
葡萄枝堆肥腐殖化的微生物养分限制机理探讨
2023-05-15 06:27齐熙平黄懿梅贾鹏辉刘一帆谢健宇傅彬欣李建波
农业环境科学学报 2023年4期
齐熙平,黄懿梅,贾鹏辉,刘一帆,谢健宇,傅彬欣,李建波
(西北农林科技大学资源环境学院,农业农村部西北植物营养与农业环境重点实验室,陕西 杨凌 712100)
我国的葡萄种植面积已达8.7 亿hm2,每年冬季修剪下的枝条可达604 万t[1],这些枝条含有丰富的生物质能源、常量元素和微量元素[2],其经过堆肥化处理可转化为含腐殖质的有机肥料或土壤调理剂,用于提高土壤肥力[3]。但葡萄枝含有大量难以降解的木质素和纤维素,致使堆肥时间长、质量低[4]。高温预处理可以破坏物料中木质纤维素的结构,增加堆肥中挥发性脂肪酸、氨基酸、多糖等营养物质的含量[5-6],促进放线菌富集以及醌类再生和缩合,增强胡敏酸聚合[7];菇渣具有相对较高的比表面积、发达的微孔结构,含有丰富的腐殖质前体物质和胞外酶,添加至堆肥中有利于有机物的降解和腐殖质的形成[8],Meng等[9]的研究表明,添加15%菇渣处理使污泥堆肥中胡敏酸含量增幅显著高于未添加菇渣处理。
好氧堆肥实质上是一个由微生物主导的有机物分解并生成稳定腐殖物质的过程[10],微生物分泌的多种胞外酶可以分解复杂底物中最具限制性的养分,将高分子量有机化合物降解成可同化的分子从而促进系统中微生物养分循环和能量流动[11]。最常见的参与C、N、P循环的相关胞外酶包括β-1,4-葡萄糖苷酶(BG)、β-1,4-N-乙酰基氨基葡萄糖苷酶(NAG)、亮氨酸氨基多肽酶(LAP)和碱性磷酸酶(AP)[12]。胞外酶化学计量比,即与C、N、P 等营养物质获取相关的胞外酶比率,可以反映微生物的营养需求和对有限资源的可用性[11]。Wang 等[12]发现添加沼渣和生物炭的稻草堆肥中参与C、N 和P 转化的4 种胞外酶的活性都显著提高,有机质分解加快,BG∶AP和(NAG+LAP)∶AP 比值相比对照显著降低,微生物对P 的需求增加,受到P 的限制,微生物会分泌更多的碱性磷酸酶来维持内稳态。然而,从胞外酶活性及其化学计量变化的微生物养分限制的角度探讨堆肥过程中影响腐植酸形成的机理研究还较少,葡萄枝中添加菇渣和使用高温预处理之后的堆肥腐殖化效果及其微生物养分限制机理尚不清楚。
因此,本研究以葡萄冬剪枝条为试材,鸡粪为调理剂,探讨添加菇渣以及高温预处理对堆肥过程中理化参数、腐植酸含量、胞外酶活性及其化学计量变化的影响,揭示堆肥过程中微生物胞外酶活性及其化学计量与腐植酸形成之间的关系,为丰富堆肥腐殖化机理和促进葡萄枝的高效肥料化利用提供理论依据和数据支撑。
1 材料与方法
1.1 堆肥材料及其基本性质
新鲜鸡粪取自杨凌永顺养鸡场;葡萄枝和香菇菇渣分别来自西北农林科技大学葡萄酒学院葡萄教学标本圃和陕西省杨凌知君菌业科技公司,使用前将其粉碎至2~3 cm。物料基本理化性质见表1。

表1 堆肥原料的理化性质Table 1 Physicochemical properties of compost raw materials
1.2 堆肥方法及样品采集
