
外源钙对蜜橘抗日灼生理病害的分子与生理机制研究
2023-02-21 08:04高志键李春侠周秋蓉李江波
西北植物学报 2023年1期
高志键,李春侠,周秋蓉,李江波,李 延,王 平*
(1 福建农林大学 园艺学院,园艺植物遗传育种研究所,福州 350002;2 抚州市南丰柑桔研究所,江西抚州 344100;3 福建农林大学 资源与环境学院,福州 350002)
日灼病是由温度过高和强直射太阳光所造成的一种生理病害,尤其是突发性的高温与强光更容易导致日灼发生[1]。柑橘果实发生日灼后,果实畸形,灼伤部位果皮呈现黄色或深褐色,果肉呈现海绵状,果汁变淡风味差,失去食用价值[2]。柑橘适宜种植在温暖且潮湿的气候下,若阳光长时间直射植株,使其表面温度过高,会导致果实的灼伤,尤其是树冠外围结果较多的南丰蜜橘,其日灼果实发生率可达40%以上[3]。而树冠内膛果实由于枝叶的遮挡,一般不会发生日灼。
在强光和高温胁迫下,果实表皮的生理代谢水平会失衡,产生大量的ROS分子,导致膜损伤和结构改变,进而引起酚类物质的酶促和非酶促褐变,造成果皮日灼[4]。正常情况下,细胞内产生ROS和抗氧化酶活性升高是处于一种动态平衡,不会使植物受到伤害,但当植物处于逆境中,这种平衡被打破,植物体自身的保护防御机制也随之不断变化[5]。由此可见,抗氧化酶防御机制对植物本身非常重要。张蓓等[6]研究发现,喷施螯合钙可以减缓蜜广橘果皮ROS的积累,增强抗氧化酶的活性,保持了抗氧化系统的动态平衡。Ca2+作为胞内的第二信使,在植物对各种逆境信号的转导中起着重要作用[7]。Ca2+信号通过与钙调素(calmodulin,CaM)、依赖钙激活的蛋白激酶(calcium-dependent protein kinases,CDPKs或CPKs)等相关靶蛋白结合而调节下游靶酶的活性,从而调节ROS的代谢[8],提高植物对逆境的适应性。前人研究发现CaM 拮抗剂处理可以明显降低抗氧化酶超氧化物歧化酶(SOD)、过氧化氢酶(CAT)、抗坏血酸过氧化物酶(APX)等酶活性,说明抗氧化酶活性受到钙信号转导系统的调节[9-10]。已有研究表明,Ca2+参与多种逆境胁迫过程如低温、盐胁迫、氧胁迫等,施用外源钙能提高植物的抗逆性[11-12]。而有关钙信号转导基因对蜜橘果实抗日灼作用的研究未见报道,为探讨钙信号系统对蜜橘抗氧化基因表达的影响,本研究以南丰蜜橘为试验材料,研究钙处理下不同发育阶段钙信号转导途径相关基因和抗氧化酶基因的变化以及果皮抗氧化酶活性和ROS含量的变化,以期为生产上用钙处理预防蜜橘日灼生理病害问题提供理论支持。
1 材料和方法
1.1 材料与处理
