
旱胁迫相关绿豆VrERF家族基因发掘及表达分析
2023-05-06 11:14李蒙鑫丁冬会张雨朋景琳超鲁建荣刘嘉斐陈吉宝杨树琼
四川农业大学学报 2023年2期
李蒙鑫,丁冬会,张雨朋,景琳超,鲁建荣,刘嘉斐,陈吉宝,杨树琼*
(1. 河南省南水北调中线水源区生态安全重点实验室,河南 南阳 473061;2. 河南省伏牛山昆虫生物学重点实验室,河南 南阳 473061)
绿豆(Vigna radiate(L.) Wilczek),别名菉豆、植豆等,是我国重要的粮食经济作物,在中国已经有两千多年的栽培历史。绿豆在生长过程中始终处于一种土壤条件,不可自行移动,具有固定性,其在整个生长发育过程中非常容易遭受干旱、盐碱和低温等非生物胁迫影响[1]。干旱胁迫不仅是限制绿豆生命活动的限制因子,也是决定其地理分布及限制其产量的主要因素[2]。部分绿豆具有一定的抗旱能力,但是在长期干旱的地区,需要为其定期补充水分,否则会对绿豆产量造成一定的影响。目前国内种植主要集中在北方,内蒙古、吉林和黑龙江等省份干旱贫瘠的山区丘陵地带[3],且大部分绿豆产区自然降雨量少,灌溉条件差。有研究表明,干旱胁迫会对绿豆的光合作用产生抑制作用,光化学效率降低导致光能过剩,其光合器官会因此遭到破坏[2]。而且随着干旱胁迫程度的增加,绿豆苗期的植株光合速率、蒸腾速率和气孔导度都会逐步减弱,并且这些指标相互之间呈现显著的正相关,随着干旱胁迫时间的增加,叶绿素含量和叶片的含水量呈现低—高—低这样的一个变化趋势,进而影响绿豆种子的品质以及产量[4]。
干旱胁迫不仅会影响植物的形态建成[5],还会诱导植物相关基因的表达调控,对其作出反应的主要调控基因就是转录因子。作为植物体内广泛存在的超大转录因子家族之一,AP2/ERF 转录因子最先在拟南芥中被发现[6],依据所含的AP2/ERF 结构域个数及其结构域同源性,将拟南芥AP2/ERF转录因子分为5 个亚家族:AP2、ERF、DREB、RAV 和Soloist。AP2/ERF家族基因功能也呈现多样化,研究发现AP2/ERF家族参与了植物的高温、干旱、盐碱以及机械损伤等非生物逆境响应过程[7-8]。AP2/ERF 转录因子可通过调节下游基因或相互合作来抵抗干旱胁迫,过表达ZmERF21显著增加了玉米叶绿素含量和抗氧化酶活性,并通过调节激素信号和胁迫应答基因的表达来增强玉米幼苗的耐旱性[9]。水稻OsERF83可调控旱胁迫相关基因的表达,过表达OsERF83显著提高了水稻的耐旱性[10]。大豆GmDREB8可负向调控植物的耐旱能力,异源表达GmDREB8转基因拟南芥比野生型对干旱胁迫更敏感,而大豆GmDREB8沉默株系表现出更强的耐旱性[11]。水稻OsERF3基因含有EAR抑制结构域基序,对干旱胁迫起负调控作用[12]。上述研究表明,AP2/ERF 转录因子在植物应对干旱胁迫的分子调控机制中起着至关重要的作用。
