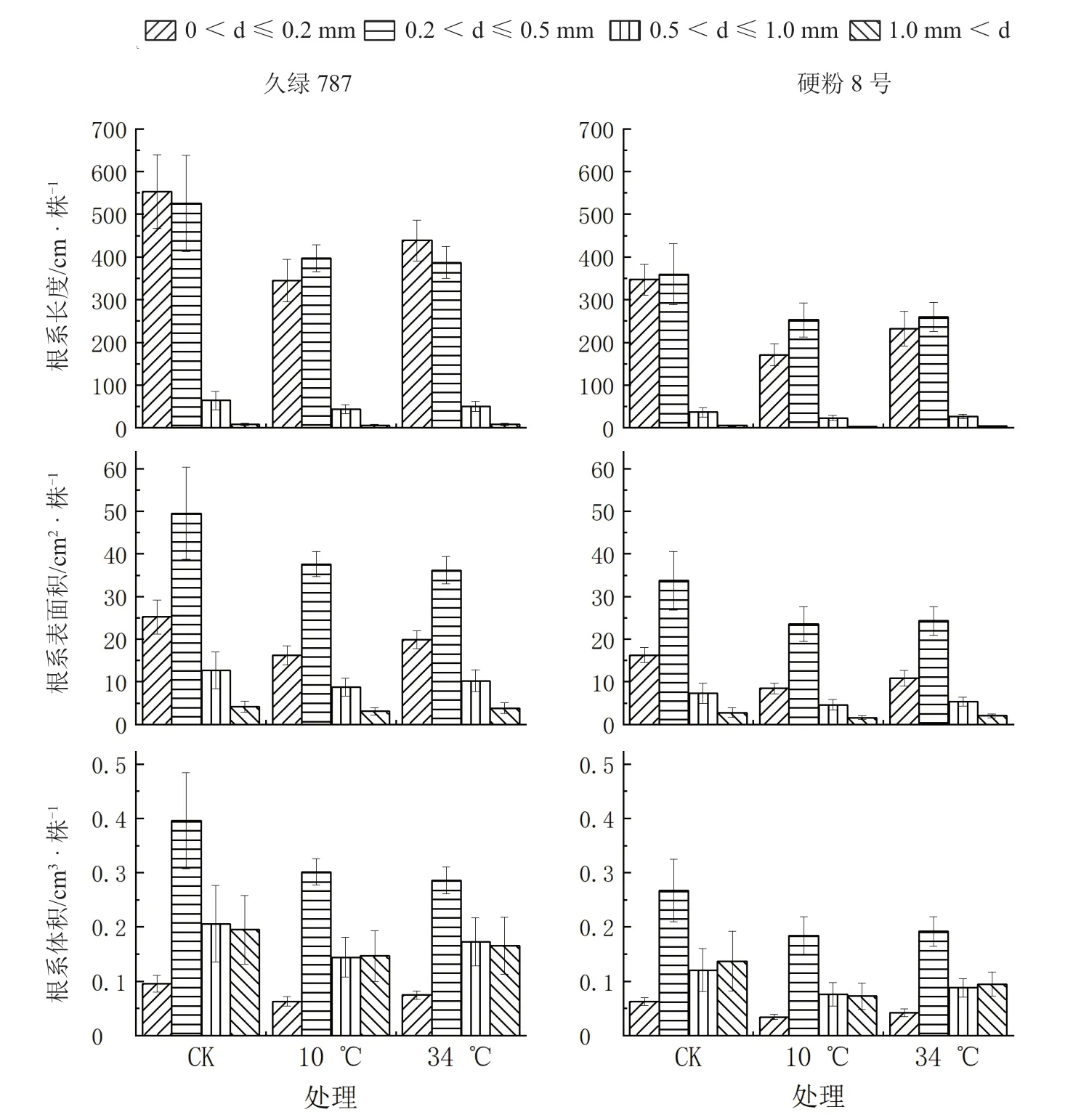
图5 根区温度胁迫后番茄幼苗根系长度、表面积和体积在不同径级中的变化
2 个品种幼苗根系表面积在径级0.2~0.5 mm的占比最大,其次是径级0~0.2 mm,表明径级为0~0.5 mm 的根系是番茄幼苗吸收水分与矿物营养的主体。根区10 ℃低温处理的久绿787 在径级0.2~0.5 mm 的根系表面积较对照降低了24.0%,硬粉8 号较对照降低了30.2%。根区34 ℃高温处理的久绿787 在径级0.2~0.5 mm 的根系表面积较对照降低了27.0%,硬粉8 号较对照降低了27.9%。对照、根区10 ℃低温和根区34 ℃高温处理的久绿787 细根表面积分别是相应处理硬粉8 号的1.50、1.68、1.59 倍。表明,久绿787 细根吸收面积在根区温度胁迫中受抑制程度低于硬粉8 号。
2 个品种幼苗根系在径级0.2~0.5 mm 的体积占总体积的比例最大,其次是径级0.5~1.0 mm 和>1.0 mm,占比最小的为径级0~0.2 mm。径级>0.5 mm 的根系长度和表面积占比远小于径级0~0.2 mm 的占比,但根系体积占比明显高于0~0.2 mm径级占比,表明径级>0.5 mm 的根系主要起运输和支持作用。对照、根区10 ℃低温和根区34 ℃高温处理的久绿787 细根(≤0.5 mm)和粗根(>0.5 mm)根系体积分别是相应处理硬粉8 号的1.49、1.67、1.54 倍和1.55、1.96、1.85 倍。
2.3 根区温度胁迫对番茄幼苗根系生物积累量的影响
如表1 所示,除久绿787 的根系干质量外,根区10 ℃低温处理均使2 个品种幼苗根系鲜、干质量受到不同程度抑制。根区10 ℃低温处理的久绿787 和硬粉8 号根系鲜、干质量在胁迫第3 天与对照相比均无显著差异;胁迫第6 天,久绿787 根系干质量显著低于对照,较对照降低了17.8%,而硬粉8 号根系鲜、干质量均受到显著抑制,较对照分别降低了22.8%和41.4%。
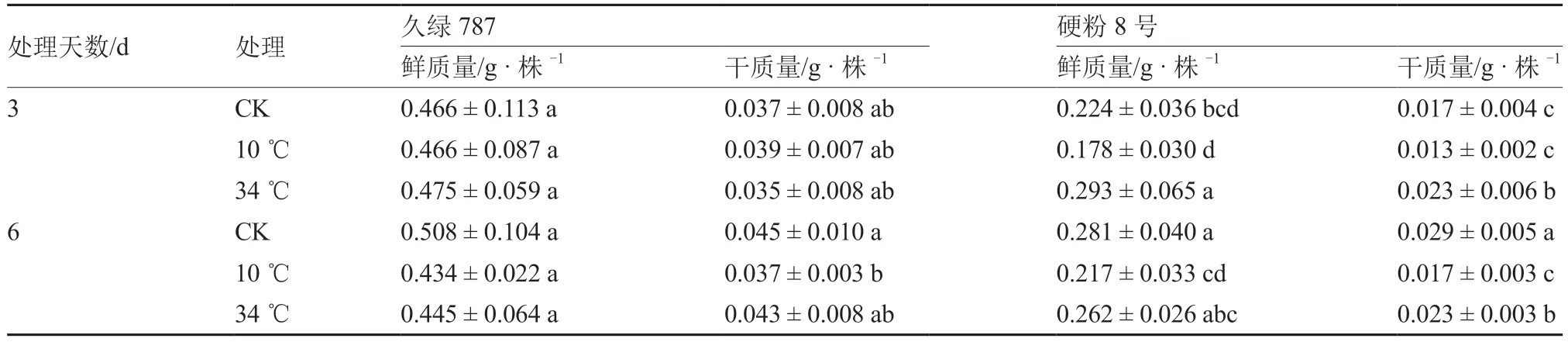
表1 根区温度胁迫对番茄幼苗根系鲜、干质量的影响
根区34 ℃高温处理对久绿787 根系鲜、干质量的抑制程度较小,均与对照显著不差异;硬粉8号根系鲜、干质量在胁迫第3 天时显著高于对照,而胁迫第6 天时均低于对照,其中根系干质量显著低于对照,较对照降低了20.7%。