试验分别于2017和2018年的6月1日到11月10日在江西省抚州市南丰县明诚果园进行,以中熟南丰蜜橘(CitrusreticulataBlanco cv.Nanfeng tangerine) 20年结果树为试验材料。选择挂果量、树势相当,朝向一致无遮挡的果树作为试验树,每个处理6株,常规管理。分别于6月1日和7月1日17:00后对不同处理试验果树的果实进行喷施处理,以果面滴水为度。2017年预试验使用螯合钙(美国布兰特股份有限公司) 喷施浓度梯度为0(清水对照)、0.10、0.14、0.18和0.22 g·L-1;2018年选择预试验效果最好的螯合钙浓度0.14 g·L-1喷施树冠外围顶部果实,以喷施清水的树冠外围顶部果实为对照。
7-11月的每月5日采样1次,共计5次,每次每株在南和西南方向随机采10个日灼果实和10个正常果实,每次每处理共采120个果实运至实验室后,40个果实为一个重复,共3个生物学重复,日灼果取日灼处果皮,正常果取相同部位同等大小的果皮,称取适当重量的果皮,分别混合均匀,用锡箔纸分装,液氮速冻,随后置于-80 ℃冰箱保存备用。
1.2 方 法
1.2.1 日灼果发生率统计高温结束后(11月10日),分别统计钙处理和对照的果实总数和日灼果数,计算果实日灼发生率。
果实日灼发生率(%)=日灼果数/果实总数×100%
1.2.2 果皮RNA提取及相关基因表达分析基因表达量测定:参照东盛生物通用RNA试剂盒进行RNA的提取,参照诺唯赞HiScriptⅡ Q RT SperMix for qPCR试剂盒合成cDNA。
基因表达分析采用实时荧光定量PCR方法,在罗氏荧光定量PCR分析仪上完成。本研究选择了柑橘Actin基因作为内参基因,根据citrus HarvEST database(http://harvest.ucr.edu/)已知EST序列分别设计了内参基因Actin、CsNADK、CsMPK8、CsCPK等特异性引物(表1)。

表1 荧光定量PCR引物Table 1 Fluorescent quantitative PCR primers
基因扩增体系为:10 μL 2 × ChamQ Universal SYBR qPCR Master Mix,0.4 μL上/下游引物,2 μL cDNA,7.2 μL ddH2O。基因扩增体系体积20 μL。PCR反应程序:95 ℃预变性30 s;循环反应:95 ℃ 10 s,60 ℃ 30 s,45个循环;溶解曲线:95 ℃ 30 s,60 ℃ 60 s,95 ℃ 15 s。试验重复3次,按照2-ΔΔCt法计算出基因的相对表达量。
1.2.3 果皮活性氧荧光染色果皮活性氧的分布:每月每个处理分别取6个果实,用冷冻切片机对日灼处果皮(2 mm×2 mm)进行切片,切片厚度为70μm,立即用氯甲基二氯二氢荧光素二乙酯(CM-H2DCFDA)对其染色,在黑暗环境下孵育20 min,放在载玻片上,用激光共聚焦显微镜进行观察。

1.2.5 抗氧化酶活性测定酶液粗提取参照曹建康等[15]方法;SOD活性测定采用氮蓝四唑光化还原法;CAT活性测定采用 H2O2紫外吸收法;POD活性测定采用愈创木酚法;APX活性的测定参考Yu等[16]的方法;谷胱甘肽还原酶(GR)活性测定参考Hasanuzzaman等[17]的方法;谷胱甘肽过氧化物酶(GPX)活性测定参考Yu等[18]方法。
