
泛癌中FGFR3表达的预后意义及其免疫调节作用*
2023-06-15 07:24许怡冉何玥李冕鲁艳芹
中国医学工程 2023年5期
许怡冉,何玥,李冕,鲁艳芹,3
(1.山东第一医科大学 临床与基础医学院,山东 济南 250117;2.山东第一医科大学 生物医学科学学院,山东 济南 250117;3.山东第一医科大学第一附属医院 内分泌科,山东 济南 250014 )
FGFR3 基因属于受体酪氨酸激酶(FGFR)家族,位于人类染色体4p16.3,长约16.5 kb,是一种具有自身磷酸化活性的跨膜酪氨酸激酶受体。与配体结合后形成二聚体激活胞内酪氨酸激酶,在细胞内引起一系列级联反应,导致DNA 合成和细胞分裂[1],并进一步激活下游MAPK 及PI3K/AKT 信号通路。其中细胞内MAPK 信号通路与肿瘤细胞增殖、迁移和转移行为相关;PI3K/AKT 信号通路与肿瘤细胞迁移、黏附及肿瘤血管的生成相关[2]。FGFR3 突变多发生在细胞外结构域(R248C,S249C),突变产生的新半胱氨酸残基可与受体二聚体共价结合,导致受体的结构性激活来参与肿瘤的发生。研究表明,约有35%膀胱癌及25%宫颈癌病例中发生了FGFR3 的基因突变,突变主要发生在浅表或乳头状低级别膀胱癌中,与良好的预后及临床分期相关[3-5]。基于大量的文献检索与临床数据,目前仍缺乏FGFR3 与各种肿瘤关系的泛癌证据。该研究利用癌症基因组图谱(The Cancer Genome Atlas,TCGA)和高通量基因表达数据库(Gene Expression Omnibus,GEO)中数据[6-8]分析基因表达、生存预后、免疫微环境和相关蛋白通路的分子特征,研究FGFR3 基因在不同肿瘤发病机制中临床预后及免疫相关性,为探讨潜在的肿瘤免疫治疗靶点提供理论依据。
1 材料与方法
1.1 基因表达分析
利用TIMER2[9]数据库(http://timer.cistrome.org//)中“Gene-DE”模块分析FGFR3 在正常组织与肿瘤组织间表达差异,评估FGFR3 具有治疗靶标的潜力。对于缺乏正常组织对比数据的肿瘤,利用GEPIA2[10]数据库(http://gepia2.cancer-pku.cn/index.html#index)的“Expression Analysis”模块分析,设置“P值=0.01,log2FC=1”,获得这些肿瘤组织与GTEx 数据库中对应的差异数据。通过GEPIA2 网站的“Stage Plot”模块分析多种不同病理分期(Ⅰ期、Ⅱ期、Ⅲ期、Ⅳ期)肿瘤组织中FGFR3 基因表达,用log2(TPM+1)转换后的数据作出“小提琴图谱”。
1.2 生存预后分析
