
四川省育成小麦品种和高代品系对条锈病的抗病性评价
2023-06-05 04:43郎晓威王凤涛姬红丽蔺瑞明冯晶曹世勤姚小波
植物保护 2023年3期
郎晓威 王凤涛 姬红丽 蔺瑞明 冯晶 曹世勤 姚小波


摘要
四川省不僅是条锈菌冬繁区,也是我国东部麦区重要的春季初侵染菌源地和毒性变异地。准确评价四川小麦新育成品种和高代品系对条锈病的抗性水平,能为合理培育和使用抗病品种提供重要依据。利用当前条锈菌优势小种和4个地点的成株期鉴定圃对来自四川省的115份小麦育成品种和299份高代品系进行苗期和成株期抗病性鉴定。结果表明,新育成品种苗期对优势小种的高抗CYR32、CYR33和CYR34的比例发生不同程度的变化,对CYR32的高抗比例为40.9%、对CYR34的高抗比例为17.4%,而供试高代品系高抗比例从52.3%降至46.2%;中抗类型育成品种从20.0%增加到40.9%,高代品系从19.6%增加至33.2%。表明条锈菌优势小种更替后一些材料仍保留了部分抗性,后期选育的高代品系兼顾对新小种的抗病性。育成品种在四川成都和甘肃清水成株期鉴定,高抗品种所占比例较低,分别为13.9%和3.5%,但高代品系高抗类型比例较高,分别为42.2%和8.5%,表明甘肃和四川条锈菌的致病类型和发病环境条件存在较大差异,近年来四川小麦成株抗条锈病育种取得显著进展。
关键词
小麦条锈病; 抗病性评价; 全生育期抗性; 成株抗性; 基因聚合
中图分类号:
S 435.121
文献标识码: A
DOI: 10.16688/j.zwbh.2022039
Disease resistance evaluation of wheat commercial cultivars and advanced breeding lines developed in Sichuan to stripe rust
LANG Xiaowei1,2,3, WANG Fengtao2,5, JI Hongli4, LIN Ruiming2,5, FENG Jing2,5,CAO Shiqin1,3*, YAO Xiaobo6*
(1. College of Plant Protection, Gansu Agricultural University, Lanzhou 730070, China; 2. State Key Laboratory for
Biology of Plant Diseases and Insect Pests, Institute of Plant Protection, Chinese Academy of Agricultural Sciences,
Beijing 100193, China; 3. Institute of Wheat Research, Gansu Academy of Agricultural Sciences, Lanzhou 730070,
China; 4. Key Laboratory of Integrated Management of Pest on Crops in Southwest China, Ministry of Agriculture and
Rural Affairs, Institute of Plant Protection, Sichuan Academy of Agricultural Sciences, Chendu 610066, China;
5. National Agricultural Experimental Station for Plant Protection in Gangu, Tianshui 741000, China;
6. Agricultural Research Institute, Tibet Academy of Agriculture and Animal Husbandry Sciences, Lhasa 850032, China)
Abstract
There is an overwintering region in Sichuan province for the pathogen Puccinia striiformis f.sp. tritici (Pst), which also serves as an important spring Pst inoculum source of the East wheat area and virulence variation center. Through accurate evaluation of disease resistance of commercial wheat cultivars and advanced breeding lines developed in Sichuan, valuable data will be presented for rational development and deployment of resistant wheat cultivars to stripe rust. Resistance of 115 commercial cultivars and 299 high breeding lines inoculated with three predominant Pst races at seedling stage and adult stage was evaluated in four field nurseries. The results showed that there were different proportions of highly resistant types among commercial cultivars at seedling stage to the predominant races CYR32, CYR33, and CYR34. The high resistance proportion to CYR32 was 40.9% and 17.4% to CYR34, while among advanced breeding lines, highly resistant type proportions reduced from 52.3% to 46.2%. The percentages of the moderately resistant type of commercial cultivars increased from 20.0% to 40.9%, while 19.6% up to 33.2% for the advanced breeding lines. The results indicated that some of the materials remained partial resistance after the predominant races shifted, and later the resistance to the emergence of the new races was integrated into advanced breeding lines. The highly resistant cultivars at the adultplant stage accounted for a relative low percentage in the field nurseries at Chengdu, Sichuan (13.9%) and Qingshui, Gansu (3.5%), but 42.2% and 8.5% of advanced breeding lines at the two locations, showing the great differences in the pathotypes and environmental conditions for pathogen infection between Gansu and Sichuan. In recent years, significant progresses have been made in wheat adultplant stripe rust resistance breeding in Sichuan.