1.3 数据分析
数据采用Microsoft excel 2019处理计算平均值和标准差,采用SPSS 19.0进行差异显著性分析(P<0.05),利用Excel 2019软件作图。
2 结果与分析
2.1 钙处理后南丰蜜橘果实日灼发生率的变化
在高温结束后,对果实日灼发生率进行统计分析发现,钙处理的果实日灼发生率显著低于清水处理,日灼发生率降低了13.76%(图1)。结果表明钙处理能显著降低南丰蜜橘果实的日灼发生率。
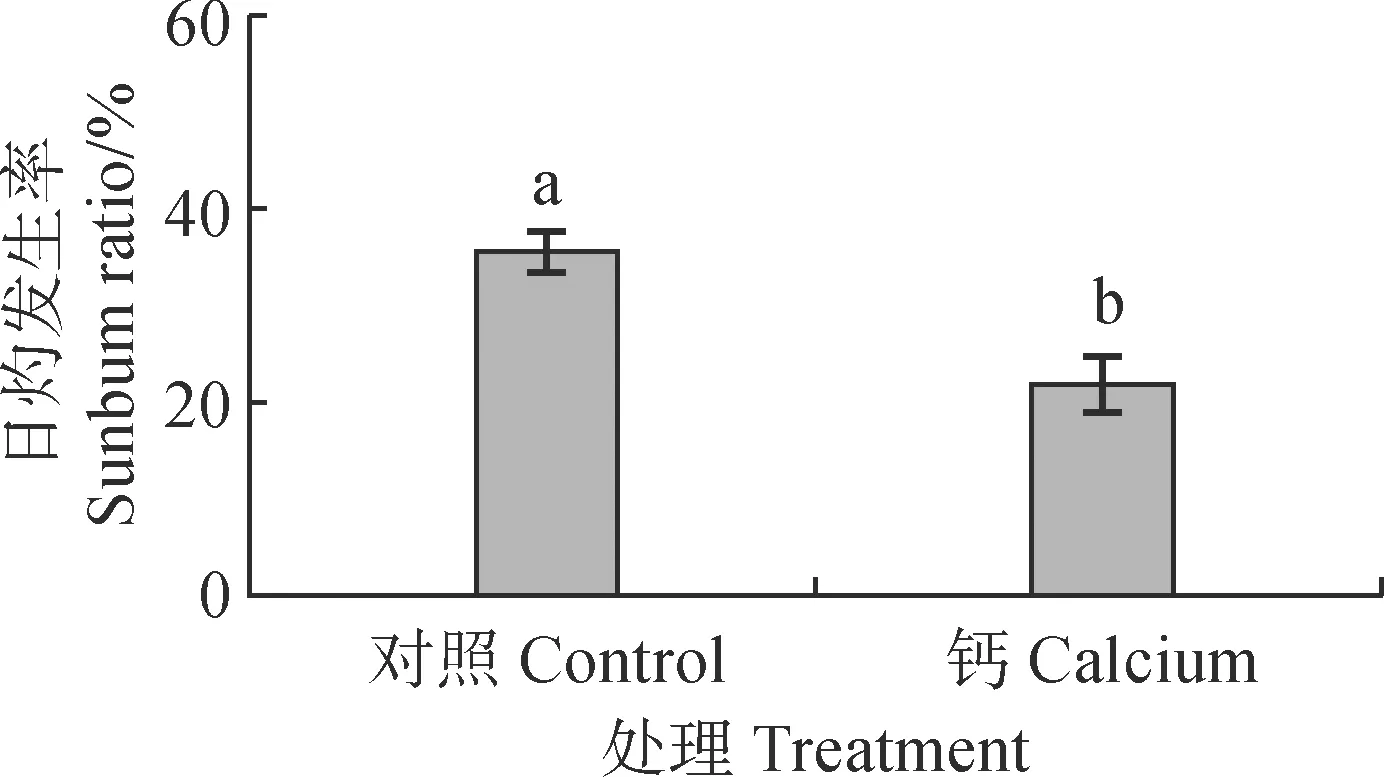
不同小写字母表示处理与对照间在0.05水平差异显著,下同图1 钙处理后南丰蜜橘果实日灼发生率变化Different normal letters indicate significant difference between treatment and control at P<0.05,the same as belowFig.1 Change of the incidence of sunburn fruits in Nanfeng tangerine after calcium treatment
2.2 钙处理后南丰蜜橘果皮钙信号转导途径相关基因的表达
图2显示了蜜橘果皮的钙信号途径相关基因相对表达量。从7月日灼病害发生至11月果实完熟,钙处理的果皮CsNADK相对表达量总体显著高于对照,在9月达到峰值;钙处理的果皮CsMPK8的相对表达量在整体上显著高于对照。从8月日灼病发生前期开始至11月果实完熟,钙处理的果皮CsCPK相对表达量始终显著高于对照。在最易发生日灼时期的7-10月,钙处理降低了蜜橘果皮CsRBOH的相对表达量。8-10月份,钙处理的果皮CsCAMK1的相对表达量均显著高于对照,在10月达到峰值。钙处理的果皮CsGAD的相对表达量从整体上显著高于对照,其中在7和8月钙处理的果皮CsGAD的相对表达量最高。说明钙处理可以显著提高蜜橘果皮钙信号途径相关基因CsNADK、CsMPK8、CsCPK、CsCAMK1和CsGAD的相对表达水平,并抑制了调控ROS产生的CsRBOH相对表达。

图2 南丰蜜橘不同发育时期果皮钙信号途径相关基因的表达Fig.2 Expressions of calcium signaling pathway related genes in peels of Nanfeng tangerine during different development stages
2.3 钙处理后南丰蜜橘果皮抗氧化酶相关基因的表达