试验共设3 个处理:对照组(CK),葡萄枝与新鲜鸡粪按鲜质量比约1∶1 混合后添加适量水,调节堆肥初始含水率为50%~60%,C/N 为30∶1~35∶1,总质量18 kg;添加菇渣组(M),在CK 基础上添加15%(湿质量)的菇渣;高温预处理组(H),按照CK 的比例,将原料混合加热至90 ℃且持续4 h 后,转入堆肥反应器堆肥。3组物料混合均匀后置于3个60 L密闭强制通风好氧发酵装置中堆肥45 d,装置外使用保温棉进行保温以减少热量损失。采用机械强制间歇曝气,每1 h曝气5 min,曝气速率为73 L·min-1。堆制期间不补充水分。每日9:00 和17:00 用电子温度计于堆体上、中、下部测定温度(T),取其平均值作为当日的堆温,同时测定环境温度。根据温度变化分别在第0、7、14、21、28、35、44 天取样,取样后手动翻堆,分别于堆肥物料上、中、下部采集样品,每份样品约200 g,样品混匀之后分为两份:一份于-20 ℃储存用于测定含水率、pH、电导率(EC)、种子发芽指数、溶解性有机碳(DOC)、-N、-N 以及胞外酶活性;另一份风干后用球磨仪研磨,过1 mm 筛,用于测定总有机碳(TOC)、总氮(TN)、总磷(TP)、腐植酸碳及其组分。
1.3 测定指标及其方法
称取新鲜样品2 g,将样品含水率折算后,按照干物质质量与液体体积比1∶10 加入相应体积的去离子水,180 r·min-1振荡1 h,过滤得浸提液。使用配有pH Intelli CAL 电极和EC Intelli CAL 电极的HQ11d电位仪测定浸提液的pH 和EC。将10 颗萝卜种子均匀散布在垫有滤纸的9 cm 培养皿中,吸取5 mL 浸提液浸润,在25 ℃避光条件下培养72 h 后测定发芽率和根长,计算种子发芽指数,种子发芽指数=(试验样种子发芽数×试验样种子平均根长)/(对照样种子发芽数×对照样种子平均根长)×100%。
含水率采用烘干法测定;TN 采用凯氏定氮法测定;TP采用钒钼黄比色法测定。DOC用TOC 仪(日本岛津TOC-L)测定。样品经KCl(2 mol·L-1)浸提稀释后采用流动分析仪测定-N 与-N[13]。有机质采用马弗炉灼烧减重法测定(550 ℃,6 h),测定值除以系数1.724 得TOC 含量[14]。腐植酸碳、胡敏酸碳(HA-C)及富里酸碳(FA-C)采用科诺诺娃法测定[15]。腐殖化率=腐植酸碳含量/总有机碳含量×100;腐殖化指数=胡敏酸碳含量/总有机碳含量×100。
胞外酶活性使用多功能酶标仪(瑞士Tecan Infinite M200 PRO 型)通过96 微孔板荧光法测定[16]。测定指标包括1种参与C 循环的酶:β-1,4-葡萄糖苷酶(BG),通过4-MUB-β-D-葡萄糖苷的释放量来表示;2种参与N循环的酶:β-1,4-N-乙酰基氨基葡萄糖苷酶(NAG)和亮氨酸氨基多肽酶(LAP),分别通过4-MUB-N-乙酰基-β-D-葡萄糖苷和L-亮氨酸-7-氨基-4-甲基香豆素盐酸盐的释放量来表示;1种参与P循环的酶:碱性磷酸酶(AP),通过4-MUB-磷酸的释放量来表示。
1.4 数据处理与分析
试验数据采用Excel 2019 进行统计与分析;采用SPSS 22.0中的单因素方差分析(One-way ANOVA)进行组间的显著性检验;采用OriginPro 2021 制图。冗余分析采用Canoco 5 完成;采用Amos 24.0 构建结构方程模型,并使用Powerpoint 2019绘制转化关系图。
2 结果与分析
2.1 堆肥温度、含水率、EC、pH、种子发芽指数的变化