Key words
wheat stripe (yellow) rust; disease resistance evaluation; allstage resistance; adultplant resistance; gene pyramiding
由條形柄锈菌小麦专化型Puccinia striiformis f.sp. tritici (Pst)引起的条锈病是我国小麦生产上的主要气传病害,一般流行年份造成减产约5%~10%[13],各小麦种植区内每年均有不同程度条锈病流行危害。根据条锈菌遗传多样性和有性杂交情况,推测Pst可能起源于喜马拉雅山脉周边高海拔冷凉区域[4],毗邻该区域的中国境内分布着世界最大且相对独立的条锈病流行区[5],几次大规模条锈病暴发给我国造成了严重经济损失[67]。四川省是条锈病常发流行区域,1999年-2002年连续4年条锈病发生面积均超过47万hm2,每年小麦减产约30万t,特别是2002年条锈病发病面积超过100万hm2,造成四川小麦减产超过40万t[8]。此外,四川省西北部高海拔地区和成都平原是重要的条锈病菌源地和毒性变异区[1, 911],该区域也是条锈菌最重要的冬繁区,为东部长江中下游和西北冬麦区提供春季流行菌源[6]。持续、有效防控条锈病对保障四川省及其周边小麦稳产高产具有重要意义[1]。
种植抗病品种是防控条锈病最经济、有效和环保的措施[3, 12]。但是,条锈菌群体具有丰富的遗传变异特性,群体数量庞大;同时生产上对主要抗条锈病基因过度单一使用,品种间抗病遗传背景高度相似,加大了对病原菌致病性变异的定向选择压力。新生理小种和致病类型相继出现和优势小种轮替,克服了主栽品种中已广泛应用的抗病基因,品种大面积应用3~5年后就成为感病品种,再次引发病害大流行[1315]。例如,全生育期抗病基因Yr26在主栽品种中广泛应用几年后,导致CYR34上升为毒性最强的优势流行小种,造成含有Yr10和Yr26的川麦系、小偃系、兰天系等众多品种丧失抗病性[9, 11, 16]。因此,只有不断发掘和合理利用抗病新基因资源才能为抗病育种和有效防控条锈病提供技术保障。
随着小麦条锈菌新致病类型的出现,许多抗病主栽品种的抗性逐渐降低甚至完全被克服[17]。四川省非常重视抗病品种培育,近年来培育出一批新的小麦品种。为了弄清新育成品种和高代品系的抗病性现状,本研究将对四川省育成品种和高代品系进行苗期和成株期抗条锈性鉴定,以明确这些材料的抗性水平及抗性类型,为发掘抗病新基因和拓宽小麦抗条锈病遗传资源提供材料,为实现小麦抗病基因合理布局提供技术支持。
1 材料与方法
1.1 供试材料
供试314份材料来自四川省小麦育种单位,其中已审定登记的育成品种115份,高代品系199份,‘铭贤169作为感病对照品种。小麦条锈菌优势生理小种CYR32、CYR33和CYR34作为苗期抗病性鉴定小种,CYR32、CYR33和CYR34及Hybrid 46类群、水源11类群和G22类群的主要致病类型混合菌株进行成株期抗性鉴定或田间气传自然菌株接种。
1.2 试验方法
1.2.1 苗期鉴定
待麦苗第2片叶完全展开后,利用电子氟化液(NovecTM 7100)分别将CYR32、CYR33和CYR34新鲜夏孢子等量混匀配制成4 mg/mL孢子悬浮液喷雾接种,每盆材料15株大约接种1 mg夏孢子,接种后置于10℃接种间黑暗保湿24 h,再转到人工气候室(昼15~18 ℃/夜11~14 ℃;光照时间12~14 h/d,光强5 000~6 000 lx)继续潜育培养[18]。
1.2.2 成株期鉴定
2018年-2020年在甘肃省天水市清水县、湖北省荆州市、河北省廊坊市和2020年在四川省成都市郫都区设置抗性鉴定圃,进行供试材料的成株期抗条锈性表型鉴定。鉴定圃行长3 m,行距0.4 m,采用穴播方式,每行播种5个品种/品系。每个品种/品系播种15~20粒种子,均匀分散在约10 cm范围内。每间隔20行鉴定材料播种1行感病对照品种‘铭贤169。鉴定圃周边播种感病品种‘铭贤169作为接种诱发行。在小麦分蘖初期,对清水县、廊坊市和郫都区鉴定圃的诱发行‘铭贤169喷雾接种混合菌株的孢子悬浮液,位于荆州市的鉴定圃利用气传菌源自然接种。