利用GEPIA2“Survival Analysis”模块获得TCGA肿瘤中包含FGFR3 基因的肿瘤总体生存率(overall survival,OS)和无病生存率(disease free survival,DFS)数据图谱,设置表达参数“cutoff-hight(50%),cutoff-low(50%)”区分高表达组和低表达组。假设检验采用Mentel-Cox 检验,通过GEPIA2 的“Survival Analysis”模块绘制单基因生存分析图(Kaplan-Meier)。
1.3 基因变异分析
利 用 cBioPortal[11]数据库(https://www.cbioportal.org/)中“Quick Search”模 块,选择TCGA 泛癌图谱分析,输入FGFR3 进行泛癌基因突变汇总,并在“Mutation”模块查询FGFR3 的基因突变频率,突变类型,拷贝数改变(CAN)、突变位点及蛋白质三维结构。利用“Comparison/Survival”模块获得FGFR3 基因在突变及未突变时,TCGA 肿瘤病例中无疾病的、无进展的和无疾病生存差异的总体数据,同时生成P值为log-rank的Kaplan-Meier 图。利用Rstudio 中TCGAbiolinks软件包下载突变注释文件,利用Spearman 相关性检验评估FGFR3 表达与TMB、MSI 间的相关性,使用配对t检验进行基因共表达分析。
1.4 免疫浸润分析
TIMER2 提供六种常见免疫细胞的渗透评分,包括B 细胞、CD4+T 细胞、CD8+T 细胞、巨噬细胞、中性粒细胞。利用Immune 模块分析TCGA 肿瘤中FGFR3 表达与免疫浸润之间的关系,采用XCell、MCPCOUNTER 和EPIC 算法进行免疫浸润评估。肿瘤纯度是本分析中的主要混杂因素[12],通过纯度调整后的Spearman's 等级相关检验得到P值及偏相关系数(partial correlation,COR)值。数据以热图及散点图的形式展现。
1.5 FGFR3 相关基因的富集分析
通过 GEPIA2 数据库的“Similar Gene Detection”模块,得到与FGFR3 表达相关的前100 个靶向基因,应用log2TPM 方块图,得到P值和相关系数R,利用以上数据制作所选基因与FGFR3 的相关性热图。STRING 数据库是一个检索已知蛋白及预测蛋白间相互作用的数据库[13](https://string-db.org/),通过“Protein by name”模块,输入蛋白名称“FGFR3”,随后设置以下参数:最低交互分数设置为低信度0.150;最大交互对象数量设置为不超过50 个;来源设置为实验。筛选出50 个经实验验证的FGFR3 结合蛋白。使用Venn[14]网站(http://bioinformatics.psb.ugent.be/webtools/Venn/)将相关性前100 基因与蛋白互作的50 个基因进行交叉分析。结合两组数据进KEGG 和GO 通路分析。将基因列表上传到David网 站[15](https://david.ncifcrf.gov/)并选择“OFFICIAL_GENE_SYMBOL”和“Homo Spaiens”分别作为基因标识符和物种,使用cnetlot 函数(Circle=F,ColorEdge=T,Node_Label=T),对生物学过程的分子功能(molecular function,MF)数据通过ggplot2 和ForestPlot 的R 包生成cnet 曲线图[16]。其中,双侧P<0.05 为差异有统计学意义。
