
甘蔗宿根矮化病病原菌致病因子pglA基因的序列分析与原核表达
2023-03-14 15:05梁永检宋修鹏颜梅新王泽平黄冬梅黎海涛韦振飞张小秋
农业研究与应用 2023年5期
梁永检 宋修鹏 颜梅新 王泽平 黄冬梅 黎海涛 韦振飞 张小秋


摘要:多聚半乳糖醛酸内切酶(endo-PG)是植物与微生物互作中极其重要的细胞壁降解酶,是病原菌定殖寄主与产生致病性的必需因子,革兰氏阴性菌Leifsonia xyli subsp. xyli(Lxx)是甘蔗宿根矮化病的致病菌,pglA是Lxx的关键致病因子。为了揭示pglA基因的功能,本研究采用重叠延伸PCR的方法获得了pglA基因的开放阅读框(ORF),ORF长1491 bp的,编码496个氨基酸,序列保守结构域包含Gly_hydro_28(Polygalacturonase)与Rho超家族,与LxxCTCB07、LxcDSM46306归为一类,相似性达100%。氨基酸序列分析显示,pglA蛋白属于疏水性蛋白,包含信号肽和跨膜区。在大肠杆菌中表达纯化获得pglA蛋白,以包涵体的形式存在于菌体中。研究结果为进一步进行Lxx的致病机制研究奠定了基础。
关键词:甘蔗;宿根矮化病;pglA基因;克隆
中图分类号:S432.1 文献标志码:A
Sequence Analysis and Prokaryotic Expression of Pathogenic Factor pglA Gene Derived from the
Sugarcane Ratoon Stunting Disease Pathogen
LIANG Yongjian2, SONG Xiupeng1, YAN Meixin1, WANG Zeping1, HUANG Dongmei1, LI Haitao3, WEI Zhenfei4, ZHANG Xiaoqiu1*
(1Sugarcane Research Institute, Guangxi Academy of Agricultural Sciences/ Guangxi Key Laboratory of Sugarcane Genetic Improvement/ Key Laboratory of Sugarcane Biotechnology and Genetic Improvement (Guangxi), Ministry of Agriculture and Rural Affairs, Nanning, Guangxi 530007, China; 2Guangxi South Subtropical Agricultural Science Research Institute, Longzhou, Guangxi 532415, China; 3Guangxi Daxin Xianggui Sugar Co., Ltd., Chongzuo, Guangxi 532300, China; 4Hengzhou Plant Protection Station, Hengzhou, Guangxi 530300, China)
Abstract: Endo-polygalacturonase (endo-PG), a crucial cell wall-degrading enzyme for plant-microorganism interaction, was an essential factor for colonization and pathogenicity of phytopathogen. Leifsonia xyli subsp. xyli (Lxx) is the pathogen of ratoon stunting disease (RSD) and pglA is the key pathogenic factor of Lxx. In order to reveal the function of pglA, through method of splice overlap extension (SOE) PCR, a 1491 bp-long opening reading frame (ORF) encoding 496 amino acids, conserved domains of Gly_hydro_28 (polygalacturonase) and Rho superfamily were found contained in pglA gene and the gene was clustered with LxxCTCB07 and LxcDSM46306 in the same group, presenting a 100% similarity between the two strains. The analysis of amino acid sequence showed that pglA protein was hydrophobic containing the signal peptides and transmembrane regions. pglA protein was expressed in Escherichia coli after purification, existing in the thallus in the form of inclusion body. The study results lays a foundation for further research on Lxx pathogenic mechanism.