1.2.3 侵染型调查
在苗期和成株期,均采用0至4级侵染型分级标准[19],0级:叶上不产生任何可见的症状,0;级:叶上产生小型枯死斑,不产生夏孢子堆;1级:叶上产生枯死条点或条斑,夏孢子堆很小,数目很少;2级:夏孢子堆小到中等大小,较少,周围叶组织枯死或显著褪绿;3级:夏孢子堆较大、较多,其周围叶组织有褪绿现象;4级:夏孢子堆大而多,周围不褪绿。其中0为免疫,0;为近免疫,1级为高抗(HR),2级为中抗(MR),3级为中感(MS),4级为高感(HS)。苗期接种后约15 d,当感病对照品种充分发病后进行第1次侵染型调查,间隔4 d进行第2次调查。在成株期,待感病对照品种发病平均严重度达80%以上开始侵染型调查,间隔7 d进行第2次调查。综合2次调查结果确定待鉴定品种的苗期和成株期抗条锈病表型。苗期抗锈病鉴定试验独立重复2次。
1.3 数据统计方法
利用软件WPS Office 2019统计分析小麦抗条锈病品系在苗期和成株期不同抗性类型的比例。
2 结果与分析
2.1 苗期抗性鉴定结果
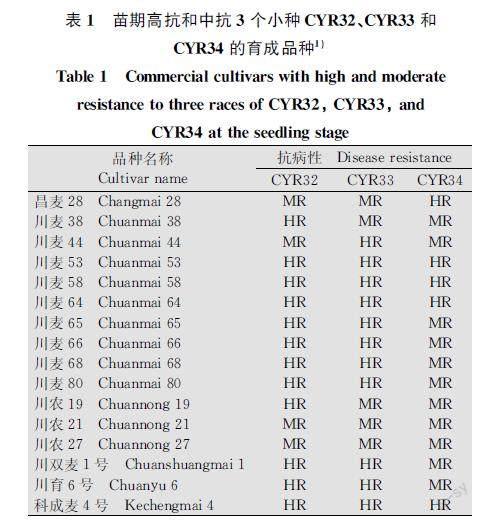
分别利用3个优势小种对115份育成品种进行苗期抗性鉴定,其中‘西科麦1号‘川农19‘川麦80等47个品种高抗CYR32,占40.9%;‘昌麦28‘绵阳32‘蜀麦1613等23个品种中抗CYR32,占20.0%;‘川育6号‘川麦44‘绵麦37等36个品种高抗CYR33,占31.3%;‘川农21‘西科麦1号‘川麦38等34个品种中抗CYR33,占29.6%;‘昌麦28‘川麦53‘中科麦138等20个品种高抗CYR34,占17.4%;‘内麦8号‘绵阳32‘川麦38等47个品种中抗CYR34,占40.9%。另外,‘资麦1号‘中科麦138‘川育6号等27个育成品种高抗CYR32和CYR33;‘资麦1号‘川麦53‘西科麦1号等11个品种高抗CYR32和CYR34;‘绵麦37‘良麦2号‘川麦64等10个品种高抗CYR33和CYR34。在所鉴定的材料中,苗期高抗3个小种的育成品种仅有‘资麦1号‘川麦53‘中科麦138等9个品种,对3个小种中抗以上的包括‘川麦38‘绵阳32‘西科麦1号等33个品种(图1,表1)。
另外,鉴定了199份高代品系苗期分别对3个优势小种的抗病性,其中‘15S15135‘15S16339‘15S18332等104个品种高抗CYR32,占52.3%;‘CD011J33‘CD01512372‘CD01515689等39个品种中抗CYR32,占19.6%;‘15S15135‘15S22971‘RC9等90个品种高抗CYR33,占45.2%;‘CD01515439‘CD01518298‘RC34等48个品种中抗CYR33,占24.1%;‘G1518‘G1520‘PZ1510等92个品种高抗CYR34,占46.2%;‘Y15211‘CD011J33‘CD01513002等66个品种中抗CYR34,占33.2%(图1)。
其中,部分高代品系同时抗2个小种,如114个品系对CYR32和CYR33两个小种均达到中抗以上,119个品系对CYR32和CYR34均达到中抗以上,112个品系对CYR33和CYR34小种均达到中抗以上。
另外,‘G1514‘G1520‘蜀麦1609等33个品系苗期高抗3个小种,‘RC7‘RC8‘科成麦7号等62个品系苗期中抗3个小种。