2 结果
2.1 基因表达分析
在本研究中,笔者探讨人类FGFR3 基因(mRNA:NM_000142.5,蛋白质 :NP_001341738.1)的致癌作用。系统发育树分析FGFR3 蛋白在不同物种间的进化关系,筛选出相似度最高的14 个物种进行同源分析,发现智人与黑猩猩,白颊长臂猿关系最近。
采用TIMER2 数据库分析TCGA 中FGFR3 mRNA 表达水平。在乳腺浸润癌(BRCA)、胆管癌(CHOL)、肝细胞肝癌(LIHC)、肺鳞癌(LUSC)、皮肤黑色素瘤(SKCM)、甲状腺癌(THCA)组织中FGFR3mRNA 表达水平明显高于正常组织,差异有统计学意义(P<0.001)。在结肠癌(COAD)、肾嫌色细胞癌(KICH)、肾透明细胞癌(KIRC)、肾乳头状细胞癌(KIRP)、肺腺癌(LUAD)组织中FGFR3mRNA 表达水平明显低于正常组织,差异有统计学意义(P<0.001),见图1A。
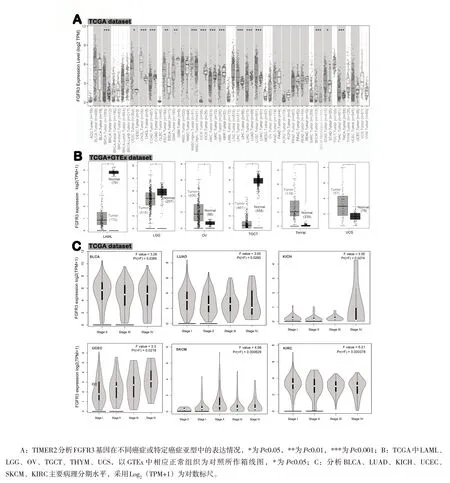
图1 FGFR3 基因在不同肿瘤和病理分期中的表达水平
将TCGA 与GTEx 数据库联合分析,补充GTEx 数据集的正常组织作为对照,进一步分析FGFR3 在卵巢浆液性囊腺癌(OV)、胸腺癌(THYM)、子宫肉瘤(UCS)、急性髓细胞样白血病、(LGG)、睾丸癌(TGCT)的正常组织和肿瘤组织中的表达差异(P<0.01),见图1B。对于肾上腺皮质癌(ACC)、弥漫性大B 细胞淋巴瘤(DLBC)和肉瘤(SARC)差异无统计学意义(P>0.05)。
使用GEPIA2 中“Pathological Stage Plot”模块观察肿瘤中FGFR3 表达与临床分期间相关性,包括膀胱尿路上皮癌(BLCA)、肾透明细胞癌、肾嫌色细胞癌、肺腺癌、皮肤黑色素瘤和子宫内膜癌(UCEC),发现上述肿瘤中FGFR3 表达普遍呈上升趋势,差异有统计学意义(P<0.05),见图1C。
2.2 生存预后分析
根据FGFR3 表达水平,将肿瘤划分为高表达组和低表达组。如图2A 所示,FGFR3 高表达与肾透明细胞癌(P=0.014)总体预后不良有关,低表达的FGFR3 与肾上腺皮质癌(P=0.012)和LGG(P=0.034)总体预后不良有关。无病生存率分析数据显示FGFR3 的高表达与头颈鳞状细胞癌(HNSC)(P=0.0067)患者预后差有关,低表达的FGFR3 与肾上腺皮质癌(P=0.015)、葡萄膜黑色素瘤(UVM)(P=0.0067)患者预后差有关(见图2B)。上述结果表明FGFR3 的表达高低与不同类型肿瘤的预后存在差异。
