
杉木高世代遗传参数估算及高世代育种材料选择技术
2023-01-31 03:00张能军邱勇斌凌高潮郑文华余雪琴
浙江林业科技 2023年1期
张能军,邱勇斌,凌高潮,郑文华,余雪琴
(开化县林场,浙江 开化 324300)
研究材料的评价和选择贯穿于林木育种的各个环节,是林木遗传改良的关键。选择育种通常是以表型值为基础,因含有环境效应,所以选择效率和准确性都较差,加之林木育种试验占地面积大、周期长,受人为干扰和外界条件的影响大,这必然会增大评价试验材料的难度,更不容易准确地选择出遗传品质真正优良的改良材料[1-2]。为克服以上缺点,国外自20 世纪80 年代中期以来,开始大量地应用育种值来评价育种材料,目前已报道的树种有白花泡桐Paulownia fortunei、湿地松Pinus elliottii、火炬松P.taeda、日本落叶松Larix kaempferi等[1-4],揭示了林木主要经济性状的遗传变异规律和遗传参数,提高了林木的遗传增益水平,为选育优良亲本和筛选高代种质提供理论基础。
杉木Cunninghamia lanceolata是我国南方最主要的用材树种之一,其遗传改良工作在国内林木遗传改良中居前列,经过几十年的遗传改良,杉木已进入到高世代改良阶段[5-7]。目前,浙江省的杉木3 代初级种子园已经投产,并开展了3 代种子园的自由授粉和控制授粉家系的子代试验林测定工作,采用育种值评选了杉木一代幼林优良家系和前向选择2 代育种亲本[8],这为研究杉木3 代育种材料的评选积累了经验。但是,对杉木高世代育种的遗传参数和亲本育种值估算研究鲜见报道。如何精确地估计林木高世代改良性状的遗传参数和育种值,提高逆向选择优良家系和前向选择高世代育种亲本效率,是当前亟需解决的问题。本研究利用GeneXpro Tools 4.0 软件,挖掘木材密度与胸径间的关系式,获得全林木材密度值和其转化分析法的结果,基于最佳线性预测(Best linear prediction,BLP)处理杉木二代半同胞家系非平衡试验数据,获得精确的调节系数,有效地估计遗传参数,可减少预测育种值的误差方差,预测不同世代个体育种值,提高选择的准确性[7],这可为林木多世代多性状改良提供理论依据和技术支撑。
本研究以浙江省杭州市余杭区长乐林场的杉木2 代种子园家系自由授粉子代林为研究对象,解决如下问题:(1)处理杉木2 代家系试验林木材基本密度抽样测量数据,以获得该性状若干参数;(2)估计杉木主要经济性状的遗传参数和家系育种值,逆向评选优良家系;(3)估计2 代杉木木材基本密度和胸径性状间的遗传相关,估算个体育种值,评选高世代育种亲本。
1 材料和方法
1.1 研究材料
从长乐林场杉木2 代初级种子园中挑选37 个自由授粉家系(种子园亲本间没有亲缘关系),于1997 年在长乐林场黄坑桥林区进行育苗造林试验,除参试材料外,还包括两份对照:CK2(龙15 混种)和CK4(浙江审定良种)。野外田间试验设计为完全随机区组,4 株小区,重复10 次。2009 年秋,对黄坑桥林区的这片杉木2代试验林进行调查,由于黄坑桥林区这片13 年生的杉木试验林高度郁闭,自然整枝厉害,难以测定树高等性状,同时其中的1 和2 区组人为破坏严重,全林测定了其余8 个区组重复的胸径(DBH),同时每个家系在每个小区抽取胸径最大的优株,于胸高处进行木芯取样,室内测定木材基本密度[9]。