AP2/ERF 转录因子对研究植物响应逆境胁迫生理过程具有十分重要的价值,然而目前为止,对绿豆AP2/ERF家族基因的逆境响应及表达调控机制研究较少。本课题组前期已筛选到87 个绿豆VrERF家族基因,并对其结构特征、理化性质等进行了初步分析。为进一步探究VrERF家族基因在绿豆逆境胁迫中的应答机制,本研究对VrERF家族基因的种间同源性、互作蛋白功能及顺式作用元件进行了预测分析,并通过转录组测序数据分析了各Vr-ERF基因的组织特异性表达、干旱胁迫下基因表达差异情况,筛选了2 个与干旱胁迫相关的VrERF基因并进行了qRT-PCR 验证。此研究将为进一步开展干旱胁迫相关基因的功能分析奠定了基础,也将为利用转录调控、基因育种等手段培育绿豆抗逆新品种提供理论依据。
1 材料和方法
1.1 绿豆VrERF 家族基因的种间同源性、互作蛋白功能预测与启动子区顺式作用元件分析
绿豆、豌豆、菜豆与大豆的基因组及转录组数据下载自NCBI-genome(https://www.ncbi.nlm.nih.gov/data-hub/genome/)和PlantBioinfoPF(https://urgi.versailles.inra.fr/Species/Pisum),各物种间全基因组比对、ERF家族基因间的共线性可视化使用Java 环境下的MCScanX 和TBtools 软件进行分析。VrERF蛋白互作网络分析使用String 蛋白互作数据库(http://string-db.org/),物种参数选择模式植物拟南芥。使用TBtools 提取VrERF家族基因起始密码子上游2 000 bp 序列,启动子区顺式作用元件预测分析使用在线软件PlantCARE(http://bioinformatics.psb.ugent.be/webtools/plantcare/html/),各顺式作用元件的可视化分析使用TBtools进行。
1.2 绿豆VrERF 家族基因的组织特异性表达、干旱胁迫下的表达及验证分析
从NCBI-SRA(https://www.ncbi.nlm.nih.gov/sra)下载绿豆发育几个关键部位:根、下胚轴、叶、花、茎尖和果实的表达数据,根据表达量进行绿豆VrERF家族基因的组织特异性表达分析,使用TBtools绘制热图。根据NCBI-SRA中正常供水和干旱胁迫处理的绿豆栽培种(VC1973A)和野生型(JP226873)V1期幼苗的叶片转录组测序数据,筛选VrERF家族基因的FPKM值,使用TBtools绘制热图。
对盆栽的绿豆栽培种(VC1973A)和野生型(JP226873)V1期幼苗进行20% PEG6000 溶液处理来模拟干旱胁迫,提取处理后0、6 及24 h 下的绿豆叶片RNA 后反转录成cDNA,以绿豆基因VrActin(Vradi03g00210)作为内参基因,在Bio-RAD荧光定量PCR 仪上采用两步法进行qRT-PCR 分析,按照2-ΔΔCt法计算VrERF7和VrERF62基因在不同处理时间下的相对表达量。
2 结果与分析
2.1 绿豆VrERF 与豌豆、菜豆及大豆ERF 家族基因的同源性