堆肥过程中高温期维持时间长有助于杀灭寄生虫和病原体[17],提高堆肥效率。如图1 所示,CK、M、H分别于第3、8、4天达到高温期(>50 ℃),且高温期分别保持了19、15、19 d,符合《粪便无害化卫生要求》(GB 7959—2012)。添加菇渣推迟了堆体进入高温期的时间,其原因可能是菇渣中存留的菌丝以及某些致病菌或污染物影响了堆体土著微生物的生长[18]。
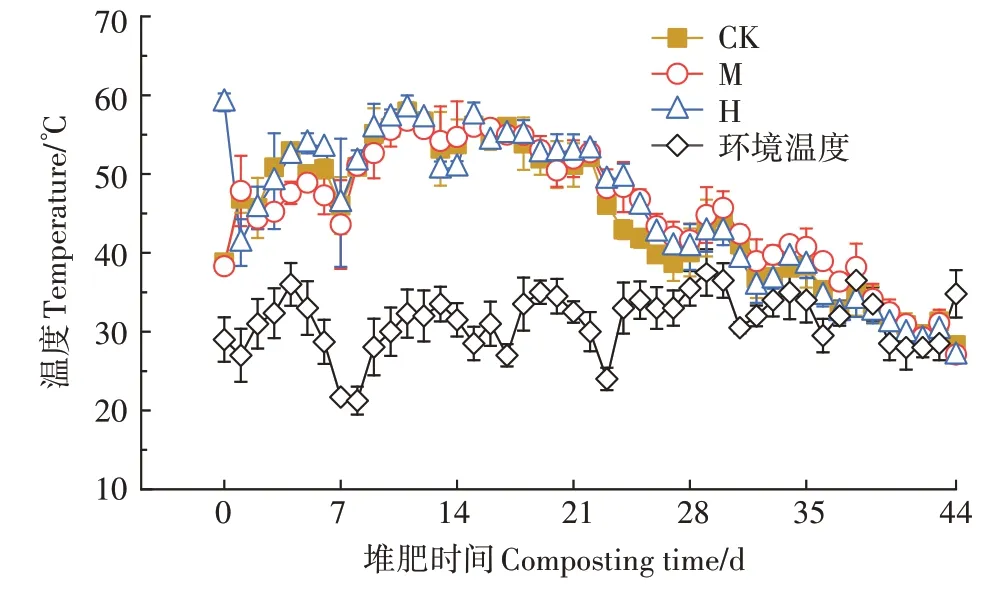
图1 堆肥过程中温度的变化Figure 1 Changes of temperature during composting
含水率变化如图2a 所示,H 的含水率降低,略低于其他两组。各组进入高温期后由于有机物降解产水,含水率稍有提升,随后逐渐下降。如图2b 所示,第7~14 天,M 和H 的EC 值显著高于CK(P<0.05),可能是相比CK,M 和H 中有机物分解产生的、挥发性脂肪酸-N和-N较多所致。14 d之后,3组的EC 值逐渐升高,可能是由于堆体水分降低引发了盐离子的富集浓缩[19]。堆肥结束时,CK、M、H 的EC值分别为3.46、3.12、3.42 mS·cm-1,差异不显著。如图2c所示,堆肥过程中3组pH呈现先升后降的趋势,这主要是由于微生物在分解有机氮过程中产生了碱性氨,14 d之后,-N 以气体的形式挥发导致pH 下降,堆肥结束时,CK、M、H 的pH 分别为8.53、8.45、8.59,差异不显著。CK、M、H 的种子发芽指数分别为103.15%、100.02%和91.00%(图2d),差异不显著,均达到70%以上,符合《有机肥料》(NY/T 525—2021)的要求。
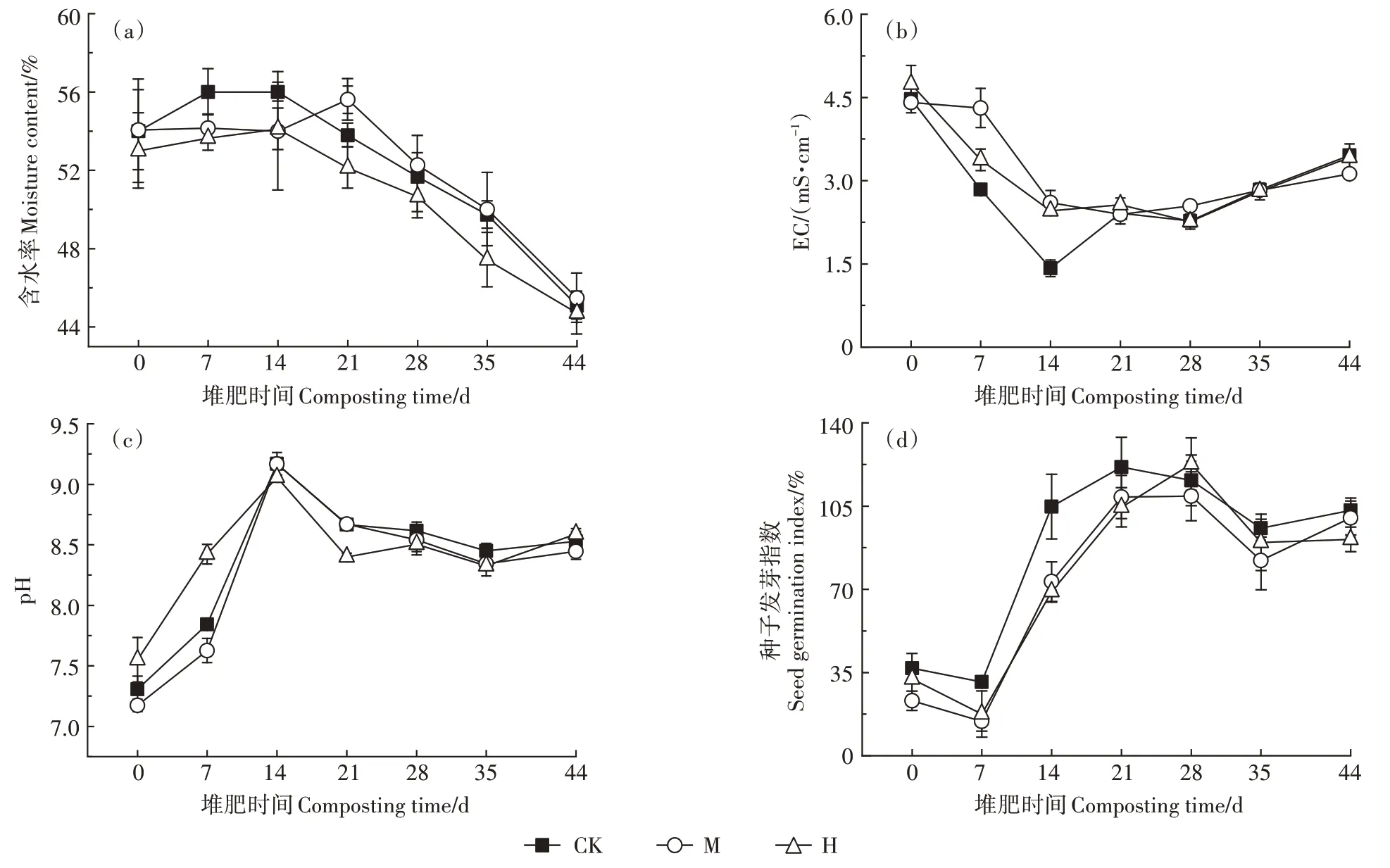
图2 堆肥过程中含水率、pH、EC、种子发芽指数的变化Figure 2 Changes of moisture content,pH,EC and seed germination index during composting
2.2 堆肥中腐植酸碳、HA-C、FA-C含量的变化
如图3a所示,第0天,H 的腐植酸碳含量较CK 高7.99%。第0~14 天,有机物降解剧烈,各组腐植酸碳含量迅速下降,之后逐渐上升。第14天和第28天,M和H 的腐植酸碳含量显著高于CK(P<0.05)。最终CK、M、H 的腐植酸碳含量分别为93.03、100.81、100.90 g·kg-1,M 和H 显著高于CK(P<0.05)。如图3b所示,第14 天,M 和H 的HA-C 含量开始同时高于CK,直至堆肥结束。总体来说,M 和H 在第14~44 天分别较CK 提高了7.64%~37.62%和9.26%~36.96%的HA-C 含量。其中M 和H 在第21~28 天的HA-C 含量较CK 增加了7.64%~37.62%和9.47%~36.96%,增速较快,这反映了M 和H 在降温阶段促进了HA-C 的形成。最终CK、M、H的HA-C含量分别为42.71、51.70、49.29 g·kg-1,M 和H 显著高于CK(P<0.05)。FA-C 的变化趋势与腐植酸碳的相似,至堆肥结束,CK、M、H的FA-C 含量分别为50.32、49.11、51.61 g·kg-1,差异不显著,与初期相比分别降低了35.77%、34.81%和40.74%。当腐殖化率≥7.0、腐殖化指数≥3.5 时堆体达到腐熟[17]。堆肥过程中腐殖化指标变化情况如表2所示,最终CK、M、H的腐殖化率和腐殖化指数分别为21.87、23.91、23.96 和10.04、12.26、11.71,均达到腐熟。相比CK,H 的腐殖化率提升了9.56%,M 的腐殖化指数提高了22.11%。