如图3所示,对照处理和钙处理的果皮CsSOD(Cu-Zn)的相对表达量在高温和强直射光胁迫的初期(7-8月)整体较高,而后期(9-11月)显著降低,但钙处理的果皮CsSOD(Cu-Zn)相对表达量在7-11月均显著高于对照。在7-10月,钙处理果皮CsCAT的相对表达量始终高于对照。对照处理和钙处理的果皮CsPOD的相对表达量呈现出先下降后上升的趋势,但钙处理果皮CsPOD的相对表达量在整个果实发育过程中一直显著高于对照,在8月达到峰值。在蜜橘果实发育期间,对照处理和钙处理的果皮CsAPX的相对表达量呈现出先上升后下降的趋势,但钙处理果皮CsAPX的相对表达量始终显著高于对照,并于8月达到峰值。在蜜橘发育过程中,对照处理和钙处理的果皮CsGSR的表达趋势与CsSOD(Cu-Zn)的相似,钙处理的果皮CsGSR的相对表达量整体上显著高于对照。从7-9月日灼病害发生最严重时期,钙处理果皮CsGPX的相对表达量显著高于对照,在8月达到峰值。以上结果说明钙处理显著提高了蜜橘果皮抗氧化酶相关基因的表达。

图3 南丰蜜橘不同发育时期果皮抗氧化酶相关基因的表达Fig.3 Expression of antioxidant enzyme related genes in peels of Nanfeng tangerine during different development stages
2.4 钙处理南丰蜜橘果皮ROS和丙二醛含量的变化
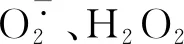
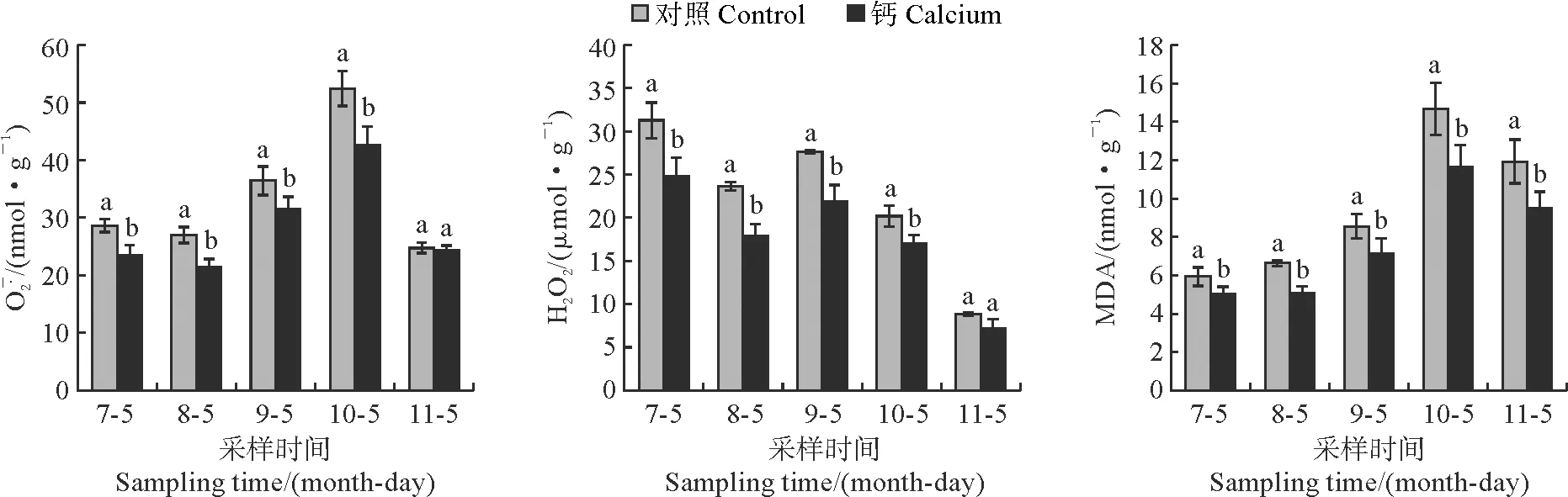
图4 南丰蜜橘不同发育时期果皮过氧化物和MDA含量的变化Fig.4 Changes of peroxide and MDA contents in peels of Nanfeng tangerine during different development stages
2.5 钙处理南丰蜜橘果皮ROS荧光强度分布的变化
由图5可以看出,在果实遭受高温胁迫初期(7月),对照处理和钙处理的外果皮和白皮层开始出现绿色荧光;在8-9月高温期,对照处理和钙处理的果皮的ROS绿色荧光已经从外果皮漫延到中果皮,但钙处理果皮ROS荧光分布扩散面积较小;10月,对照处理和钙处理的外果皮ROS荧光继续积累,钙处理果皮白皮层绿色荧光分布面积逐渐减少;11月份对照处理和钙处理的外果皮、中果皮细胞间隙急剧增大,白皮层细胞部分崩溃,ROS荧光信号部分大范围消失。在蜜橘不同的发育时期,2个处理的ROS荧光强度均呈现先上升后下降的趋势(图5)。图6表明,7-10月果实遭受高温胁迫期,对照的外果皮ROS荧光强度最高,钙处理的ROS荧光强度均显著弱于对照,二者差异显著。11月份果实进入成熟期,对照处理和钙处理果皮荧光强度急剧下降(图6)。说明果皮细胞在经过长时间ROS的毒害作用,产生ROS的细胞器(线粒体和叶绿体)严重损伤,生成ROS能力下降,导致对照处理和钙处理的果皮ROS荧光强度低于其他月份。

图5 南丰蜜橘不同发育时期果皮ROS的荧光分布Fig.5 Distribution of ROS in the peels of Nanfeng tangerine during different developmental stages