Keywords: Sugarcane; ratoon stunting disease ; pglA gene; cloning
甘蔗宿根矮化病(Ratoon stunting disease,RSD) 是影響甘蔗产量和宿根性的关键病害[1, 2]。RSD由革兰氏阳性菌(Leifsonia xyli subsp. xyli, Lxx)引起。多聚半乳糖醛酸内切酶基因pglA是Lxx的关键致病因子 [3]。
多聚半乳糖醛酸酶(Polygalacturonase, PG; EC 3.2.1.15)是降解植物果胶骨架结构的主要酶之一,根据底物位置的不同,PGs可分为内切多聚半乳糖醛酸酶(endo-PG)、外切多聚半乳糖醛酸酶(exo-PG)或鼠李糖半乳糖苷酶(olio-PG)[4]。多聚半乳糖醛酸内切酶(endo-PG)是植物与微生物互作中极其重要的细胞壁降解酶,是病原菌定殖寄主与产生致病性的必需因子[5, 6]。原菌在侵染过程中上调表达endo-PG基因,在某一阶段降解植物细胞壁以获取营养供自身营养[7]。为了抵御病原菌的多聚半乳糖醛酸酶,植物细胞内信号转导、转录重组和防御代谢等合成表达[8-10]。
目前克隆表达了不少来源于不同作物不同病原的PG基因,如番茄枯萎病病原菌尖孢镰刀菌番茄专化型Fusarium oxysporum f.sp. lycopersici, FOL)的pg5基因,经克隆后纯化获得具活性的PG蛋白[11],马铃薯软腐病(Erwinia carotovora subsp. carotovora)中的peh基因,经原核表达并检测其酶活性为0.024 U·mL-1·min-1[12],烟草靶斑病菌(Rhizoctonia solani)的endoPG1和endoPG2,经克隆后分析表明endoPG1和endoPG2的推测蛋白均具有PLNO3003基因家族保守结构域,烟草互作可诱导其表达[13],剑麻斑马纹病菌(Phytophthora nicotianae)中的5个PG基因,Szpg1-Szpg5,在被检测的剑麻斑马纹病菌中均被成功克隆[14]。但甘蔗宿根矮化病菌Lxx中的PG基因并未见有克隆表达的,为了揭示pglA的基因功能,本研究克隆并表达了pglA基因,并对其序列进行了生物信息学分析,为进一步进行Lxx的致病机制研究奠定基础。
1 材料与方法
1.1 pglA基因的扩增
从NCBI中调取Lxx中的pglA基因序列(登录号:AE016822.1;Locus tag:Lxx07750),通过密码子优化方法以及采用重叠延伸PCR方法设计56条引物,分成两个片段,第1~28条引物扩增第一个片段,第27~56条引物扩增第二个片段,进行两轮PCR扩增。PCR反应采用pfu高温聚合酶,第一轮PCR分别使用第1~28条引物和第27~56条引物进行2个体系的扩增,引物1~28的扩增体系为,引物mix1(引物1~28)各0.4 μL,共11.2 μL;dNTP 1 μL(每條25 mM);10ⅹ pfu Buffer 5 μL;1号和28号引物各2 μL,Pfu(5 u/μL) 0.4 μL,补充ddH2O至50 μL。引物27~56的扩增体系,使用引物mix2(引物27~56)各0.4 μL,27号和56号引物各2 μL,其它组分同引物1~28的扩增体系。两次PCR反应程序为,95℃ 3 min,95℃ 22 s,55℃ 20 s,72℃ 50 s,18个循环,72℃ 5 min。第二轮PCR反应体系为,1号引物和56号引物各2 μL,第一轮PCR扩增产物各1 μL,dNTP(每条25 mM) 1 μL, Pfu(5 u/μL) 0.4 μL,补充ddH2O至50 μL。PCR反应程序为,95℃ 3 min,95℃ 24 s,58℃ 20 s,72℃ 80 s,22个循环,72℃ 5 min。