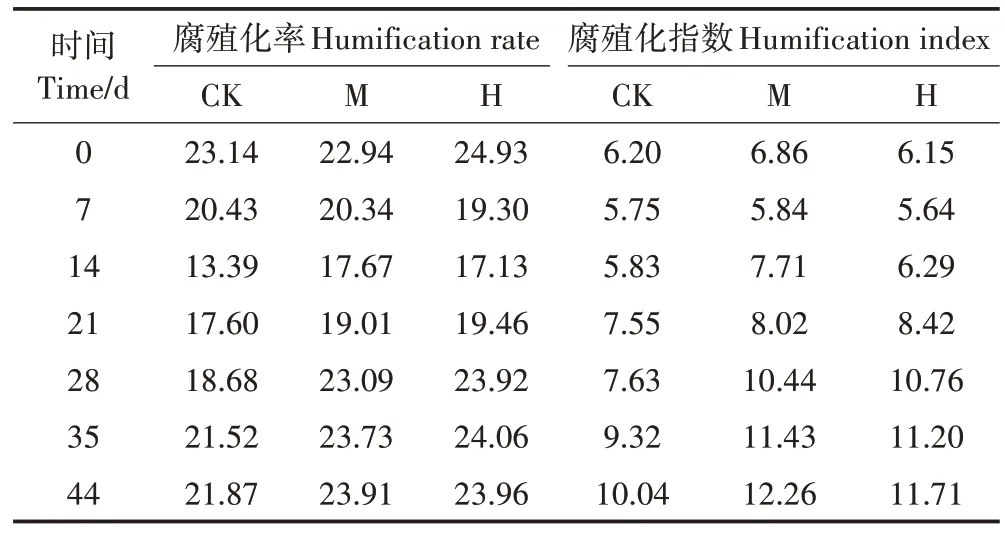
表2 堆肥过程中腐殖化指标变化情况Table 2 Changes of humification indicators during composting
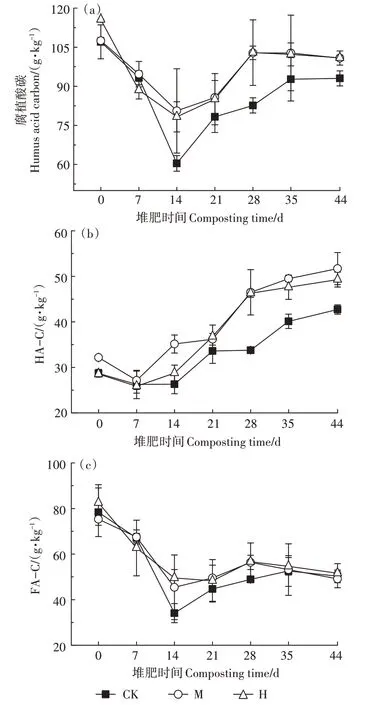
图3 堆肥过程中腐植酸碳及其组分的变化Figure 3 Changes of humus acid carbon and its components during composting
2.3 堆肥中养分及化学计量的变化
如图4a 至图4c所示,3组的TOC 含量呈下降趋势,TN 和TP含量呈上升趋势。堆肥第14~21天,H 的TOC 含量下降速率最快,为4.35%,而CK 和M 只有1.39%和1.14%。堆肥结束时,CK、M、H 的TOC 含量分别较初始下降了8.06%、10.00%、9.19%,M 和H 的TOC降解率相比CK分别提高了1.94个和1.13个百分点。CK、M、H 初始TN 含量分别为15.31、13.50、13.21 g·kg-1,M 和H 显著低于CK(P<0.05),7 d后,M 和H 的TN 含量开始高于CK。堆肥结束时,CK、M、H 的TN含量较初始分别提升了66.44%、107.28%、116.85%,M 和H 的TN 提升率比CK 分别高出40.84 个和50.41个百分点,TN 含量较CK 分别提高了9.81% 和12.44%。CK、M、H 的TP 含量较初始分别提升了54.09%、95.47%、125.49%,M 和H 的提升速率明显高于CK(P<0.05),分别高出41.38 个和70.40 个百分点,最终TP含量较CK分别提高了9.84%和9.04%。
DOC 是堆肥中微生物合成自身生命体最直接的碳源,因此容易发生微生物降解[3]。各组DOC 含量在0~14 d 内快速下降(图4d),其原因是微生物对易利用碳的快速分解利用,第14~28 天,M 和H 的DOC 含量显著高于CK(P<0.05),可能与M 和H 中难降解有机物被降解为小分子可溶性有机物有关。堆肥结束时,各组DOC 稳定在11 g·kg-1左右,差异不显著。由于有机氮的降解,各组-N 在第7 天稍有提升,之后随着堆肥的进行逐渐降低,最终稳定在0.15 g·kg-1左右(图4e),且各组间差异不显著。各组的-N含量呈降低趋势(图4f),主要是堆肥中反硝化作用使-N 转化为N2O 挥发所致,35 d 之后硝化作用增强,M 和H 的-N 含量上升,且显著高于CK(P<0.05)。
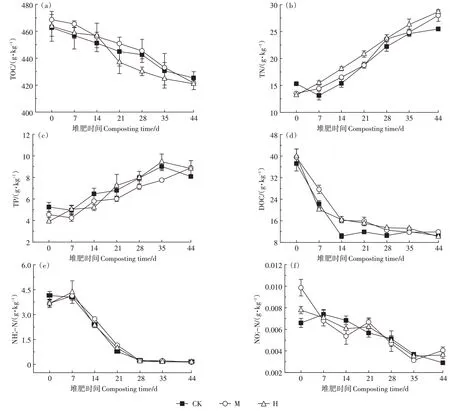
图4 堆肥过程中TOC、TN、TP、DOC、-N、-N的变化Figure 4 Changes of TOC,TN,TP,DOC,-N,-N during composting