2.4 根区温度胁迫对番茄幼苗根系中葡萄糖、果糖和蔗糖含量的影响
如图6 所示,根区10 ℃低温处理第3、6 天时,2 个番茄品种幼苗根系中葡萄糖、果糖和蔗糖含量均显著高于对照。其中,久绿787 根系中葡萄糖、果糖和蔗糖含量在胁迫第3 天较对照分别增加了30.8%、39.8%、77.0%,胁迫第6 天较对照分别增加了170.4%、148.4%、56.2%;硬粉8 号胁迫第3 天较对照分别增加了58.3%、133.6%、147.6%,胁迫第6 天较对照分别增加了232.1%、272.9%、165.7%。虽然久绿787 根系中葡萄糖、果糖和蔗糖含量的增长率低于硬粉8 号,但其含量高于硬粉8 号。
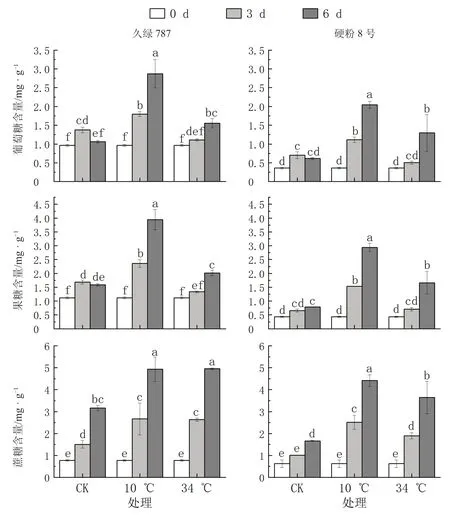
图6 根区温度胁迫对番茄幼苗根系中葡萄糖、果糖和蔗糖含量的影响
与对照相比,根区34 ℃高温胁迫下久绿787和硬粉8 号根系中葡萄糖和果糖含量均呈先降低后增加的变化趋势,且在胁迫第6 天时显著高于对照;而蔗糖含量均显著增加。胁迫第6 天,久绿787 根系中葡萄糖、果糖和蔗糖含量较对照分别增加了46.5%、27.1%和56.9%,硬粉8 号较对照分别增加了110.3%、110.5%和119.3%。虽然久绿787 根系中葡萄糖、果糖和蔗糖含量的增长率低于硬粉8号,但其含量仍高于硬粉8 号。
2.5 根区温度胁迫对番茄幼苗根系蔗糖代谢相关酶活性的影响
由图7 可知,根区10 ℃低温胁迫使久绿787根系蔗糖代谢相关酶活性随胁迫时间延长呈逐渐上升的趋势,而硬粉8 号酶活性变化趋势各不相同。根区10 ℃低温处理的久绿787 在胁迫第3 天SPS 活性显著高于对照,SS-I 活性显著低于对照,CWIN、SAI 和NI 活性与对照均无显著差异;胁迫第6 天,SPS、SAI 和NI 活性均显著高于对照,SS-I 和CWIN 活性与对照无显著差异。根区10 ℃低温处理的硬粉8 号在胁迫第3 天除NI 活性受到显著抑制外,其余酶活性与对照差异均不显著;胁迫第6 天,SPS 和CWIN 活性均显著低于对照,SS-I 和NI 活性与对照均无显著差异,SAI 活性显著高于对照。表明,久绿787 在根区低温胁迫中根系蔗糖代谢相关酶活性受抑制程度要低于硬粉8号,且低温胁迫下蔗糖代谢中积极响应的酶为SPS和SAI。