1.2 研究方法
有鉴于该田间试验设计为正交试验类型,故可将原试验的4 株小区转化为单株小区,8 个区组转化为32 个区组重复的试验,模型为:
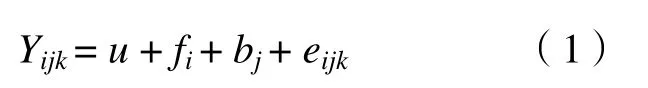
式中,i=1→a;j=1→b;k=1 或0;u为群体平均效应;fi是家系效应;bj是重复效应;eijk是随机误差。
试验获得的非平衡数据采用转化分析法进行统计分析[10],期望均方结构见表1。

表1 单因素随机区组不平衡数据的期望均方结构Tab.1 Expected mean square structure of imbalanced datasets from single-factor randomized block
主要计算以下遗传参数:
表型变异系数(PCV)=σf/群体平均值;
遗传变异系数(GCV)=σp/群体平均值;
1.3 育种值的估计及高世代亲本选择
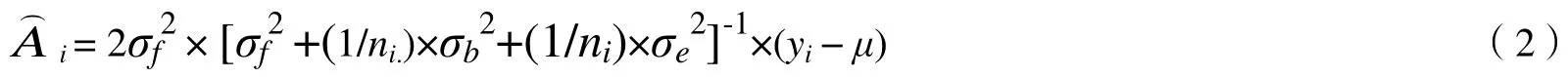
1.3.2 前向选择高世代育种亲本时的育种值 个体育种值的通用公式为:
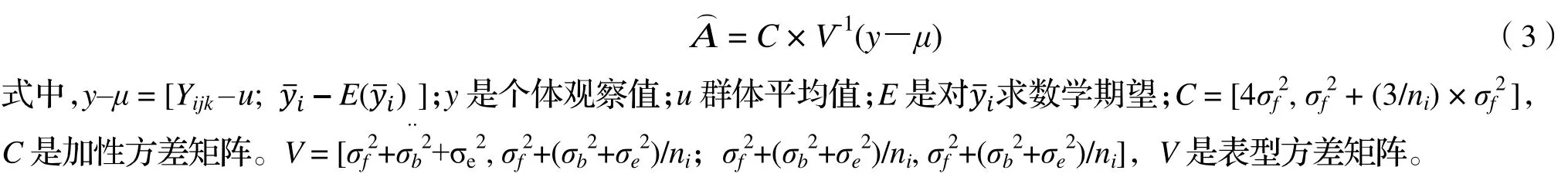
1.4 杉木木材基本密度全林数据的获得及遗传参数的估计
利用GeneXpro Tools 4.0 软件,挖掘木材基本密度与胸径间的函数关系:

式中,x是胸径(cm),F(x)是木材基本密度。
根据该函数获得全试验林的木材基本密度值,对于有实测值的样株,仍采用实测值参与木材基本密度的统计分析,获得木材基本密度若干遗传参数。在此基础上,利用抽样数据,估算木材基本密度的个体育种值。
1.5 数据处理
采用Excel 和Matlab R2016a 软件处理采样数据。应用基因表达式编程GeneXpro Tools 4.0 软件挖掘与拟合木材基本密度与胸径关系,基本参数设置:训练样本39;染色体数30;头长7;基因数2;变异量0.044;转置量0.1;基因重组参数0.1;基因连接运算符为加、减、乘和除。获得的挖掘函数如下:
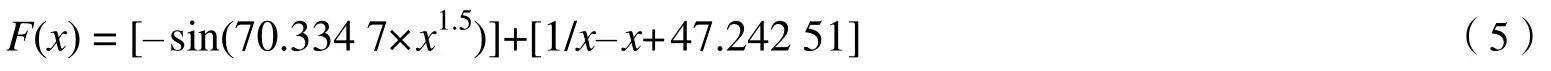
式中,木材基本密度分析时扩大100 倍。
2 结果与分析
2.1 杉木2 代自由授粉子代试验林的胸径和木材基本密度的差异分析
13 年生杉木2 代自由授粉家系的胸径平均值为14.45 cm,表型变异系数为18.79%,遗传变异系数为6.78%。由表2 可知,胸径在高世代自由授粉家系间差异显著,半同胞家系的遗传力为0.810 1,单株遗传力为0.390 9;木材基本密度的平均值为31.56,表型变异系数PCV为8.59%,遗传变异系数GCV为3.07%。
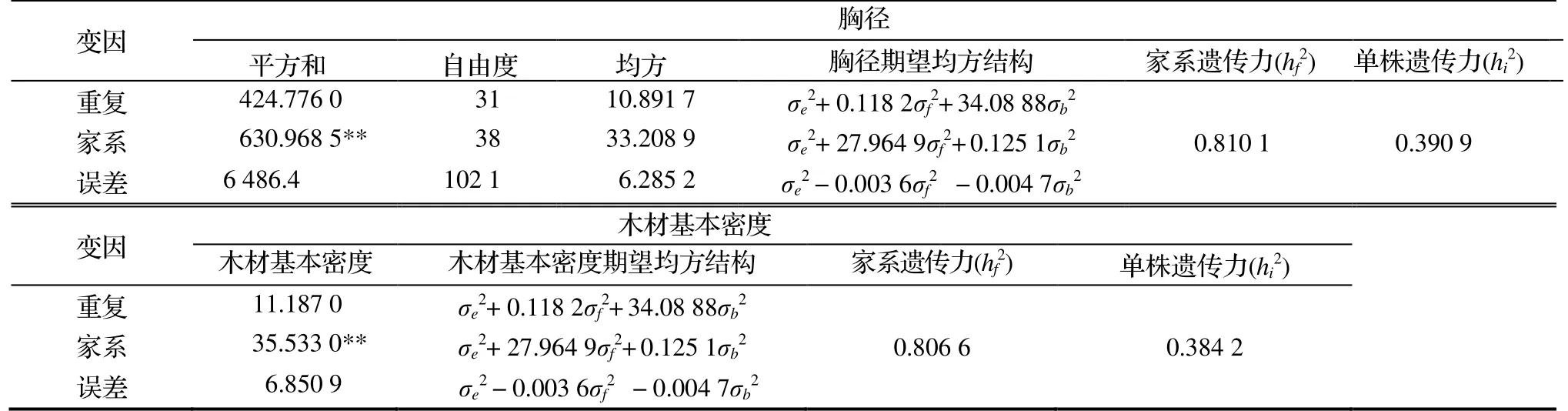
表2 随机模型条件下杉木半同胞家系的胸径和木材基本密度的方差分析结果Tab.2 ANOVA on DBH and basic density of half-sib C.lanceolata families under random model