如图5a 所示,第0 天,CK、M、H 的C∶N 分别为30.22、34.71、35.10,M 和H 显著高于CK(P<0.05),原因是菇渣本身C∶N 较高以及高温预处理期间会使一部分氮素以气体形式损失。随着堆肥进行,有机碳的降解速率高于有机氮的降解速率,C∶N逐渐降低。第14 天,CK、M、H 的C∶N 分别是29.33、27.63、25.25,H下降最快,较第0天下降了28.06%。堆肥结束时各组C∶N 稳定在15 左右,M 和H 显著低于CK(P<0.05)。如图5b 所示,第0 天,M 和H 的C∶P 显著高于CK(P<0.05),第21~35 天,M 的C∶P 高于其他两组。最终3组的C∶P 稳定在50 左右,且差异不显著。如图5c 所示,3 组的N∶P 比值从堆肥开始到结束变化较小,都在3左右,M和H的N∶P在整个堆肥期间都高于CK。
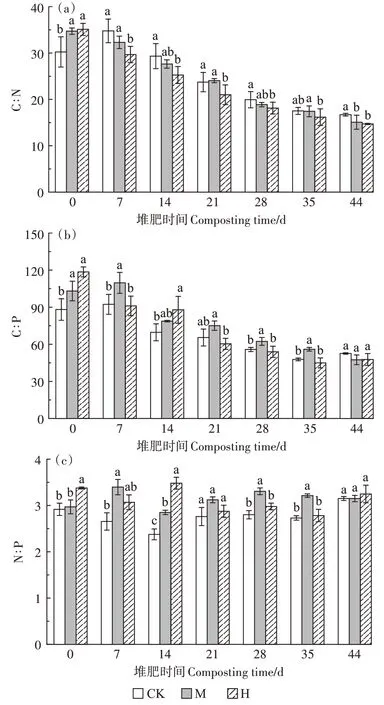
图5 堆肥过程中化学计量的变化Figure 5 Changes of stoichiometry during composting
2.4 堆肥中胞外酶活性及其化学计量的变化
BG 可将寡糖水解成单糖,为微生物提供可利用的底物和能源[20];NAG可以催化细胞壁主要成分几丁质和肽聚糖的水解[21],主要用于获取微生物容易利用的N[22]。LAP 能切割蛋白质中的肽键,在蛋白水解中起重要作用[12]。AP 是堆肥过程中参与P 循环的主要磷酸酶,可将有机磷水解成可被植物吸收和代谢的不同形式的磷酸盐[23]。
从图6可以看出,第0~7天,高温预处理导致H的各种酶活性较CK 显著降低(P<0.05),而添加菇渣可显著提升各种酶的活性(P<0.05)。第14 天,CK 和M中的AP 活性受高温影响较前期略微下降,而H 中的不降反升,并从此时到堆肥结束H 的AP 活性一直高于CK。第14天直至堆肥结束,H的BG活性显著高于其他两组(P<0.05),且在高温期时也有较大的活性(1 540.15 nmol·h-1·g-1),比CK 和M 分别高8.97%~13.07% 和9.82%~44.87%。第14天温度达到56~58℃,各组的NAG活性急剧下降,而LAP活性同时升高,CK、M、H的LAP活性较第0天分别提升了176.33%、128.48%、368.76%,M 和H 的LAP 活性显著高于CK(P<0.05)。第21 天直至堆肥结束,3 组的NAG活性都较高,除第28天外,其他时间下H的NAG活性显著高于CK(P<0.05)。第28~44 天,M 的LAP活性高于另外两组。
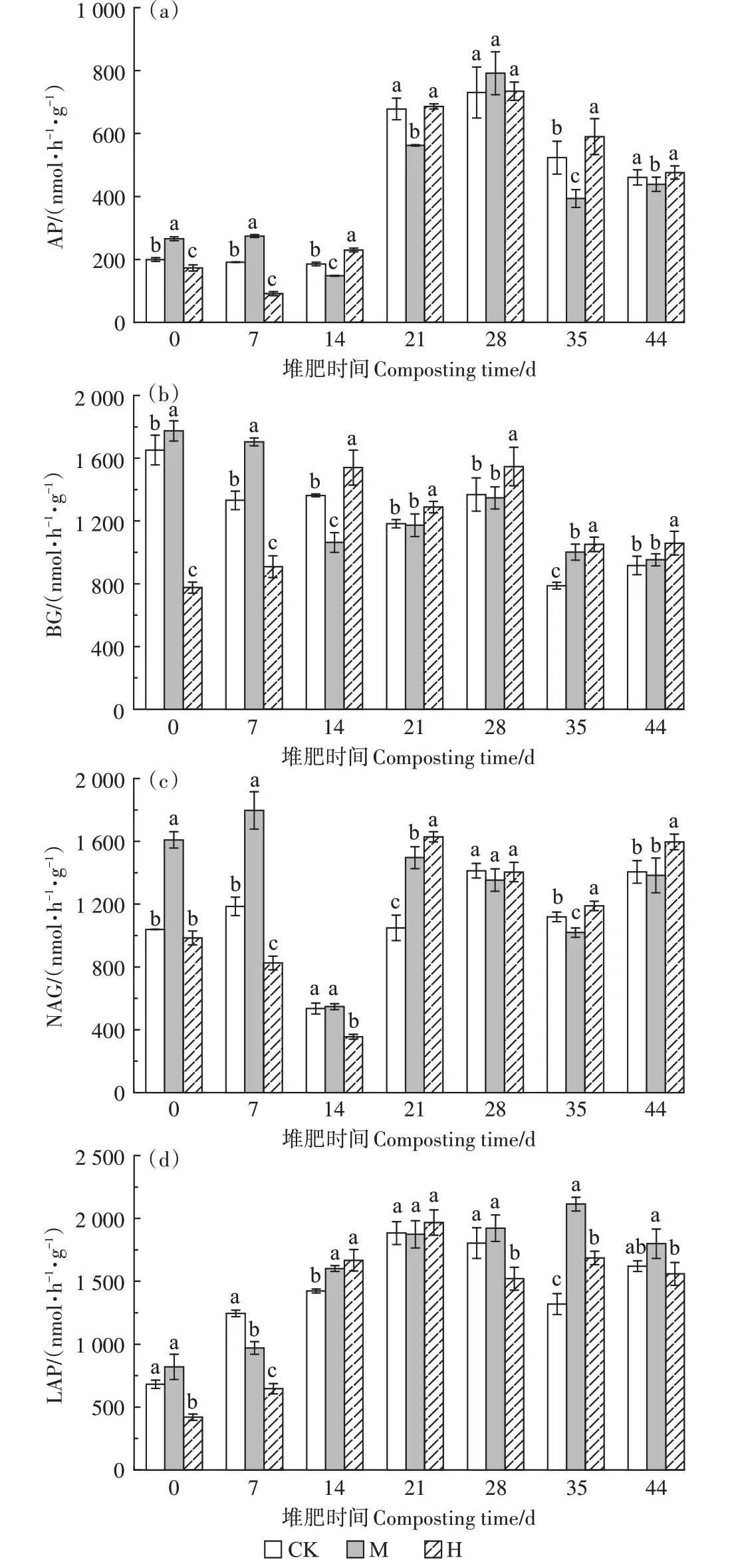
图6 堆肥过程中酶活性的变化Figure 6 Changes of enzyme activity during composting
如图7所示,堆肥第0天,CK、M、H的BG∶(NAG+LAP)比率分别为1.00、0.74、0.55,BG∶AP 比率分别为8.63、6.83、4.40,(NAG+LAP)∶AP比率分别为8.69、9.15、7.96,相比CK,M 和H中的微生物受到N和P 养分的限制。第14 天,M 的BG∶(NAG+LAP)显著低于CK,(NAG+LAP)∶AP显著高于CK,与CK相比M中的微生物受到N的限制;H的BG∶AP 和(NAG+LAP)∶AP 显著低于CK,与CK 相比H 中的微生物受到P 的限制。从第21 天开始到堆肥结束,3 组的BG∶(NAG+LAP)、BG∶AP、(NAG+LAP)∶AP 比值变化无较大起伏,第44 天,CK、M、H 的酶促C∶N∶P活性比为1.00∶3.22∶0.45、1.00∶3.22∶0.47、1.00∶2.78∶0.40,各组间无显著差异。