前期研究结果证实了VrERF 蛋白与其他物种ERF蛋白在结构上具有高度的相似性,本研究进一步分析了VrERF与PsERF、PvERF和GmERF家族基因间的同源性(图1)。各物种ERF家族基因间的同源性分析结果显示,PsERF和VrERF家族基因之间有62个同源基因对,但并不是所有的ERF基因间存在着1对1关系,如2对1(Psat2g181560/Psat6g201400-VrERF42)、3 对1(Psat2g181560/Psat4g067400/Psat6g201400-VrERF45)等。PvERF和VrERF家族基因之间有126 个同源基因对,同样存在基因间多对一关系,如2 对1(PHAVU_008G046400g/PHAVU_010G050600g-VrERF21)、4对1(PHAVU_001G031200g/PHAVU_002G009100g/PHAVU_003G232600g/PHAVU_009G074300g-VrERF76)。GmERF和VrERF家族基因之间有194 个同源基因对,基因间多对一关系更普遍,仅有9个同源基因对是1对1关系。综上可推测VrERF家族基因与其他豆科物种ERF基因间应该具有比相似的功能特征。
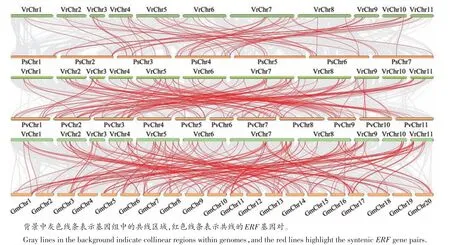
图1 绿豆与豌豆、菜豆、大豆ERF家族基因的共线性分析Figure 1 Synteny analysis of ERF genes between mung bean and pea,kidney bean,and soybean
2.2 VrERF互作蛋白的功能预测
以拟南芥为参考物种,利用string 蛋白数据库对VrERF互作蛋白进行了预测分析,部分结果如图2所示。VrERF25/38/40/42/43/44/45/46/48/58 同源蛋白CBF4不仅与VrERF1/13/14/17/18/19/21同源蛋白ERF13 的互作蛋白都包括bZIP,还与VrERF51/52/53/56/57 同源蛋白TINY2 的互作蛋白都包括AP2。VrERF1/13/14/17/18/19/21 同 源 蛋 白ERF13 和Vr-ERF2/33/35/71 同源蛋白ERF-1 的互作蛋白都包括STZ 和WRYK40;VrERF9/10/11/23/30/34/70 同源蛋白TDR1 的 互 作 蛋 白 包 括AMR1、CSN5B 和WRYK30 等;VrERF72/84/85/86/87 同 源 蛋 白PLT1的互作蛋白包括SMB、SHR 和RGF1 等;VrERF78/79/80/81同源蛋白ADAP 的互作蛋白包括ZFP1、BCCP2 和ADCL 等;VrERF73/74/75/76 同 源 蛋 白ANT的互作蛋白包括WUS、CYP78A5和JAG等。综上结果推测绿豆VrERF家族基因与同缘关系较近的拟南芥AtERF13、AtERF028及AtERF98等具有相似的生物学功能,VrERF 通过与以上蛋白互作参与绿豆生长发育过程。
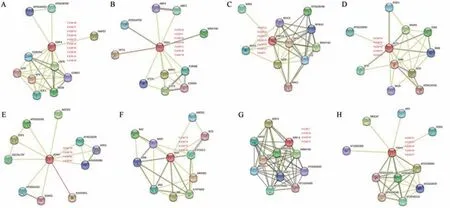
图2 VrERF互作蛋白功能预测网络图Figure 2 Prediction of functional network of interacting proteins of VrERF
2.3 VrERF家族基因启动子区顺式作用元件分析