使用SanPrep 柱式 DNA 胶回收试剂盒(生工,上海)回收纯化目的片段。将纯化的目的片段与pMD-18T载体连接,并热激导入感受态大肠杆菌DH5α,在LB固体平板中添加X-gal、氨苄青霉素和IPTG进行蓝白斑筛选。挑取白色阳性菌落进行LB液体振荡培养,经通用引物PCR扩增验证目的大小正确后,测序验证。
1.2 pglA基因的生物信息学分析
通过BioXM分析pglA的氨基酸序列,利用NCBI上的CD search在线软件分析pglA的保守域,利用ExPASy服务器中的Protparam (https://web.expasy.org/protparam/)工具分析蛋白的理化性质,利用ExPASy软件中的ProtScale分析pglA蛋白的疏水性。使用TMHMM Server v. 2.0 (https://servi ces.healthtech.dtu.dk/service.php?TMHMM-2.0) , Signa lP-5.0 (https://services.healthtech.dtu.dk/service.php?SignalP-5.0) 分别进行跨膜区和信号肽的预测分析。使用SOPMA (https://npsa-prabi.ibcp.fr/cgi-bin/npsa_automat.pl?page=npsa_sopma.html)进行二级机构预测,使用NCBI中的Blast软件对pglA进行比对后,使用MEGA6对选取的序列构建系统进化树。
1.3 pglA基因表达载体的构建与转化
将PCR产物进行酶切,酶切体系为,纯化好的目的基因1 μg,10ⅹFD buffer 5 μL,NdeI (10 u/μL)1 μL,XhoI(10 u/μL)1 μL,ddH2O 23 μL,共50 μL,放入37 ℃恒温水浴锅中反应2 h。同时,将pET21a载体按以上的体系进行酶切。使用SanPrep 柱式DNA胶回收试剂盒(生工,上海)回收纯化酶切的载体和目的片段,将回收纯化的载体与目的片段连接,连接体系20 μL,目的片段:8 μL,载体pET21a 4 μL,10ⅹ T4 DNAligase Buffer 2 μL,T4 DNAligase 1 μL (5 u/ul),ddH2O 补充至20 μL。将连接液在22℃ PCR仪上孵育1 h,完成后将连接液热激转入top10感受态中,检测筛选出阳性克隆后测序验证。PCR验证引物为:上游引物5-ATGCGCACACGACTGCCTGC-3和下游引物5-CAGGGGGCGGGATAGGTCGG-3。
1.4 pglA蛋白的表达检测与纯化
将重组质粒转入Rosetta感受态中,热激后涂布于含氨苄抗性的平板上,单菌落长出后,挑取单菌落至含有氨苄抗生素的液体培养基中培养,当OD600值达到600时,添加1 mM IPTG诱导表达,分别于20℃条件下培养16 h、37℃ 条件下培养4 h,未添加诱导剂的为阴性对照。12000 rpm离心,弃上清,收集菌体,在收集到的菌体中加入缓冲液A(PBS, pH 7.4)悬浮,超声破碎仪破碎,12000 rpm,4℃离心,收集沉淀和上清。沉淀使用缓冲液B(8 M Urea, 50 mM Tris-HCl, 300 mM NaCl, pH 8.0)进行溶解,使用SDS-PAGE分别对上清和沉淀进行表达检测。
使用缓冲液C(8 M Urea, 50 mM Tris-HCl, 300 mM NaCl, 0.1% Triton X-100, pH 8.0)溶解细胞菌体,超声破碎,离心收集上清粗蛋白,用于蛋白纯化。取5 mL Ni-NTA,用5倍柱床体积的Binding buffer清洗平衡柱子,流速5 mL/min,将粗蛋白与平衡后的柱填料孵育1 h,将孵育后的产物上柱,收集流出。用Binding buffer清洗平衡柱子,用Binding buffer清洗平衡柱子,用Elution buffer洗脱,收集流出。对粗蛋白、洗杂流出、洗脱流出分别处理,制樣,准备 SDS-PAGE纯化检测。