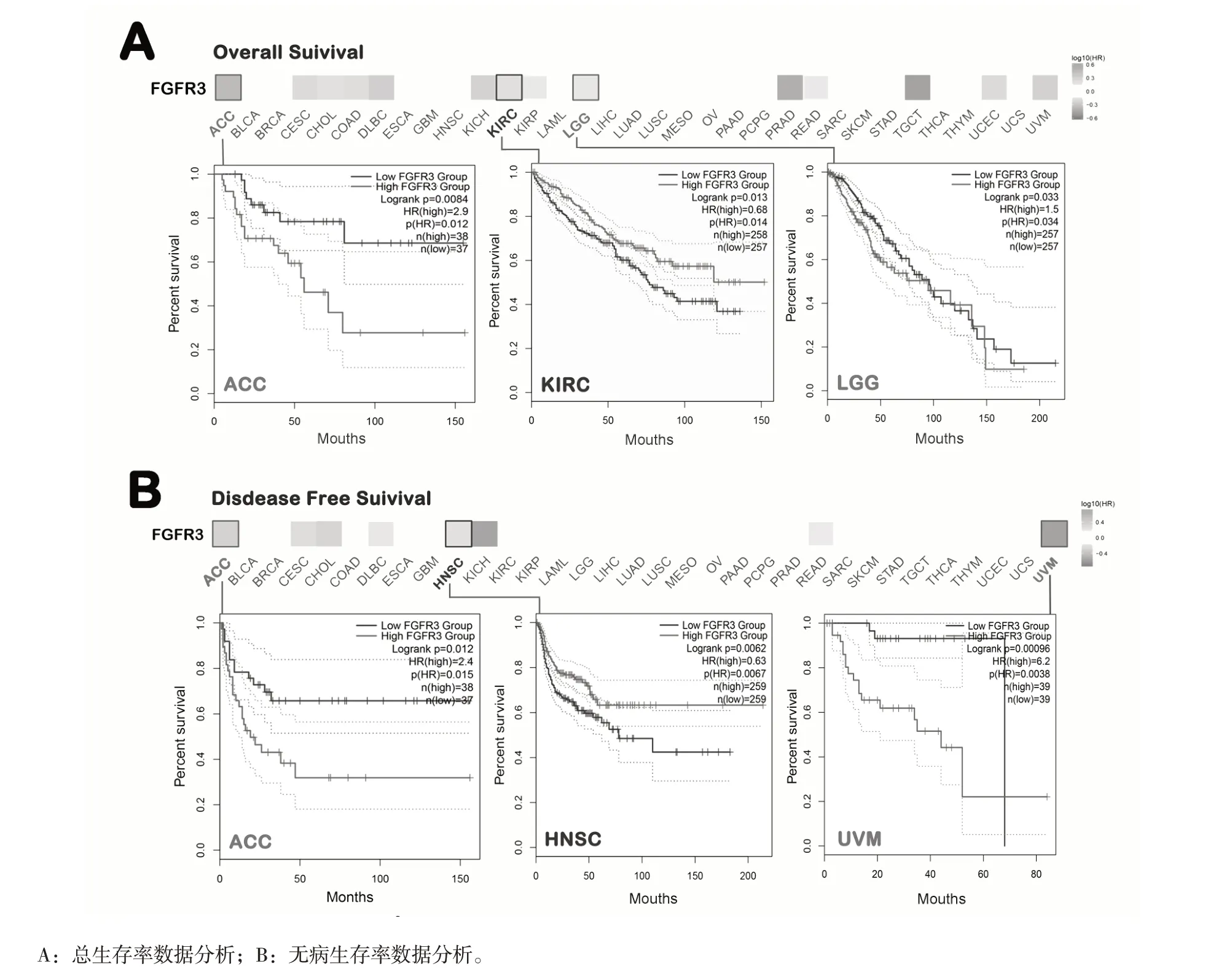
图2 FGFR3 基因与肿瘤生存预后的关系
2.3 基因突变分析
以FGFR3 突变为主要类型的膀胱尿路上皮癌患者,FGFR3 基因的变异频率最高(>18.73%),子宫肉瘤以FGFR3 扩增为主,其变异频率>15.79%,FGFR3 扩增是肾上腺皮质癌和胆管癌患者的唯一变异类型,其变异频率>2%(见图3A)。进一步显示FGFR3 基因变异的类型和位点(见图3B),发现FGFR3 错义突变是遗传变异的主要类型,在1 例KRPCC、4 例LSCC、4 例头颈鳞状细胞癌、32 例LUCC 病例中检测出I-set 结构域中使FGFR3基因249 号位点发生拼接突变(见图3C)。

图3 TCGA 肿瘤中FGFR3 基因突变特征及基因共表达分析
评估FGFR3 表达与不同肿瘤类型的TMB 之间的关系,发现FGFR3 表达与膀胱尿路上皮癌、宫颈鳞癌和腺癌(CESC)、结肠癌、食管癌(ESCA)、头颈鳞状细胞癌、肾乳头状细胞癌、肺腺癌、间皮瘤(MESO)、皮肤黑色素瘤、胸腺癌中的TMB 相关,而与其他肿瘤无显著相关(见图3D)。此外,评估了不同肿瘤中FGFR3 表达与MSI 状态之间的关系,发现FGFR3 表达与肺腺癌、卵巢浆液性囊腺癌、胰腺癌(PAAD)、前列腺癌(PRAD)、直肠腺癌(READ)、子宫内膜癌、葡萄膜黑色素瘤、结肠癌、肾透明细胞癌中的MSI 相关,而与其他肿瘤无明显相关性(见图3E)。
2.4 免疫浸润分析
使用不同算法研究免疫细胞浸润水平与FGFR3 表达间的相关性。基于全部结肠癌、头颈鳞状细胞癌、胰腺癌、前列腺癌和睾丸癌各肿瘤中FGFR 的表达与肿瘤相关成纤维细胞的浸润水平呈负相关,FGFR3 表达与卵巢浆液性囊腺癌相关成纤维细胞呈正相关(图4A)。在此,将上述肿瘤其中一种算法产生的相关性散点图作为代表展现(图4B,Rho=-0.391,P=1.00e-06)。

图4 FGFR3 基因表达与癌相关成纤维细胞免疫浸润分析
2.5 FGFR3 相关基因的富集分析
为了进一步研究FGFR3 基因在肿瘤发生中的分子机制,筛选出FGFR3 靶向结合蛋白及FGFR3表达相关基因进行一系列富集分析。采用STRING数据库,得到50 个基于实验数据支持的FGFR3结合蛋白。图5A 为这些蛋白相互作用网络。使用GEPIA2 工具结合TCGA 肿瘤的表达数据,得到FGFR3 表达相关的前100 个基因。图5C 所示,FGFR3 表达水平与ARHGAP23(R=0.33)、LET(R=0.31)、PVRL1(R=0.34)、SDC1(R=0.32)和TACC3(R=0.4)基因的表达水平呈正相关(P<0.001)。相应热图数据显示FGFR3 在不同肿瘤中的表达差异性(图5D)。将蛋白互作的50 个基因与相关性前100 基因取交叉分析筛选出共同基因PVRL1(图5B)。