根区34 ℃高温胁迫使2 个品种幼苗根系蔗糖代谢中分解酶(SS-I、CWIN、SAI 和NI)活性均显著低于对照。根区34 ℃高温处理的久绿787 在胁迫第6 天SPS 活性高于对照,但未达显著水平;硬粉8 号SPS 活性与对照相比呈先增加后降低的变化趋势,在胁迫第3 天时显著高于对照,胁迫第6天时显著低于对照。
3 结论与讨论
根区温度胁迫是影响植物生长的重要因素(Tahir et al.,2010),使幼苗株高、茎粗、叶面积以及生物积累量等受到抑制,叶片净光合速率、气孔导度和蒸腾速率等下降,进而影响光合作用(张耀丰,2019;陈连珠 等,2020;Hmiz &Ithbayyib,2021)。张耀丰(2019)研究表明,番茄幼苗在根区低温胁迫下根系长度、表面积、体积等表型指标均受到显著抑制。李润儒(2015)研究发现,根区35 ℃高温对叶用莴苣(生菜)根系生长产生显著抑制。本试验结果表明,番茄幼苗在根区10 ℃低温和34 ℃高温胁迫下根系总长度、表面积和体积均受到明显抑制,久绿787 根系表型指标受抑制程度要小于硬粉8 号,与陈连珠等(2020)研究根区35.8 ℃高温对耐热与不耐热苗用型大白菜根系的影响结果相似。根区温度胁迫对久绿787根系生物积累量的显著抑制只发生在10 ℃低温胁迫下,而硬粉8 号在10 ℃低温和34 ℃高温胁迫下均有发生。
根系长度和表面积是衡量根系吸收水分和矿物营养能力的重要因子(孙三杰 等,2012)。不同径级的根系吸收能力不同,径级较小的细根吸收能力最强(Sullivan et al.,2000)。根区温度胁迫使久绿787 细根(≤0.5 mm)根长和表面积均受到抑制,但受抑程度明显低于硬粉8 号,表明胁迫处理后的久绿787 根系吸收水分和矿质营养能力要强于硬粉8 号。粗根(>0.5 mm)虽然在根长和根系表面积中的占比较小,但在根系体积中的占比高于径级为0~0.2 mm 的细根,表明粗根起着重要的运输和支持作用。
单羽等(2021)研究表明,可溶性糖在提高植物抗性中的作用很大,一部分用于自身代谢所需的能量物质;一部分用于积累含量,作为渗透调节物,提高抗性。本试验中,根区温度胁迫第6 天时2 个番茄品种幼苗根系中葡萄糖、果糖和蔗糖含量均增加,可提高其抗性,但糖含量变化在10 ℃低温胁迫和34 ℃高温胁迫下有差别,根区10 ℃低温处理的2 个品种幼苗根系中葡萄糖、果糖和蔗糖含量均显著高于对照,起到抗逆作用,与张耀丰(2019)研究根区低温胁迫叶片糖含量结果相似。