2 结果分析
2.1 pglA基因的克隆
电泳结果显示,在1200~1500 bp大小处出现单一条带的目的带(图1A),载体酶切结果显示在5443 bp处有单一大小的条带(图1B),表明条带正确。蓝白斑筛选的PCR产物,条带大小正确(图1C)。
2.2 pglA基因编码的氨基酸序列及其理化性质
经测序比对分析,获得了pglA基因的的ORF(图2A),ORF长1491 bp,编码496个氨基酸,分子量50.63 kDa,等电点(pI)5.23(图2B)。序列保守结构域包含Gly_hydro_28(Polygalacturonase)与Rho超家族(图2C,D)。将序列上传至NCBI,登录号为MG983518。
PortScale 结果显示,疏水性最大值与亲水性最小值之和为正值,因此推测pglA蛋白为疏水性蛋白(图3)。
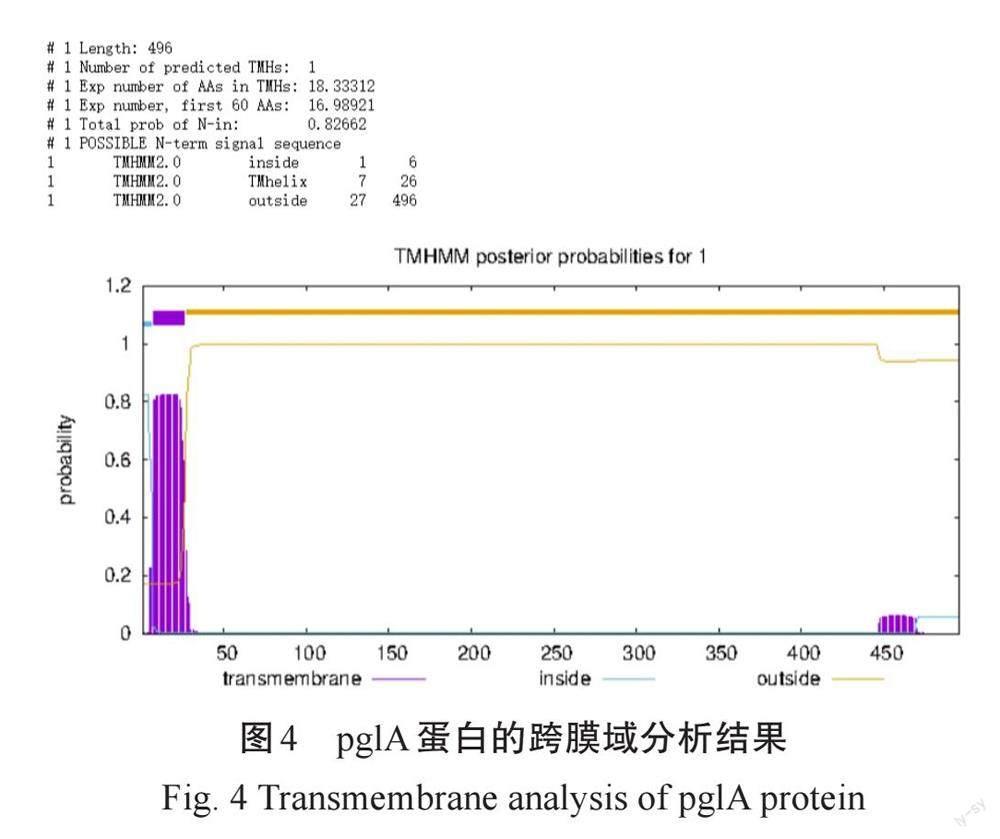
TMHMM预测结果显示,pglA有一个跨膜螺旋,Exp number, first 60 AAs数值为16.98921,也说明存在的跨膜螺旋很可能是信号肽(图4)。
Signal P-5.0 预测结果显示,Signal peptide (Sec/SPI)值为0.8999,TAT Signal peptide (Tat/SPI) 值为0.0389,Lipoprotein signal peptide (Sec/SPII) 值为0.0563,Other值为0.0049,推测pglA为分泌蛋白,存在信号肽结构(图5)。
SOPMA软件预测pglA的二级结构包含5.85%的α螺旋、29.23%的延伸,64.92%的卷曲(表1,图6)。NCBI比对显示克隆获得的pglA与LxxCTCB07、LxcDSM46306归为一类,相似性达100%(图7)。
2.3 pglA蛋白的表达与纯化
对重组质粒进行双酶切验证,电泳结果显示酶切获得5443 bp大小的质粒和1491 bp大小的目的带,克隆测序也表明序列正确,说明重组质粒构建成功(图8)。
SDS-PAGE结果显示,pglA蛋白在IPTG诱导前未表达,IPTG诱导后,表达存在于沉淀中,以包涵体的形式存在,20℃和37℃均能诱导蛋白的大量表达(图9A)。咪唑能对纯化蛋白有较好的洗脱效果,50 mM和500 mM洗脱后均获得单一的目的带,效果好于20 mM(图9B)。
3 讨论
MONTEIRO-VITORELL等[3]在2004年报道Lxx基因组中存在pglA,但至今未见关于pglA基因表达与功能研究的相关研究。本研究分析了pglA的基因序列特征,构建了融合表达载体,纯化获得pglA蛋白,为进一步进行pglA的互作基因的筛选及致病性功能等研究奠定了基础。
生物信息学分析显示,pglA中存在信号肽与跨膜域,在后续构建诱饵菌株用于酵母杂交筛选互作基因时,应注意信号肽的去除[15]。Gly_hydro_28家族包含多聚半乳糖醛酸酶 (PG)、 鼠李聚糖半乳糖醛酸酶(RG)和木糖聚半乳糖醛酸酶(XG),这些酶在细胞壁代谢中有重要的作用[16]。pglA的保守域分析显示其包含保守域Gly_hydro_28(位置从577-1320),表明Lxx自身具有分泌多聚半乳糖醛酸酶的能力。在作者的前期研究中,发现甘蔗感染宿根矮化病后,蔗茎细胞壁溶解[17],也说明Lxx可能是通过分泌多聚半乳糖醛酸酶来使细胞壁溶解,以穿梭于甘蔗组织内。