2.2 杉木2 代自由授粉家系木材基本密度和胸径的育种值及优良家系选择
从表3 可得,胸径育种值突出的家系有8 个,分别是B42、B11、A77、B105、B101、B13、B164 和B163,其育种值均超过了1.000;而木材基本密度突出的家系有9 个,分别是C23、C30、B9、C32、J30、B111、B68、B149 和B122,其育种值也均超过了1.000。

表3 杉木高世代自由授粉家系木材基本密度和胸径的育种值及成活子代数Tab.3 Breeding values and filial generation survival number of basic density and DBH of open-pollination families in advanced generation orchard of C.lanceolata
2.3 杉木研究性状的遗传相关及高世代亲本的选择策略
木材基本密度与胸径间的遗传相关系数R=-0.981 6,这与表3 中胸径育种值突出的家系与木材基本密度育种值突出的家系不同的结果相一致,与胡德活等[11]研究结果有很大的不同。林木高世代亲本的选择策略通常是先进行逆向选择,评选优良家系,然后再在优良家系内进行前向选择优良个体(即下个世代的育种亲本)。由于本试验林中也是先进行逆向选择优良家系,然后在此基础上对入选优良家系的抽样子代进行个体育种值的估算,采用独立淘汰法,控制亲缘关系,选择优良个体。由于各家系的参试子代样本数不同,所以个体育种值须分家系求算;优良家系内个体木材基本密度的育种值(BVi)公式:BVi=C×Vi×(y–u)。
例如:本试验中木材基本密度突出的一个家系C23,有子代样本数23 个,求各子代木材基本密度的育种值:

C23 家系内的8 个体育种值如表4。

表4 C23 号家系内8 个区组重复中的个体育种值Tab.4 Individual breeding values of C23 family with 8 block repeat
其它家系中的个体育种估算仿上述过程进行,家系内个体胸径性状的育种值估算,亦仿上述过程进行。由于本试验中杉木胸径与木材基本密度呈重度负相关,高世代亲本的多性状选择,不能像常规方法那样采用构建选择指数的方式进行,而要采用独立淘汰法:入选的木材基本密度优良的个体其胸径值不得低于群体平均胸径;入选的胸径优良个体其木材基本密度值不得低于群体平均值。为了控制亲缘关系,每个优良家系内仅选1 株,最多2 株,总共评选出其18 株高世代亲本,其株号及育种值见表5。针对本试验结果,再次说明杉木高世代育种宜采用亲本互补的杂交育种的技术路线。
3 讨论与结论