结合上述两组数据集进行KEGG 和GO 富集分析。KEGG 数据表明,FGFR3 在肿瘤发病机制中的作用可能与肿瘤信号通路(pathways in cancer)、MAPK 信号通 路(MAPK signaling pathway)和 PI3K-Akt 信号通 路(PI3K-Akt signaling pathway)有关(图5E)。GO 富集分析进一步表明,这些基因大多数与细胞增殖、分化及血管形成途径密切相关(图5F)。
3 讨论
通过文献检索,笔者没有检索到任何从整体角度对FGFR3 进行泛癌分析的文献。本文首次系统地通过生信分析发现FGFR3 在肝癌、睾丸癌等肿瘤组织中高表达,在肺腺癌、结肠癌、子宫肉瘤等组织中低表达。FGFR3 基因在肝癌中高表达,且其高表达和肝癌的恶性程度密切相关[17]。数据显示,肝癌是全球第五大癌症相关死亡原因,在我国其死亡率位居恶性肿瘤第2 位。全球每年新发肝癌病例85.4 万例,中国46.6 万例,约占全球的55%[18],因而研究与开发肝癌特异性靶点药物在我国更加具有重要的意义。FGFR3 通过自分泌或旁分泌机制促进肝癌细胞的生长,但肝癌中信号通路间常存在广泛的交叉,FGF 信号通路联合其他途径将是今后研究的重要方向[19]。睾丸癌是一种多发于青壮年男性的恶性泌尿系统肿瘤,近年来其发病呈上升的趋势[20]。生信研究显示FGFR3 在睾丸癌中表达显著高于正常组织,其高表达与睾丸癌患者预后不良有关。FGFR3 信号在婴儿期和青春期前的精原细胞中活跃,并与磷酸化受体向细胞核的移位有关,FGFR3 信号促进精原细胞存活和增殖,在激活该途径突变的情况下导致睾丸癌的发生[21]。由于睾丸肿瘤比较少见,占泌尿系统肿瘤的5%[20],在临床上关于睾丸癌的研究鲜有报道,睾丸癌在诊断、治疗、预后等方面缺乏敏感性与特异性的理想肿瘤标志物,因此,利用生物信息学手段,寻找特异性较高的睾丸癌标志物具有重要的临床意义,FGFR3 可能成为睾丸癌新的预后标志物及诊疗的潜在靶点。已有文献证实,FGFR3 基因是肺癌细胞转移的重要标志物,FGFR3 基因通过参与RAS/RAF/MEK/MAPK途径及促进上皮间质转化而促进肺癌的转移[22]。中国每年预计新增肺癌病例约70 万,新增肺癌死亡病例约40 万[23]。近期研究表明FGFR3 基因沉
默介导的基质金属蛋白酶9 下调可明显抑制肺癌细胞A549 的侵袭能力[24]。生信分析发现FGFR3高表达与肺癌患者OS、PPS、FP 预后不良有关,表明FGFR3 可能成为肺癌新的诊疗靶点和有效分子预后标志物。FGFR3 在结肠癌中低表达,是重要的肿瘤负调节因子。结肠癌是常见的恶性肿瘤之一,调查显示2021 年结肠癌为全球癌症死亡的主要原因[25]。研究表明FGFR3 表达抑制了结肠癌细胞的增殖、迁移和侵袭能力,通过阻断FGFR3-IIIc 信号传导促进了结肠癌细胞凋亡[26]。表明FGFR3 有望成为治疗结肠癌的潜在靶点,可能是结肠癌预后的重要分子标志物。