重叠延伸PCR(Splice overlap extension PCR, SOE PCR)根据目的基因序列,设计多对具有互补末端的引物,分段进行扩增,使PCR产物形成重叠链,经过重叠链的相互搭桥、互为模板来延伸,从而将来源不同的扩增片段重叠拼接起来,可快速、简洁地获得高质量的PCR产物,目的产物特异性高。该技术广泛应用于大片段基因的人工合成、基因定点突变等方面[18]。Lxx体外难培养,且不同菌株间基因组相似性高达93%,pglA序列间无突变碱基[17],在已知pglA基因序列的情况下,采用重叠延伸PCR的方法合成pglA序列,可快速、准确地获得基因片段,保证下游实验的顺利进行。在微生物中表达异源蛋白是获得酶最简单有效的方法,密码子优化是实现异源蛋白高效表达的关键技术方法,可获得高浓度和高质量的异源蛋白[19]。本研究中,通过密码子优化,在大肠杆菌中表达了pglA蛋白,通过镍柱纯化获得了pglA蛋白,下一步将进行纯化蛋白的活性和致病性研究。
参考文献
[1] LI Y R, YANG L T. Sugarcane agriculture and sugar industry in China[J]. Sugar Tech, 2015, 17(1): 1-8.
[2] VISWANATHAN R. Varietal degeneration in sugarcane and its management in India[J]. Sugar Tech, 2016, 18(1): 1-7.
[3] MONTEIRO-VITORELLO C B, CAMARGO L E A, van SLUYS M A, et al. The genome sequence of the gram-positive sugarcane pathogen Leifsonia xyli subsp. xyli[J]. Molecular Plant-Microbe Interactions: MPMI, 2004, 17(8): 827-836.
[4] COOK B J, CLAY R P, BERGMANN C W, et al. Fungal polygalacturonases exhibit different substrate degradation patterns and differ in their susceptibilities to polygalacturonase-inhibiting proteins[J]. Molecular Plant-Microbe Interactions: MPMI, 1999, 12(8): 703-711.
[5] BASHI Z D, RIMMER S R, KHACHATOURIANS G G, et al. Brassica napus polygalacturonase inhibitor proteins inhibit Sclerotinia sclerotiorum polygalacturonase enzymatic and necrotizing activities and delay symptoms in transgenic plants[J]. Canadian Journal of Microbiology, 2013, 59(2): 79-86.
[6] TAYI L, MAKU R V, PATEL H K, et al. Identification of pectin degrading enzymes secreted by Xanthomonas oryzae pv. oryzae and determination of their role in virulence on rice[J]. PLoS One, 2016, 11(12): e0166396.
[7] LIU N N, MA X W, SUN Y, et al. Necrotizing activity of Verticillium dahliae and Fusarium oxysporum f. sp. vasinfectum endopolygalacturonases in cotton[J]. Plant Disease, 2017, 101(7): 1128-1138.