根据苗期抗性鉴定结果,发现育成品种高抗CYR32、CYR33和CYR34的比例分别为40.9%、31.3%、17.4%,对3个小种的高抗比例逐渐下降;但中抗品种所占比例从CYR32的20.0%增加到CYR34的40.9%;高代品系中高抗上述3个优势小种的比例从CYR32的52.3%降低至CYR34的46.2%,中抗品系比例从19.6%增加至33.2%(图1)。比较育成品种与高代品系分别对3个优势小种表现高抗的比例变化,高抗CYR32、CYR33和CYR34的育成品种所占比例减少幅度较大,但高代品系表现高抗类型的比例变化幅度相对较小。
2.2 成株期抗性评价
在甘肃清水县鉴定圃,‘川农19‘川农21‘乐麦3号‘川麦62共4个育成品种表现为高抗,占3.5%,而‘川农17‘绵阳35‘川农18等71个品种表现为中抗,占61.7%;在四川郫都区鉴定圃,‘川农21‘乐麦3号‘科成麦6号等16个育成品种表现为高抗,占13.9%,而‘绵阳12‘绵农7‘绵阳35等75个品种表现中抗,占65.2%;在湖北荊州市鉴定圃,‘川农19‘川育19‘乐麦3号等40个育成品种表现为高抗,占34.8%,‘绵阳12‘川育9号‘绵农7等67个品种表现为中抗,占58.3%;在河北廊坊市鉴定圃,‘川育16‘西科麦1号‘川农26等50个育成品种表现为高抗,占43.5%,‘西辐12 ‘绵麦37‘内麦9号等47个品种表现为中抗,占40.9%(图2,表2)。成株期在4个鉴定圃中均表现高抗的育成品种仅有‘川农21‘乐麦3号,表现中抗以上品种包括‘绵阳31‘川麦38‘西辐12和‘中科麦138等54份育成品种。
199份高代品系在不同的鉴定圃中成株期抗性表型也存在差异。例如在甘肃清水县,‘G1522 ‘Y162756‘LW15B2976等17个品系表现为高抗,占8.5%,‘Y15211‘蜀麦1615‘CD011J33等157个品系表现为中抗,占78.9%;但在四川郫都区,‘15S20335‘CD015153310‘CD01515379等84个品系表现为高抗,占42.2%,‘15S16339‘15S22971‘蜀麦1615等101个品系表现为中抗,占50.8%(图2)。高代品系‘CD01518907‘CD01519035‘G1520‘G1522和‘PZ152在4个鉴定圃中成株期均表现为高抗,而‘RC20‘RC34和‘RC63等148份品系表现为中抗。
育成品种和高代品系在4个不同地点的鉴定圃成株期对条锈病反应型不同,例如在甘肃清水和四川成都的鉴定圃中,高抗类型育成品种分别占3.5%和13.9%,明显低于在湖北荆州(34.8%)和河北廊坊(43.5%)的比例,但在甘肃和四川两个鉴定圃的中抗育成品种数量较为接近;高代品系在四川有42.2%为高抗类型,低于在湖北(74.4%)及河北(75.9%)的比例,而明显高于在甘肃(8.5%)的比例(图2)。
依据供试品种/品系在苗期和成株期抗性表型鉴定结果,获得具有全生育期抗性和成株抗性类型的材料。‘川麦38‘川农19‘川农21等20份育成品种和‘Y15211‘CD01512681‘CD01515234等74份高代品系在苗期对3个生理小种和成株期在4个鉴定地点均为抗病表型,属于全生育期抗性类型,其中‘CD01519035‘G1520‘G1522‘PZ152为全生育期高抗类型。
在本研究中,还发现一些品种/品系在苗期为抗病表型但在成株期为感病,如‘川育6号‘川麦35‘川育16‘绵阳32等34个育成品种苗期抗优势小种,但在甘肅省清水县成株期属于中感类型;在四川郫都,‘绵阳26‘川农16‘内麦8号‘蓉麦2号等22个苗期抗病的育成品种成株期属于中感类型。
2.3 不同年份育成品种的抗性比较
90个育成品种分别在2002年-2009年及2013年-2015年间审定,依据成株期四川鉴定圃的鉴定结果,每年审定的高抗类型品种所占比例从12.5%增加到28.6%,平均为10.7%;中抗类型品种所占比例平均为68.0%。在90份育成品种中,平均每年审定高抗和中抗CYR32小种的品种所占比例分别为51.0%和18.8%,而高抗和中抗CYR34小种所占比例分别为18.0%和38.8%(图3),说明优势小种更替严重影响了抗病育种的选择方向。本研究中2016年通过审定的‘蜀麦1613‘科成麦5号‘绵麦112在苗期对3个优势小种和成株期在4个鉴定圃均表现为中抗水平以上。