根区34 ℃高温处理主要促进了根系中蔗糖的积累,与短期35 ℃高温胁迫(张洁 等,2006)和昼夜35 ℃高温胁迫(郭金妹 等,2007)对番茄幼苗叶片中糖含量的影响结果相似。根区10 ℃低温和34 ℃高温处理的久绿787 根系中葡萄糖、果糖和蔗糖含量均高于硬粉8 号,但增长率明显低于硬粉8 号,这可能与久绿787 细根(≤0.5 mm)生长受抑制程度低于硬粉8 号,其需要靠分解糖来提供能量和其他物质,而硬粉8 号根系中可溶性糖多以渗透调节物起抗逆作用有关。
蔗糖代谢相关酶主要包括催化合成蔗糖的蔗糖磷酸合成酶(SPS);分解蔗糖的蔗糖合成酶(SS)分解方向(SS-I)和转化酶(INV),INV 又可分为细胞壁转化酶(CWIN)、可溶性酸性转化酶(SAI)和中性转化酶(NI)。本试验中,NI 活性比其他分解酶活性低很多,与程智慧等(2007)研究结果一致。根区温度胁迫对番茄幼苗根系蔗糖代谢的影响较为复杂。根区10 ℃低温和34 ℃高温处理的久绿787 根系中SPS 活性均有明显升高的趋势,表明SPS 在响应温度胁迫中起积极作用,而分解酶(SS-I、CWIN、SAI 和NI)活性表现截然不同,根区10 ℃低温处理的根系中分解酶活性显著高于根区34 ℃高温处理。久绿787 和硬粉8 号在根区10 ℃低温处理下分解酶活性变化各不同,与华利静(2011)的研究结果相似。INV 在响应低温胁迫中有重要作用(Ruan et al.,2010;Ruan,2014),本试验中,根区10 ℃低温处理第6 天时2 个番茄品种根系中SAI 活性均显著高于对照,表明转化酶中SAI 主要响应低温胁迫。高温胁迫抑制SS-I 和INV 的活性(张洁 等 2006;郭金妹 等,2007;陈宏艳 等,2022),与本试验中根区34 ℃高温处理对2 个品种根系中分解酶(SS-I、CWIN、SAI 和NI)的活性均产生显著抑制的结果一致。
猜你喜欢
中国农业大学学报(2022年11期)2022-11-07
广西林业科学(2022年4期)2022-11-07
农业现代化研究(2022年3期)2022-06-29
林业调查规划(2022年3期)2022-06-13
广西糖业(2020年3期)2020-09-25
浙江工业大学学报(2017年5期)2018-01-22
中国生态农业学报(中英文)(2017年2期)2017-02-16
中国糖料(2016年1期)2016-12-01
西北植物学报(2015年9期)2015-07-05
中南林业科技大学学报(2015年8期)2015-05-22