图6 南丰蜜橘不同发育时期果皮ROS的荧光强度Fig.6 Fluorescence intensity of ROS in the peels of Nanfeng tangerine during different developmental stages
2.6 钙处理南丰蜜橘果皮抗氧化酶活性变化
图7显示,在7月日灼病害开始发生到11月果实完熟,对照处理和钙处理果皮中的抗氧化酶活性均呈现出先上升后下降的趋势。说明在果实日灼病害发生初期,高温和强直射光胁迫激发了抗氧化酶活性,病害发生后期,随着ROS的积累,膜脂过氧化,使抗氧化酶的活性受到抑制。在不同的发育时期,钙处理果皮的抗氧化酶活性均高于对照。在8月份钙处理果皮的SOD、CAT、POD、APX、GR、GPX活性最高,与对照相比分别增加了35.82%、47.96%、31.48%、31.01%、47.65%和123.19%。随着时间延长,钙处理对抗氧化酶活性影响愈小。说明喷钙能在一定程度上提高抗氧化酶的活性。
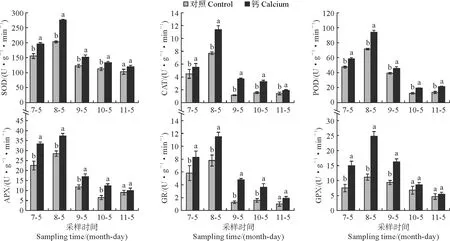
图7 南丰蜜橘不同发育时期果皮抗氧化酶活性的变化Fig.7 Changes of antioxidant enzyme activities in peels of Nanfeng tangerine during different developmental stages
3 讨 论
果皮日灼生理病害一直是蜜橘生产上的常见问题,尤其是近几年全球气温的上升逐渐使日灼病害加重。目前生产上的防控措施主要是喷水降温、果实套袋、遮阳网、喷石灰水和高岭土以及果园生草等方式[2,19],但这些方法不同程度上存在低效、可操纵性差和成本高等问题。本研究中,高温来临前期在果实表面喷施螯合钙,是一种简便有效的预防蜜橘日灼病害的生产措施。
钙是植物生长发育必需的大量营养元素之一,在植物细胞生理活动中,它不仅能维持细胞壁、细胞膜和膜结合蛋白的稳定性,还通过钙信号转导途径相关基因参与调控许多重要的生理生化过程,从而影响植物对外界环境变化的响应与适应,以增强植物对逆境的抵抗能力[9,20]。本研究通过蜜橘果皮活性氧测定和活性氧的高质荧光染色发现,在7—10月高温和强直射光胁迫下,果皮细胞产生了过量的ROS,从而产生过多的丙二醛,细胞膜脂过氧化,细胞结构破坏,细胞防御系统崩溃,导致了日灼果实的发生。前人研究表明,钙信号转导系统可以调节ROS的生成和清除,使细胞氧化-还原平衡,增加植物的抗逆性[8]。钙信号可以通过CaM控制NAD激酶 (nicotinamide asenine dinucleotide kinase,NADK)的活性,调节ROS的产生[21]。水稻中AtNADK2的过表达提高了NADP/NAD比值,从而提高植物对氧化损伤的耐受性[22]。本研究结果表明,钙处理能明显刺激CsNADK基因,增加其表达量,增强植物对高温和强直射光胁迫的抵抗力,这与前人研究结果一致。