3 结论与讨论
3.1 四川小麦育成品种抗条锈性变化趋势
在本研究中,四川115个育成品种中仅‘繁6和‘雅安早是20世纪70至80年代广泛种植的品种,其余113个为审定品种。在2002年-2009年及2013年-2015年期间审定的90个品种中,成株期在四川鉴定圃高抗和中抗的品种在每年审定品种中所占比例变化不明显。主要是因为病原菌的毒性变异加快,特别是2009年出现并快速上升为优势流行小种的CYR34[16, 2022],导致大量后期育成的含有抗病基因Yr10、Yr26品种丧失抗病性。在90份育成品种中,平均每年审定的高抗CYR32品种比例(51.0%)显著高于高抗新小种CYR34的比例(18.0%),但对CYR32和CYR34小种中抗的品种比例从18.8%增加到了38.8%(图3)。说明在条锈菌流行小种更替过程中,虽然大部分品种丧失了抗病性,但是其中一些品种仍保留了部分抗性,而抗性由高抗转变为中抗。因此,应重视利用抗性较为稳定的基因和多基因聚合策略。
3.2 四川小麦育成品种和高代品系在不同生态区的成株期抗条锈性
小麦条锈病成株抗性类型(adult plant resistance, APR)一般具有非小种专化性,因而具有潜在的持久抗病性[2324]。APR基因利用也是我国条锈病抗性育种的重要内容,本研究中,育成品种和高代品系在4个位于不同条锈病流行区的鉴定圃成株期抗条锈病鉴定结果不同,说明成株期抗性表型与鉴定圃所在地域环境条件密切相关。育成品种成株期在四川成都(13.9%)和甘肃清水(3.5%)高抗类型所占比例显著少于在湖北荆州(34.8%)和河北廊坊(43.5%),但在甘肃和四川中抗品种数量较为接近;高代品系成株期在四川属于高抗类型所占比例(42.2%)低于在湖北(74.4%)和河北(75.9%)所占比例,但明显高于育成品种中的比例(13.9%),在甘肃仅有8.5%高代品系属于高抗类型(图2)。因此,四川小麦成株期抗条锈病育种取得巨大进展,在四川鉴定圃相对于育成品种13.9%的高抗比例,高代品系的高抗比例增加到42.2%;而中抗材料比例从65.2%减少到50.8%。但在甘肃清水鉴定圃,四川的育成品种和高代品系高抗类型比例仍比较低,说明在甘肃和四川这两个最重要的菌源地,条锈菌的致病类型和侵染环境条件存在较大差异。
3.3 四川小麦材料抗条锈病基因型鉴定与利用
四川省内及其周边条锈菌菌源基地存在丰富的致病变异类型,导致新致病类型不断产生和主栽品种推广应用3~5年后丧失抗病性[1, 8]。因此,培育和筛选抗性持久稳定的主栽品种对有效控制条锈病尤为迫切。在本研究苗期利用3个优势小种和成株期4个地点系统鉴定,可以从中发掘抗性稳定具有持久抗性潜力的基因型。如对小种CYR32、CYR33和CYR34均为高抗类型的全生育期抗病育成品种‘资麦1号‘良麦2号‘科成麦4号‘绵麦37‘川麦53‘川麦58‘川麦64‘宜麦8号和高代品系‘中科麦138,以及在4个鉴定圃均为高抗类型的成株抗性品种‘乐麦3号。‘川农21苗期中抗3个流行小种,成株期在4个鉴定圃中均为高抗类型,含有来自小麦-黑麦1BL/1RS易位系抗病基因Yr41,其抗条锈性已保持了近20年。但其他来自小麦-黑麦1BL/1RS易位系的育成品种如‘川农19‘川农26‘川农27虽然携带Yr41[25],但抗性水平低于‘川农21,推测该品种携带有其他的抗病遗传位点。‘川麦53‘川麦58‘川麦68除含有Yr26外,还含有其他基因[2627]。另外,‘川麦87‘绵麦161‘绵麦319等12份抗条锈病材料可能携带Yr5,‘川麦83‘川育34‘绵麦903等12份材料可能携带Yr15[28]。Yr5是高抗我国当前流行小种如CYR32、CYR33和CYR34全生育期抗性基因[17],但已在澳大利亚[29]、土耳其[30]、印度[31]和中国[32]发现了对Yr5高毒性菌株。2016年后审定的全生育期抗病品种‘蜀麦1613可能携带Yr9+Yr10[33],而‘蜀麦691可能携带Yr17+Yr29+Yr30+Yr67+Yr81[34],通过多基因聚合策略有效地提高了抗性水平。