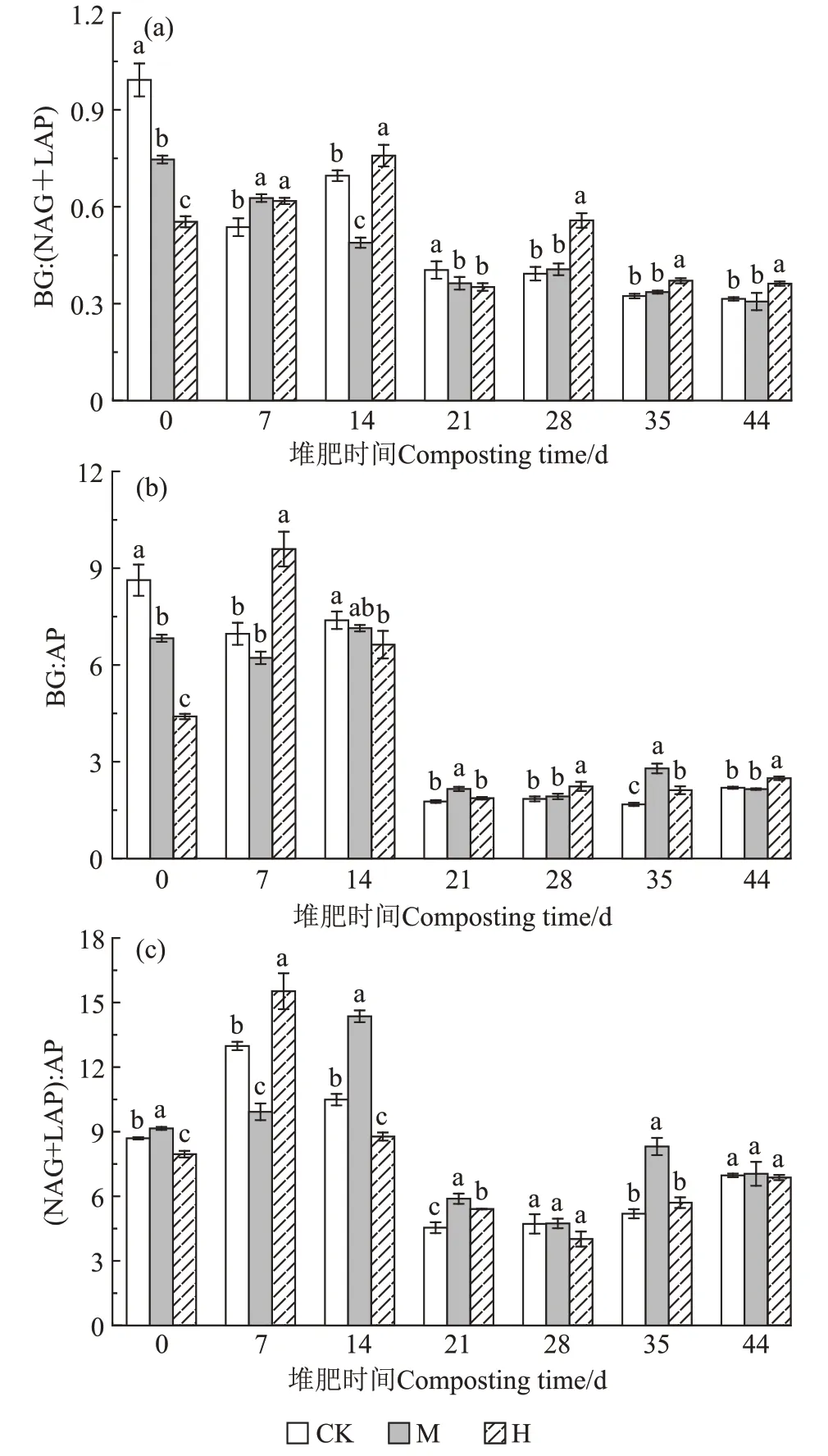
图7 酶化学计量在不同时期的变化Figure 7 Enzyme stoichiometry in different periods
2.5 腐植酸组分与理化性质、胞外酶活性之间的关系
堆肥理化性质、化学计量、胞外酶活性(蓝色箭头)和腐植酸组分(红色箭头)的冗余分析(RDA)结果如图8所示:HA-C与AP、NAG、LAP呈正相关,与TN、TP、N∶P、pH呈正相关,与FA-C、BG、DOC、TOC、C∶N、C∶P、T、EC、-N、-N呈负相关。
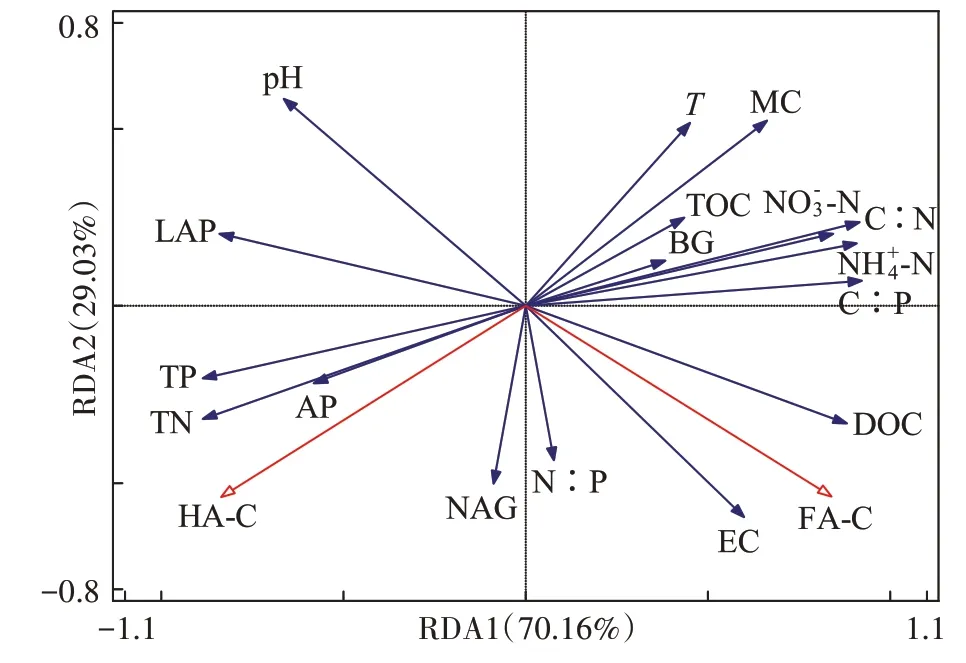
图8 堆肥过程中基本理化性质与腐植酸碳组分的冗余分析Figure 8 Redundancy analysis of basic physicochemical properties and carbon components of humic acid in composting process
根据RDA 相关性分析结果构建的结构方程模型(SEMs)如图9 所示,可以看出3 组化学计量影响相关胞外酶活性进而影响HA-C 的形成,3 个模型的拟合指数已经满足完美拟合指标的标准(X2/df<2,P>0.05,GFI>0.950,CFI>0.900 以及RMSEA<0.05)[24]。3 组影响HA-C 形成的关键胞外酶不同:在CK 中,C∶N 调节BG 和AP的活性,从而影响HA-C 的生成,C∶N、BG 和AP 都与HA-C 的生成呈负相关;在M 中,随着堆肥的进行,C∶N 逐渐降低,LAP 活性增强,促进HA-C 的生成;同理,在H 中,随C∶N 的降低,AP 活性增强,进而促进了HA-C的生成。
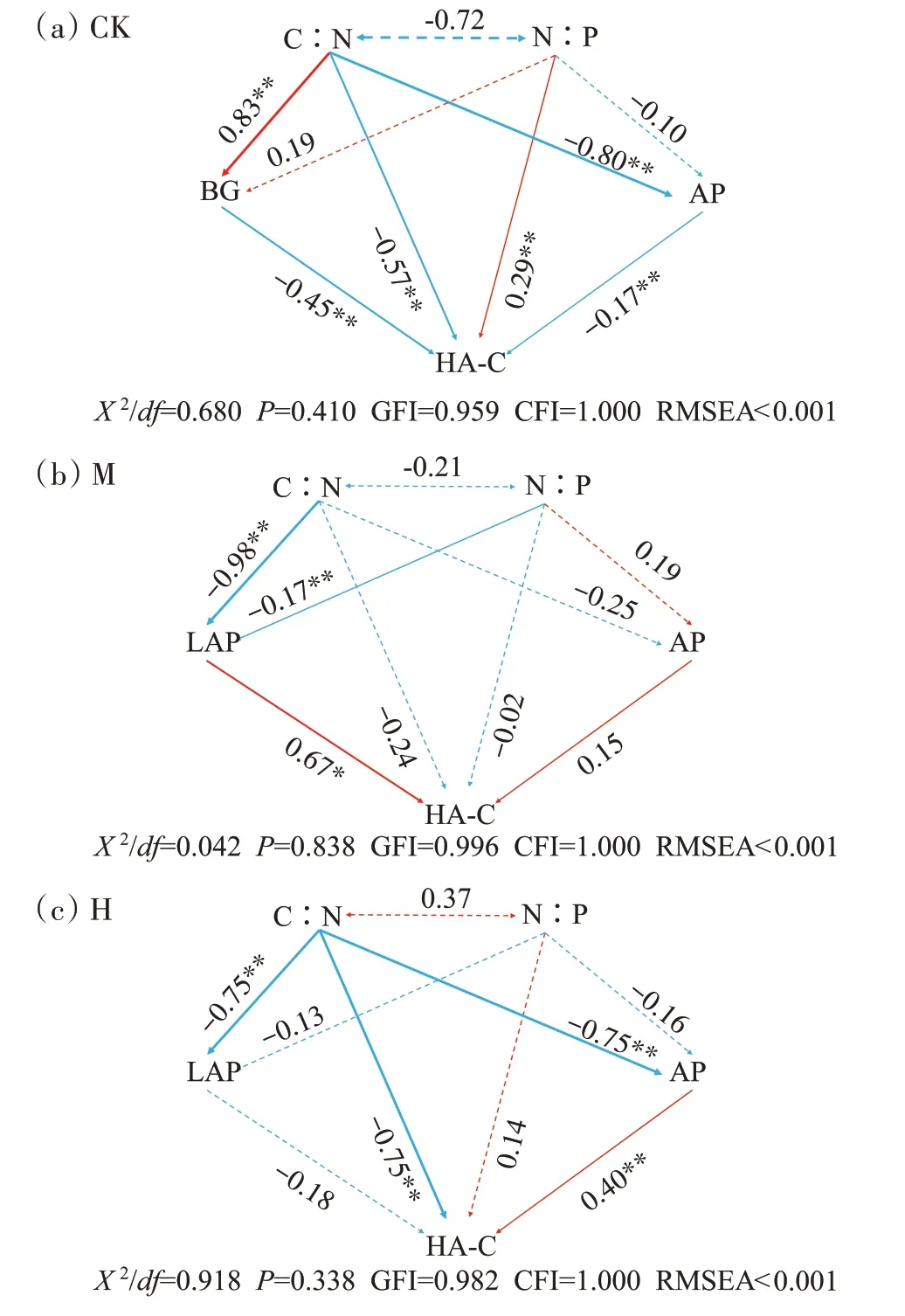
图9 化学计量和胞外酶对HA-C形成的影响Figure 9 Effects of stoichiometry and extracellular enzymes on the formation of HA-C
3 讨论
3.1 添加菇渣和高温预处理对葡萄枝堆肥腐殖化的影响
本研究3 个处理的堆肥在堆制45 d 后都达到了无害化卫生标准要求,且添加菇渣和高温预处理促进了葡萄枝堆肥胡敏酸的生成。在堆肥过程中,糖类和氨基酸等腐殖质前体可被微生物降解和转化成腐植酸,当可利用的有机物不足时,不稳定的腐植酸会被微生物降解,以提供微生物所需的能量[25]。本研究堆肥化的第7~14 天,温度较高,CK 的腐植酸碳含量陡降,下降速率为35.26%,而M和H只下降了14.89%和11.57%,且腐植酸中较稳定的组分胡敏酸开始升高,上升速率较CK 快,说明在M 和H 中一些有机碳更多地是转化为胡敏酸而不是被降解,从而提高了堆肥的腐植酸含量。高温预处理可促进堆体中多糖与类蛋白反应产生类腐殖质物质,羧基、羟基和芳核等腐殖质物质也会增多[26],因此,高温预处理初始和最终的腐植酸含量都较高。菇渣为芳构化和氧化反应提供了丰富的底物,底物中含有大量的纤维结构成分,而且菇渣本身含有较多的腐植酸[27],因此添加菇渣的堆肥中最终HA-C 含量最多。堆肥化第7 天,M 和H 的TN 就已经高于CK,说明添加菇渣和高温预处理可以减少堆肥中的N 损失,加速TN 的浓缩效应。