钙信号转导系统还可以通过调节植物呼吸爆发氧化酶(respiratory burst oxidase homologue,Rboh,又称NADPH氧化酶)来间接影响ROS的产生,Rboh是参与植物ROS产生的关键酶,在调节NADPH氧化酶活性产生ROS中起到重要作用[23]。有丝分裂原激活的蛋白激酶(mitogen-activated protein kinase,MAPK)是真核生物中的信号分子,参与许多生物或非生物的胁迫反应的过程[24],MPK8是目前研究较多的MAPK激酶。前人发现拟南芥中 CAM 会激活 MPK8,被激活的MPK8 通过转录控制 RbohD 表达来负调控 ROS 积累,揭示了 CAM 介导的信号转导在胁迫下微调 ROS 稳态的新机制[25]。另外CPKs的磷酸化作用也可以调节RBOH的活性[26],水稻中的OsCPK12通过诱导ROS清除基因OsAPX2/OsAPX8的上调,抑制OsRBOHI的表达,调节细胞内ROS的平衡,增加水稻的耐盐性[27]。本研究发现,钙处理显著增加CSMPK8和CSCPK基因的表达,进而显著降低CSRBOH的相对表达量,从而降低了ROS的积累,提高了蜜橘抗日灼的能力。
钙/钙调素依赖性蛋白激酶(calcium/calmodulin-dependent protein kinases,CaMKs)是真核生物钙信号途径中钙调素下游的一类重要靶蛋白,能够引起众多代谢的关键酶或转录因子磷酸化,从而完成对细胞代谢活动或某些基因表达的调节[28]。在植物中发现CaMKs有广泛的作用。如玉米的ZmCCaMK基因可以激活抗氧化防御[29]。在本研究中,钙处理使CsCAMK1基因显著上调,且在逆境中后期表达量最强,说明CsCAMK1可能参与了蜜橘抗日灼的过程。
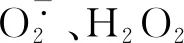
在强光和高温胁迫下,钙处理降低了南丰蜜橘果实的日灼发生率。本研究发现,钙处理可促进蜜橘果皮细胞钙信号转导途径相关基因的表达,并通过调控抗氧化系统相关基因表达以及抗氧化酶活性,清除细胞中过量的ROS,维持果皮细胞ROS代谢相对平衡,防止或降低了细胞膜脂过氧化,从而提高蜜橘果实对高温和强光的耐受性。本研究证明了果面喷施螯合钙防控日灼病害的可行性,为生产上预防和减轻蜜橘果实的日灼伤害提供了理论依据。
猜你喜欢
心声歌刊(2021年5期)2021-12-21
心声歌刊(2021年4期)2021-10-13
恋爱婚姻家庭·青春(2021年6期)2021-08-23
中小学校长(2021年7期)2021-08-21
恋爱婚姻家庭·养生版(2019年3期)2019-03-29
创新作文(小学版)(2018年25期)2018-11-30
学生导报·东方少年(2018年15期)2018-05-14
创新作文(1-2年级)(2018年9期)2018-01-22
小学生导刊(2017年15期)2017-05-17
今古传奇·武侠版(2016年5期)2016-06-08