參考文献
[1] CHEN Wanquan, WELLINGS C, CHEN Xianming, et al. Wheat stripe (yellow) rust caused by Puccinia striiformis f. sp.tritici [J]. Molecular Plant Pathology, 2014, 15(5): 433446.
[2] WELLINGS C R. Global status of stripe rust: a review of historical and current threats [J]. Euphytica, 2011, 179(1): 129141.
[3] LINE R F. Stripe rust of wheat and barley in North America: a retrospective historical review [J].Annual Review of Phytopathology, 2002, 40(40): 75118.
[4] ALI S, GLADIEUX P, LECONTE M, et al. Origin, migration routes and worldwide population genetic structure of the wheat yellow rust pathogen Puccinia striiformis f.sp. tritici [J/OL]. PLoS Pathogens, 2014, 10(1): e1003903. DOI: 10.1371/journal.ppat.1003903.
[5] WAN Anmin, CHEN Xianming, HE Zhonghu. Wheat stripe rust in China [J]. Australian Journal of Agricultural Research, 2007, 58(6): 605619.
[6] 陈万权, 康振生, 马占鸿, 等. 中国小麦条锈病综合治理理论与实践[J]. 中国农业科学, 2013, 46(20): 42544262.
[7] 马占鸿. 中国小麦条锈病研究与防控[J]. 植物保护, 2018, 45(1): 16.
[8] 姚革, 蒋滨, 田承权, 等. 四川省小麦条锈病持续流行原因及防治对策[J]. 西南农业学报, 2004, 17(2): 253256.
[9] HAN Dejun, WANG Qilin, CHEN Xianming, et al. Emerging Yr26virulent races of Puccinia striiformis f. tritici are threatening wheat production in the Sichuan basin, China [J]. Plant Disease, 2015, 99(6): 754760.
[10]ZHENG Shigang, LI Yunfang, LU Lu, et al. Evaluating the contribution of Yr genes to stripe rust resistance breeding through markerassisted detection in wheat [J]. Euphytica, 2017, 213(2): 116.