提取绿豆VrERF家族基因上游2 000 bp的基因组序列,利用PlantCARE 数据库对87个VrERF基因启动子区的顺式作用元件进行了分析(图3)。绿豆VrERF基因启动子区的顺式作用元件可分为生长发育相关、胁迫响应及激素响应元件,其中生长发育相关元件包含胚乳表达、分生组织表达等元件,胁迫响应元件包含缺氧、干旱和低温防御等相关元件,激素响应元件有赤霉素、脱落酸和生长素等元件。光响应(453 个)、脱落酸响应(180 个)、厌氧诱导(180 个)及茉莉酸甲酯响应(136 个)元件是绿豆VrERF家族基因启动子序列所具有的主要顺式作用元件,其中光响应元件在几乎所有VrERF基因的启动子区均有分布,除VrERF58(0 个)和VrERF14(1个)外,其余基因包含多个光响应元件,这表明绿豆VrERF家族基因可能参与了植株的光形态建成或光照相关的环境适应。启动子区具有脱落酸、茉莉酸甲酯、赤霉素和生长素等激素响应元件的绿豆VrERF基因数量分别为72、63、39及33个,说明绿豆VrERF家族基因广泛参与了不同植物激素的信号途径。此外,36个VrERF基因启动子区存在干旱响应元件,24 个VrERF基因启动子区存在低温响应元件,这部分基因也参与了绿豆干旱、低温等非生物逆境响应过程。

图3 绿豆AP2/ERF基因的激素和非生物胁迫响应相关的顺式作用元件分析Figure 3 Analysis of cis-acting elements related to hormone and abiotic stress response of mung bean AP2/ERF genes
2.4 VrERF家族基因的组织特异性表达分析
利用TBtools 对绿豆87 个VrERF家族基因的根、下胚轴、叶、茎尖、花和果实6个不同器官组织的表达情况进行了分析(图4)。结果显示,在整个绿豆发育过程的各个组织中均有VrERF家族基因表达,但不同基因在不同器官组织的表达模式差异较为明显,其中46个基因在所有6个器官组织中均有表达,6 个基因只在花中有表达。依据基因表达量来看,多数VrERF家族基因的表达具有较强的组织特异性,如VrERF7/17/22/26在根中特异性的高表达,VrERF2/4/8/24/49/61/62在下胚轴中特异性的高表达。值得注意的是,VrERF4/7/62在所有组织器官中均有较高的表达,VrERF4在花中的表达量显著高于其他基因,VrERF7在根、下胚轴、茎尖和果实中的表达量显著高于其他基因,VrERF62在叶中的表达量显著高于其他基因。综上结果表明,不同Vr-ERF家族基因在绿豆各组织器官中功能具有差异性。

图4 VrERF在绿豆VC1973A不同器官组织中的基因表达情况Figure 4 Expression levels in different organs of VrERFs in mung bean VC1973A
2.5 VrERF 家族基因在干旱胁迫下的表达模式分析
与正常浇水相比,绿豆栽培种(VC1973A)和野生型(JP226873)在干旱胁迫处理下,绿豆VrERF基因家族中共检测到68 个基因有差异表达,19 个基因未检测到差异表达(图5)。与正常浇水相比,干旱胁迫的VC1973A 叶片中有45 个VrERF基因表达下调,其中VrERF7/22/62显著下调;18 个基因表达上调,但均未显著上调。干旱胁迫的JP226873叶片中有50 个VrERF基因表达下调,其中VrERF7/62显著下调;10 个基因表达上调,其中VrERF22显著上调。未受胁迫时,VC1973A 叶片中表达量高于JP226873的VrERF基因数量为37,表达量等于或低于JP226873 的基因数量也为37;干旱胁迫后,VC1973A 叶片中表达量高于JP226873 的VrERF基因数量为30,表达量等于或低于JP226873的基因数量同样为30;表明绿豆VC1973A 和JP226873 对干旱胁迫的响应没有明显差异。