本研究以13 年生杉木2 代试验林为对象,主要解决如何从试验林抽样测定方法获得的部分木材基本密度数据来获得其全林数据,从而使分析结果成为有价值的遗传参数结果,以及当改良性状间存在严重的负遗传相关时,如何评选林木高世代的育种亲本。木材基本密度与生长性状之间的关系十分复杂。由于林木高世代改良性状间的遗传相关大小影响着选择方法,故此有必要对林木的材性(木材基本密度)与生长性状的遗传相关进行讨论。Zobel 等通过对欧洲云杉Picea abies研究发现,幼龄材的木材密度和加权比重都与生长速度(包括胸径)存在着负相关,遗传相关通常最强,环境相关居中,而表型相关最弱[12]。与一般的文献进行比较看来,木材密度与生长性状间的负相关对一些林木种类是相当典型的,例如,研究赤桉Eucalyptus camaldulensis、白云杉Picea glauca、毛果杨Populus trichoocarpa、一球悬铃木Platanus occidentalis、火炬松Pinus taeda、湿地松P.ellitottii、辐射松P.radiata、欧洲赤松P.sylvestris和花旗松Pseudotsuga menziesii等树种均有报道其生长性状与木材密度呈负相关[13-14]。杉木的遗传相关参数是制订杉木育种方案的基础和依据。与其他速生林木报道的结果相似,杉木种内生长(胸径、材积)与木材密度的遗传相关也是复杂的。何贵平等[5]和胡德活等[11]报道了杉木木材基本密度与胸径、材积间存在明显的负遗传相关的结果,本研究也发现杉木木材基本密度与胸径负的遗传相关高达-0.981 6,这个结果很可能与参加试验的材料特性、造林地好的立地条件和管理等诸多因素有关。
本研究获得的其它遗传参数比较理想,但与Zhang 等在黄花落叶松Larix olgensis的研究中发现其生长性状(树高、胸径和材积)的家系遗传力分别为0.951、0.955 和0.943,而单株遗传力分别为0.958、0.983 和0.866的研究结果不同[15]。这是由于研究材料、采用的分析方法等因素不同造成的,其通过构建Henderson 方法I 的线性模型,采用混合模型条件下使用广谱性的SPSS 软件来估算研究性状的遗传参数,而本文则是采用转化理论构建统计分析的线性模型,在随机模型条件下使用Matlab R2016a 软件来处理非平衡试验数据,获得研究性状的遗传参数。通过以上比较分析还可发现,转化分析法较Henderson 方法I 更科学和精确,这与以前的研究结果相一致[10]。
综上所述,本研究表明,当林木试验林的某些研究性状只能获得部分数据时,要想办法获得全林的数据,然后采用转化分析法可获得高精度的参数。当改良性状间存在严重的负遗传相关时,高世代亲本的评选应先估算育种值,然后采用独立淘汰法进行;当对具有复杂相关的多性状进行遗传改良时,可采用主成分分析,然后选取两个主要的主分量进行独立淘汰法评选,或估算改良性状的育种值再构建选择指数,进行亲本评选[7]。
猜你喜欢
长江蔬菜(2021年12期)2021-04-04
农民致富之友(2020年8期)2020-05-11
中国果业信息(2019年11期)2019-01-05
现代园艺(2017年23期)2018-01-18
现代园艺(2017年23期)2018-01-18
西南农业学报(2016年5期)2016-05-17
广西林业科学(2016年2期)2016-03-20
广东海洋大学学报(2015年4期)2016-01-13
听力学及言语疾病杂志(2015年5期)2015-12-24
首都医科大学学报(2015年4期)2015-12-16