[11]MCINTOSH R, MU Jingmei, HAN Dejun, et al. Wheat stripe rust resistance gene Yr24/Yr26: A retrospective review [J]. The Crop Journal, 2018, 6: 321329.
[12]吴立人, 牛永春. 我国小麦条锈病持续控制的策略[J]. 中国农业科学, 2000, 33(5): 4654.
[13]LINE R F, CHEN Xianming. Successes in breeding for and managing durable resistance to wheat rusts [J]. Plant Disease, 1995, 79(12): 12541255.
[14]万安民, 袁文焕. 1991~1996年我国小麦条锈菌生理专化研究[J]. 植物病理学报, 1999, 29(1): 35.
[15]CHEN Xianming. Epidemiology and control of stripe rust [Puccinia striiformis f.sp. tritici]on wheat [J]. Canadian Journal of Plant Pathology, 2005, 27(3): 314337.
[16]刘博, 刘太国, 章振羽, 等. 中国小麦条锈菌条中34号的发现及其致病特性[J]. 植物病理学报, 2017, 47(5): 681687.
[17]曾庆东, 沈川, 袁凤平, 等. 小麦抗条锈病已知基因对中国当前流行小种的有效性分析[J]. 植物病理学报, 2015, 45(6): 8392.
[18]庞云星, 崔宏梅, 蔺瑞明, 等. 89份河北、山西小麦品种抗条锈性评价及抗条锈病基因检测[J]. 植物保护, 2021, 47(6): 4957.
[19]CHEN Wanquan, WU Liren, LIU Taiguo, et al. Race dynamics, diversity, and virulence evolution in Puccinia striiformis f.sp. tritici, the causal agent of wheat stripe rust in China from 2003 to 2007 [J]. Plant Disease, 2009, 93(11): 10931101.
[20]沈麗, 罗林明, 陈万权, 等. 四川省小麦条锈病流行区划及菌源传播路径分析[J]. 植物保护, 2008, 35(3): 220226.
[21]向运佳, 章振羽, 沈丽, 等. 2005-2010年四川省小麦条锈病菌毒性变化动态[J]. 西南农业学报, 2013, 26(5): 18581863.
[22]LIU Taiguo, PENG Yunliang, CHEN Wanquan, et al. First detection of virulence in Puccinia striiformis f.sp. tritici in China to resistance genes Yr24 (=Yr26) present in wheat cultivar ‘Chuanmai 42 [J]. Plant Disease, 2010, 94(9): 1163.
[23]ELLIS J G, LAGUDAH E S, SPIELMEYER W, et al. The past, present and future of breeding rust resistant wheat [J/OL]. Frontiers in Plant Science, 2014, 5: 641. DOI: 10.3389/fpls.2014.00641.
[24]GEBREWAHID T W, ZHANG Peipei, ZHOU Yue, et al. QTL mapping of adult plant resistance to stripe rust and leaf rust in a Fuyu 3/Zhengzhou 5389 wheat population [J]. The Crop Journal, 2020, 8(4): 655665.
[25]LUO Peigao, REN Zhenglong, ZHANG Huaiyu, et al. Identification, chromosome location, and diagnostic markers for a new gene (YrCN19) for resistance to wheat stripe rust [J]. Phytopathology, 2005, 95(11): 12661270.
[26]鄭建敏, 罗江陶, 万洪深, 等. 四川省小麦育成品种系谱分析及发展进程[J]. 遗传, 2019, 41(7): 599610.
[27]郑建敏, 罗江陶, 万洪深, 等. 川麦44及其9个衍生品种比较分析[J]. 西南农业学报, 2018, 31(12): 24722477.
猜你喜欢
湖北植保(2022年4期)2022-08-23
南方农业学报(2022年11期)2022-03-18
作物学报(2022年3期)2022-01-22
今日农业(2021年7期)2021-07-28
农民致富之友(2019年31期)2019-11-23
农民致富之友(2019年16期)2019-07-01
西藏农业科技(2018年4期)2018-04-25
植物保护(2017年4期)2017-08-09
中国马铃薯(2017年1期)2017-03-02
植物保护(2017年3期)2017-01-21