[8] AKAGI A, ENGELBERTH J, STOTZ H U. Interaction between polygalacturonase-inhibiting protein and jasmonic acid during defense activation in tomato against Botrytis cinerea[J]. European Journal of Plant Pathology, 2010, 128(4): 423-428.
[9] DANGL J L, HORVATH D M, STASKAWICZ B J. Pivoting the plant immune system from dissection to deployment[J]. Science, 2013, 341(6147): 746-751.
[10] KALUNKE R M, TUNDO S, BENEDETTI M, et al. An update on polygalacturonase-inhibiting protein (PGIP), a leucine-rich repeat protein that protects crop plants against pathogens[J]. Frontiers in Plant Science, 2015, (6): 146.
[11] GARC?A-MACEIRA F I, DI PIETRO A, HUERTAS-GONZ?LEZ M D, et al. Molecular characterization of an endopolygalacturonase from Fusarium oxysporum expressed during early stages of infection[J]. Applied and Environmental Microbiology, 2001, 67(5): 2191-2196.
[12] 李洋, 張俊莲, 白江平, 等. 马铃薯软腐病菌Erwinia carotovora subsp. carotovora 714种胞外酶基因的克隆和原核表达分析[J]. 草业科学, 2014, 31(4): 561-574.
[13] 赵艳琴, 吴元华, 赵秀香, 等. 烟草靶斑病菌内切多聚半乳糖醛酸酶基因endoPGs的克隆及表达特征分析[J]. 中国农业科学, 2014, 47(10): 1939-1946.
[14] 吴伟怀, 梁艳琼, 郑金龙, 等. 剑麻斑马纹病菌5个多聚半乳糖醛酸酶基因的克隆与序列分析[J]. 热带作物学报, 2015, 36(12): 2204-2209.
[15] HOLCROFT J, GANSS B. Identification of amelotin- and ODAM-interacting enamel matrix proteins using the yeast two-hybrid system[J]. European Journal of Oral Sciences, 2011, 119 (Suppl 1): 301-306.
[16] VILLARREAL F, STOCCHI N, TEN HAVE A. Functional classification and characterization of the fungal glycoside hydrolase 28 protein family[J]. Journal of Fungi, 2022, 8(3): 217.
[17] ZHANG X Q, CHEN M H, LIANG Y J, et al. Morphological and physiological responses of sugarcane to Leifsonia xyli subsp. xyli infection[J]. Plant Disease, 2016, 100(12): 2499-2506.
[18] URBAN A, NEUKIRCHEN S, JAEGER K E. A rapid and efficient method for site-directed mutagenesis using one-step overlap extension PCR[J]. Nucleic Acids Research, 1997, 25(11): 2227-2228.
[19] 楊云彭, 马晓焉, 霍毅欣. 密码子优化策略在异源蛋白表达中的应用[J]. 生物工程学报, 2019, 35(12): 2227-2237.
责任编辑:谢红辉
基金项目:广西自然科学基金项目(2020GXNSFBA297040;2023GXNSFAA026495);国家自然科学基金青年科学基金项目(32001606);广西科技重大专项(桂科AA22117004;桂科AA22117002);广西创新团队建设项目(nycytxgxcxtd-2021-03-06);广西农业科学院基本科研业务专项(桂农科2021YT007)。
第一作者:梁永检(1988—),男,助理研究员,研究方向甘蔗育种与无人机遥感,E-mail:724808829@qq.com。
*通信作者:张小秋(1989—),女,副研究员,研究方向为甘蔗植保防控,E-mail:ZhangXiaoQiuXHD@163.com。
收稿日期:2023-08-06
猜你喜欢
儿童故事画报·自然探秘(2022年12期)2022-11-24
环球时报(2022-09-20)2022-09-20
基层中医药(2021年3期)2021-11-22
今日农业(2020年24期)2020-12-15
小猕猴学习画刊(2019年8期)2019-09-16
小学科学(学生版)(2019年5期)2019-05-21
特别健康(2018年3期)2018-07-04
中国糖料(2016年1期)2016-12-01
作文大王·低年级(2016年9期)2016-09-21
兽医导刊(2016年12期)2016-05-17