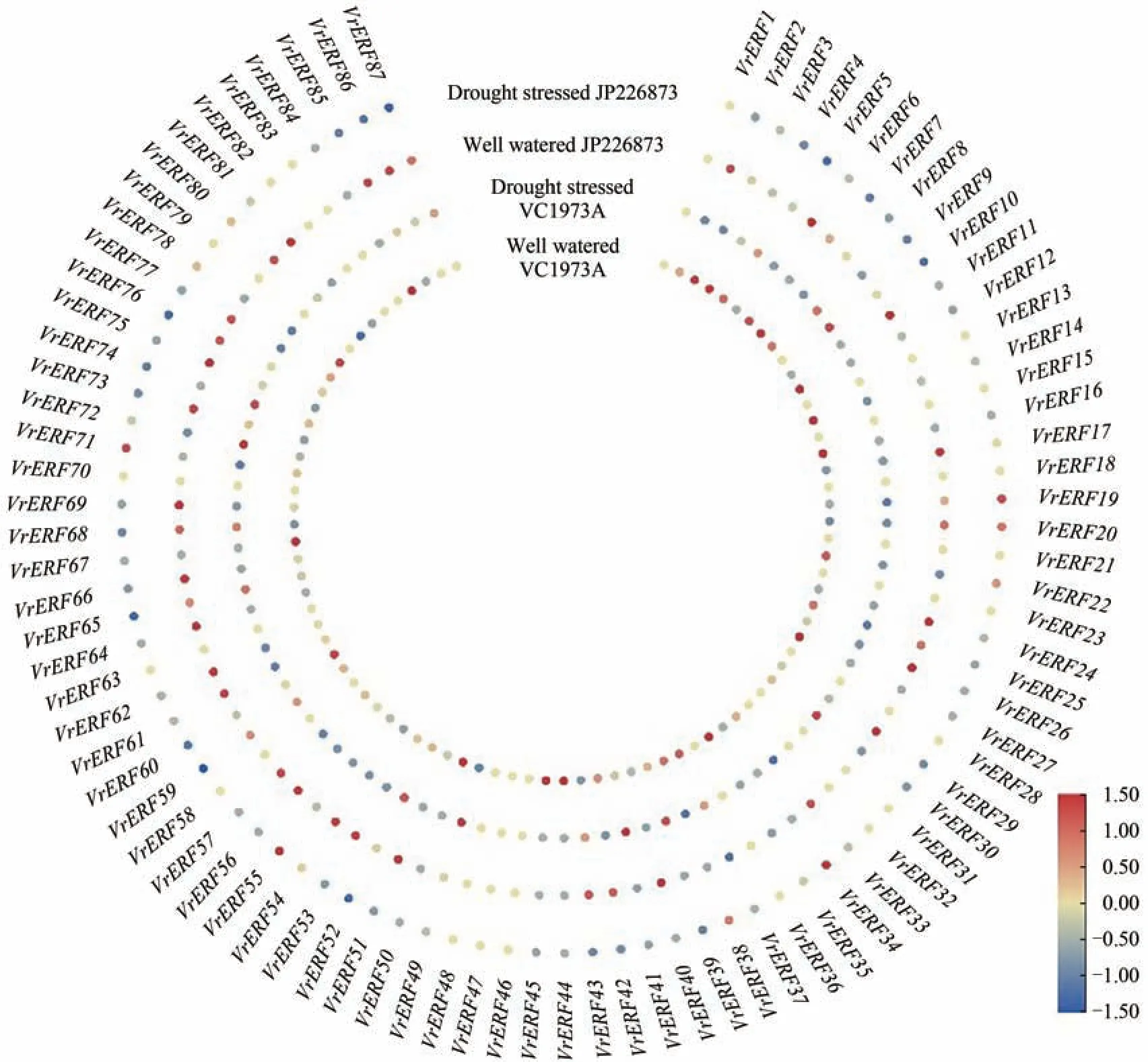
图5 正常浇水和干旱胁迫下VC1973A和JP226873中VrERF基因的表达情况Figure 5 Expression levels of VrERF in VC1973A and JP226873 under normal irrigation and drought stress
2.6 干旱胁迫下VrERF7/62表达验证分析
根据RNA-seq 构建的绿豆VrERF家族基因表达谱,挑选了干旱胁迫下在绿豆栽培种(VC1973A)和野生型(JP226873)叶片中均显著下调表达的2个基因(VrERF7和VrERF62),采用qRT-PCR 对Vr-ERF7/62在干旱胁迫处理下的表达情况进行了验证分析(图6)。结果显示,随着干旱胁迫处理时间的增加,绿豆栽培种(VC1973A)和野生型(JP226873)叶片中VrERF7和VrERF62的表达量均呈显著下降趋势。与VrERF7基因在VC1973A叶片中的表达量变化相比,它在JP226873叶片中的表达量变化呈现出更显著的下降趋势。在同品种绿豆中,VrERF62表达下降趋势较VrERF7更显著。干旱胁迫下,Vr-ERF7/62基因表达的qRT-PCR 验证分析结果总体上与转录组检测结果一致。
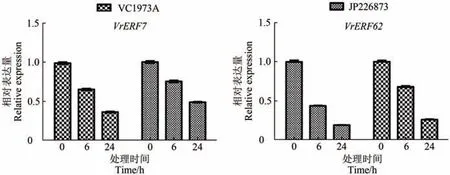
图6 VrERF7/62在干旱胁迫处理VC1973A和JP226873中表达的qRT-PCR分析Figure 6 Expression of VrERF7/62 in VC1973A and JP226873 under different abiotic stress
3 讨论与结论
AP2/ERF基因家族是植物最大的转录因子家族之一,在生长发育和逆境胁迫响应中扮演重要的角色。为筛选绿豆旱胁迫相关的AP2/ERF 转录因子,基于生物信息学、转录组测序及qRT-PCR分析,本研究初步筛选了可能与旱胁迫相关的VrERF家族基因。自Y. Sakuma等[13]在2002年从拟南芥中鉴定出145 个AP2/ERF 转录因子开始,目前已在水稻[14]、大豆[15]和小麦[16]等植物中分别鉴定出了170、148及565个AP2/ERF转录因子,不同物种AP2/ERF家族基因具有相似的结构并在进化上可能具有共同的起源。绿豆与豌豆、菜豆和大豆ERF家族基因间的共线性分析发现,PsERF、PvERF和GmERF分别与VrERF家族基因间至少存在62个同源基因对,这说明豆科植物ERF家族基因具有相同的进化起源。同时还发现豆科植物ERF的同源基因对间存在多对一现象,这也表明豆科植物ERF家族基因存在数量扩张现象,特别是多倍体大豆。