菇渣中的易降解有机物较多,其快速分解导致较多短链脂肪酸累积[28],从而可中和一部分NH+4-N,减少N 素损失,这与Meng 等[9]在污泥堆肥中添加菇渣的研究结果一致。高温预处理会抑制反硝化细菌生长以及产生有机酸降低物料pH 而起到保N 作用[29]。研究表明,部分腐植酸的形成是通过含N 和含P 化合物与其他腐殖质前体的缩合实现的[30-31]。本试验中,TN、TP 与HA-C 的含量都是随着堆肥的进行逐渐增加,三者之间存在正相关关系(图8),说明一些有机物分解产生的N、P离子以与其他前体螯合的形式被回收,并存在于HA-C 中。TN、TP 和HA-C 的含量同时有效提升,与Jia 等[25]在菇渣堆肥中添加园林废弃物的研究结果一致。这也是添加菇渣和高温预处理可促进胡敏酸生成的原因之一。
3.2 添加菇渣和高温预处理对胞外酶活性的影响
胞外酶是微生物释放的可以降解复杂大分子的蛋白质,参与养分循环并具有重要作用,其活性强弱可以表示养分转化能力的大小[32]。添加菇渣在堆肥初期可显著提升4 种胞外酶的活性。香菇菇渣富含降解纤维素及木质素的真菌[4],可增加堆肥中有机物的含量,为真菌、细菌等微生物的生长提供了更多更易利用的碳源和氮源,从而使堆肥初期酶活性较高。M 的NAG 酶活性在堆肥初始达到峰值,可能是由于菇渣中富含真菌菌丝,微生物残余积累的肽聚糖丰富,死亡的真菌菌丝体通过微生物演替而分解[33]。由于90 ℃的高温预处理,H 的AP、BG、LAP 活性在堆肥初期显著低于其他两组(P<0.05),可能是因为超高温堆肥衍生出的芳烃比例较高,且酚基碳的强度更高[34],而酚类物质的浓度增加会导致和养分循环相关的微生物活性的降低[12]。有机物的分解速率可以用胞外酶活力来表征[32],各组高温时期只有BG 和LAP的活性没有因为高温而明显下降,可能由于此时微生物大量繁殖,纤维素、蛋白质等碳源、氮源快速分解,微生物的同化吸收更需要这两种酶参与,H 的4 种胞外酶活性在高温期和腐熟期都显著高于CK(P<0.05),表明高温预处理有助于大分子有机物的降解。各组4 种酶的活性都是在第28 天开始下降并逐渐稳定,原因是在升温和高温阶段,易代谢底物较多,微生物大量繁殖,大分子有机物迅速分解;而在后期,大部分容易利用的有机物已被降解完全,堆体稳定,微生物活性减弱,酶活性降低。此外,酶和腐殖质之间形成的复合物也破坏了酶与底物的相互作用,导致在堆肥后期胞外酶活性降低[12]。
3.3 堆肥养分和胞外酶活性的化学计量与HA-C 生成的关系
化学计量能够有效说明养分限制类型,是反映C、N、P 循环以及养分平衡的重要参数[35]。堆肥C∶N是体现堆肥有机物分解速率的指标,比值越高表明有机物的矿化分解速率越慢[16]。本研究中,从第7 天开始到堆肥结束,H 的C∶N 比其他两组都低,尤其在高温期迅速下降,下降速率相比CK 高247.41%,也是在同一时期,H 的HA-C 含量较CK 显著上升,表明高温预处理有利于有机物降解矿化和胡敏酸的快速形成。C∶P 也能在一定程度上反映有机质的矿化速率,通常C∶P 越低,越利于微生物对有机物的分解,同时提高堆肥全P 的含量[36]。与C∶N 相同,H 的C∶P 下降速率快,也显示出高温预处理有利于有机物矿化。第14天之前,M 和H 的C∶P 高于CK,第14 天之后,H 的C∶P开始低于CK,第44天,M和H的C∶P都低于CK。