恶性肿瘤的最大标志是不受控制的细胞增殖,酪氨酸激酶在增殖信号的转导上有重要作用,当它由于体内突变使得调节异常时,可以促进肿瘤的发生。以往人们对FGFR3 突变的研究主要围绕骨发育相关的人类遗传病,如FGFR3 特殊位点突变导致软骨发育不良[27]。近年来国外研究表明,FGFR 作为酪氨酸家族的一员,证实与肿瘤的发展密切相关[28],尤其在膀胱癌中。生信分析观察到,FGFR3 最主要的变异类型是错义突变,在膀胱尿路上皮癌中突变频率最高(>18.73%),FGFR3 的表达与膀胱尿路上皮癌患者的肿瘤突变负荷具有相关性,并且与大多数免疫点基因呈负相关。膀胱癌是发病率较高的癌症类型[29],已有实验证实,FGFR3 突变与膀胱癌预后差有关[30],FGFR3 突变能预测肌层浸润性膀胱癌术前新辅助化疗的疗效[31],说明FGFR3 是膀胱癌重要的治疗和预后分析辅助标志物。分析发现FGFR3 在子宫肉瘤中的变异频率同样较高(>15.79%),FGFR3 在子宫肉瘤中低表达,并且其扩增与子宫肉瘤预后差有关。子宫肉瘤是一类恶性间叶组织源性肿瘤,预后较差[32],研究证实,RB1、TP53 和PTEN 是子宫肉瘤的重要分子标记物,子宫平滑肌肉瘤患者中TP53 突变的频率为24%~30%,PTEN 突变的频率为42%~58%[33]。因此FGFR3 也可能成为子宫肉瘤的新的诊断和预后分子标志物,这需要在前瞻性的临床试验中加以评估。
阻断免疫检查点途径是肿瘤免疫的有效措施,分析发现,在膀胱尿路上皮癌、结肠癌和睾丸癌中FGFR3 高表达与绝大部分检查点基因表达水平呈显著负相关,表明FGFR3 可能参与多种肿瘤的免疫调节。这些检查点基因的上调可能与FGFR3诱导MAPK/ERK 通路,引起的免疫细胞分化和极化有关。在结肠癌、睾丸癌中FGFR3 表达与肿瘤相关成纤维细胞的免疫渗透水平呈负相关。因此,FGFR3 不仅是肿瘤潜在的预后预测因子,还可能是重要的肿瘤免疫调节因子。
综上所述,FGFR3 的靶向免疫治疗研究尚处于初级阶段,很多问题仍然存在,如某些突变使得酪氨酸激酶耐药性的出现及MAPK/ERK 信号网络的代偿性作用,均可使FGFR3 的分子靶向治疗出现阻碍。因此,以FGFR3 为靶向的肿瘤基因治疗在临床上的应用价值尚待探索。通过FGFR3 的泛癌分析,有助于从临床角度理解FGFR3 在肿瘤发生发展中的作用,为临床诊断治疗提供新的思路。
猜你喜欢
中国临床医学影像杂志(2019年4期)2019-06-18
中成药(2018年7期)2018-08-04
西南国防医药(2016年7期)2016-12-01
中国组织化学与细胞化学杂志(2016年3期)2016-02-27
西南军医(2016年3期)2016-01-23
中国卫生标准管理(2015年1期)2016-01-14
磁共振成像(2015年1期)2015-12-23
中国当代医药(2015年17期)2015-03-01
郑州大学学报(医学版)(2015年1期)2015-02-27
西南军医(2014年5期)2014-04-25