启动子区顺式作用元件是基因功能的重要组分,能够反映基因潜在的功能和调控途径[17]。绿豆AP2/ERF家族基因启动子区的顺式作用元件主要由光响应、激素响应及胁迫响应等类型组成,不同VrERF基因启动子区的响应元件数量及类型不同,这表明不同亚家族基因在功能上产生了分化。但不同亚家族基因的启动子区可能含有同类型的响应元件,它们也可能共同参与了相同的调控途径[18]。同时,构建的VrERF蛋白互作网络显示,绿豆、拟南芥等不同植物的AP2/ERF 亚家族蛋白间存在广泛的互作关系,这也进一步说明不同亚家族的AP2/ERF 转录因子可能参与同一调控途径[17-19]。AP2/ERF 家族转录因子应对干旱胁迫时具有多种调节方式,但主要通过激素调节,如在水稻中异源表达番茄TSRF1,诱导水稻脱落酸(ABA)合成相关基因SDR以及脯氨酸合成和光合作用相关基因的表达,从而增强转基因水稻对渗透胁迫和干旱胁迫的抗性[20-21]。在绿豆基因组中,VrERF家族基因的启动子序列中不仅存在脱落酸、赤霉素等激素响应元件,还具有一些干旱响应元件,这说明绿豆VrERF基因可能通过多种调节方式来调控绿豆应对干旱胁迫。
AP2/ERF家族基因具有多样的生物学功能,不同亚家族基因在植物生长发育和逆境胁迫响应中发挥不同作用。不同器官组织的特异性表达分析显示,大多数VrERF家族基因在绿豆根、下胚轴和叶等器官组织中普遍都有表达,但各VrERF家族基因在不同组织器官中的表达水平存在差异,如VrERF61下胚轴中特异性的高水平表达,而VrERF7在所有组织器官中均有较高水平的表达,各VrERF基因表达模式的分化也表明了其功能上的分化。现已有研究表明,AP2/ERF 转录因子也可作为响应干旱负向调控基因发挥作用,水稻OsERF109基因可通过抑制乙烯的释放进而对水稻的抗旱性起负调控作用,蒙古冰草AmERF4-2在干旱胁迫下呈现下调表达并呈现负调控作用[22-23]。本研究的干旱胁迫RNA-seq表达谱分析显示,与其他VrERF家族基因相比,VrERF7/62基因在干旱胁迫下呈现出显著的下调表达,qRT-PCR 验证分析也表明VrERF7/62明显受干旱胁迫诱导下调表达。同时我们还发现,VrERF7启动子区具有脱落酸响应元件,VrERF7 互作蛋白RAP2.2为乙烯响应蛋白;VrERF62启动子区具有脱落酸和干旱胁迫响应元件,VrERF62 互作蛋白RAP2.4 为干旱响应蛋白。因此,我们推测VrERF7/62参与了绿豆响应干旱胁迫过程,可能发挥了负向调控作用,但具体基因功能仍需开展后续实验进行验证。
猜你喜欢
华人时刊(2023年1期)2023-03-14
中学生天地(A版)(2023年1期)2023-02-17
汉字汉语研究(2021年2期)2021-08-30
科学大众(2020年23期)2021-01-18
汉字汉语研究(2019年2期)2019-08-27
华人时刊(2018年15期)2018-11-18
启蒙(3-7岁)(2018年8期)2018-08-13
生命科学研究(2018年1期)2018-05-29
上海农业学报(2017年3期)2017-04-10
山东农业工程学院学报(2016年6期)2016-12-01