整个堆肥期间,M 和H 的N∶P 一直高于CK,M 在第7 天到第14 天的C∶P 显著高于其他两组(P<0.05),结合相应时期M 的BG∶AP 较小以及N∶P 较大,说明M 组微生物受环境影响需要分泌更多的AP来满足生长需求,在此期间M 中的微生物受到P养分的限制。研究证明,微生物会最大限度地优化自己的储备资源,易利用的底物是提升微生物生物量和分泌胞外酶的基础[37]。堆肥过程中,当可用的能量和营养物质变得有限时,微生物就会释放该元素的获取酶,以应对养分供应的变化,从而改变生态酶的化学计量特征[37]。SEMs 显示(图9),在整个过程中,各组的HA-C 含量受不同胞外酶的影响:CK 的HA-C 含量与BG 和AP都呈显著负相关(P<0.01)。当BG 酶分泌较多时,纤维素和木质素逐渐被分解,而它们的类衍生物是构成腐植酸的核心骨架和主要成分[17],因此不利于胡敏酸聚合。M 的HA-C 含量与LAP 呈显著正相关(P<0.05),在H 中HA-C 含量和AP 呈显著正相关(P<0.01)。即在M 和H 中产生的微生物N 和P 限制会提高蛋白质、有机磷降解酶的活性,导致N 和P 矿化增加,以应对N、P 限制,矿化后的物质有利于合成胡敏酸:如NH+4-N 可通过非生物途径作为前体直接参与胡敏酸的形成[38];P 和腐植酸、胡敏酸之间可以螯合,螯合产物还能促进P 的溶解度和生物利用率[31]。从图7 可以看出,BG∶(NAG+LAP)、BG∶AP、(NAG+LAP)∶AP的值都是在第21天之前变化较大,与CK相比此时M 和H 中的微生物受到的养分限制较大,第21 天之后比值减小并稳定,也是因为前期有机质降解,组分变化快,难以降解的大分子有机物需要微生物分泌相应的胞外酶来降解。
4 结论
(1)葡萄枝鸡粪好氧堆肥高温期(>50 ℃)持续10 d 以上,种子发芽指数均达到70%以上,满足堆肥无害化和腐熟标准。添加菇渣(M)和高温预处理(H)改变了堆肥基质的特性,加速了有机质降解,提升了养分含量,促进了腐殖化。堆肥第44 天,M 和H 的TN提升率相比CK 提高了40.84 个和50.41 个百分点,TP含量提升率相比CK 提高了41.38 个和71.40 个百分点、TOC 含量的降解率分别比CK 提高了1.94 个和1.13个百分点;相比CK,H 的腐殖化率提升了9.56%,M的腐殖化指数提升了22.11%。
(2)添加菇渣和高温预处理影响了参与C、N、P循环功能的酶的活性和化学计量,显著提升了4 种胞外酶的活性:M 组在第0~7 天胞外酶活性较高,H 组在第14~44 天胞外酶活性较高,以应对N 和P 养分的限制,降解更多的大分子有机物,提升N 和P 的含量,有利于胡敏酸的生成,进而提升了胡敏酸的含量。
猜你喜欢
腐植酸(2022年1期)2022-03-03
江苏农业科学(2020年2期)2020-04-17
腐植酸(2020年1期)2020-03-04
广东教育·高中(2017年11期)2017-12-04
江苏农业科学(2017年10期)2017-07-21
湖北农业科学(2015年17期)2015-10-09
腐植酸(2013年1期)2013-09-25
环境与可持续发展(2013年3期)2013-03-11
植物营养与肥料学报(2012年5期)2012-10-26
现代营销·经营版(2010年5期)2010-05-14
