
野鸦椿与圆齿野鸦椿对NaCl和NaHCO3耐受性的比较
2023-07-10 00:28吴慧平黄运腾姜雪茹涂淑萍
江苏农业科学 2023年11期
吴慧平 黄运腾 姜雪茹 涂淑萍
摘要:比较野鸦椿和圆齿野鸦椿的抗盐性,为野鸦椿及圆齿野鸦椿的引种、栽培、养护和管理提供科学依据。以3年生盆栽野鸦椿和圆齿野鸦椿为材料,采用不同质量浓度的NaCl(浇施2.920、5.256、7.008 g/盆)和NaHCO3(浇施4.200、7.560、10.080 g/盆)进行单盐胁迫处理,并以浇施去离子水为对照(CK),比较不同处理下2个树种叶片PSⅡ最大光化学效率(Fv/Fm)、PSⅡ实际光化学效率[Y(Ⅱ)]、光化学淬灭系数(qP)、非光化学淬灭系数(NPQ)、丙二醛(MDA)含量、超氧化物歧化酶(SOD)活性、可溶性蛋白(SP)含量和可溶性糖(SS)含量的变化。利用皮尔逊相关性分析法对野鸦椿和圆齿野鸦椿2个树种不同盐浓度与上述各生理生化指标进行相关性分析,在此基础上,运用隶属函数法进行2个树种对2种盐的耐受能力的综合性评价。结果表明,当NaCl溶液浇施浓度≥5.256 g/盆时,野鸦椿叶片PSⅡ最大光化学效率、实际光化学效率以及光化学淬灭系数均显著低于CK,而非光化学淬灭系数则高于CK;浇施不同质量浓度的NaHCO3处理下野鸦椿上述4个指标与CK处理无显著差异。圆齿野鸦椿在以上2种盐不同质量浓度胁迫处理下上述4个指标与CK均无显著差异。随着NaCl和NaHCO3浓度的升高,叶片丙二醛含量表现为:野鸦椿呈先升后降趋势,而圆齿野鸦椿则呈持续上升趋势;叶片可溶性蛋白和可溶性糖含量表现为:2个树种均呈先下降后上升趋势,且叶片可溶性蛋白含量表现为:野鸦椿均高于圆齿野鸦椿,叶片可溶性糖含量则表现为:野鸦椿均低于圆齿野鸦椿。隶属函数分析结果表明,野鸦椿对NaCl和NaHCO3的耐受性均比圆齿野鸦椿强。
关键词:圆齿野鸦椿;野鸦椿;生理生化特性;盐胁迫;耐盐性
中图分类号:S718.43文献标志码:A文章编号:1002-1302(2023)11-0160-07
收稿日期:2022-09-03
基金项目:江西省自然科学基金面上青年项目(编号:20212BAB215014);江西省林业局科技创新专项资金项目(编号:201812);江西省教育厅青年项目(编号:GJJ200446)。
作者简介:吴慧平(1997—),女,河南新蔡人,硕士研究生,主要从事园林植物繁育与栽培研究。E-mail:1286151012@qq.com。
通信作者:涂淑萍,教授,主要从事园林植物繁育与栽培研究。E-mail:jxtsping@163.com。土壤盐碱化是一个世界性的环境和生态问题,同时也是土地退化的形式之一[1]。土壤盐碱化严重抑制了植物的正常生长和发育,甚至引起植物死亡[2-3]。我国盐碱化土地面广量大,在东北西部、华北、西北和滨海地区均有分布[4]。因此,培育耐盐碱的优良观赏树种的需求也日益迫切[5]。野鸦椿[Euscaphis japonica (Thumb.) Dippel]和圆齿野鸦椿(E. konishii Hayata)隶属于省沽油科(Taphyleaceae)野鸦椿属(Euscaphis)的2种药赏兼用植物[6]。前者为落叶树种,后者为常绿树种。野鸦椿属植物观赏价值高,开发利用前景十分广阔,探究其对盐碱胁迫抗性对于其向海滨盐碱城市引种推广以及日常栽培养护具有重要意义。目前,有关野鸦椿和圆齿野鸦椿的抗逆性研究多见于水分协迫和温度协迫等方面。有研究表明,在干旱胁迫条件下不同家系的圆齿野鸦椿耐旱性不同,并筛选了2种优良抗旱家系[7]。圆齿野鸦椿具有一定的抗涝性[8],属中度抗寒植物[9]。刘福辉探究了高温胁迫下圆齿野鸦椿幼苗生理代谢的变化,发现高温会影响其碳同化及光合作用等[10]。此外,在圆齿野鸦椿苗木生长期喷施多效唑、喷施宝、圆丰素等植物生长调节剂有利于提高其抗性[11-13],但尚未见野鸦椿和圆齿野鸦椿的耐盐碱性研究以及两者之间耐盐性强弱的研究报道。本试验以3年生野鴉椿和圆齿野鸦椿苗为试验材料,比较这2个树种分别在中性盐(NaCl)和碱性盐(NaHCO3)胁迫下生理生化指标的变化,旨在探究二者抗盐碱性的强弱,为野鸦椿和圆齿野鸦椿的栽培养护管理以及在盐碱地区的引种推广提供理论依据。
1材料与方法
1.1试验地概况
本研究地点位于江西农业大学花卉基地(28°46′N、115°55′E,海拔 50 m)[14],年平均气温为17.5 ℃,年日照时长为 1 903.9 h,年降水量为 1 596.4 mm。
1.2试验材料
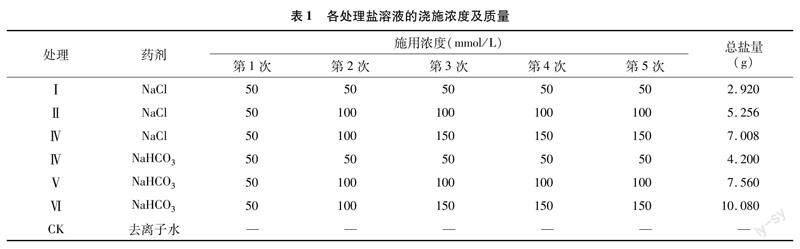
以生长状况基本一致的野鸦椿和圆齿野鸦椿3年生盆栽苗各63株为试验材料。2021年3月将试验用野鸦椿及圆齿野鸦椿苗木移栽至口径16 cm、高17.5 cm的1加仑盆中,栽培基质为园土 ∶泥炭 ∶蛭石 ∶有机肥=2 ∶1 ∶1 ∶0.5。日常管理措施基本保持一致。于2021年8月进行浇盐处理。盆底垫托盘收集渗出液,以避免处理溶液盐分流失。
试验所用药剂为NaCl和 NaHCO3均为西陇科学股份有限公司生产的分析纯。
1.3试验方法
1.3.1试验设计本试验根据NaCl和NaHCO3浇施质量的不同各设置3个处理,以浇施去离子水为对照(CK),共7个处理。施用方法采用土壤浇灌法,于17:00左右浇施1次/d,浇灌量为200 mL/次,浇施5次盐溶液后,各处理植株均浇施200 mL/d去离子水,详见表1。每个处理3盆,每盆1株,重复3次。以完成第5次浇施盐溶液处理的时间记为胁迫处理后0 d,于胁迫处理后10 d进行叶片叶绿素荧光参数测定,于处理后17 d分别采集各处理植株顶芽下第3、第4枚复叶,用于生理生化指标的测定。
1.3.2测定指标及方法叶绿素荧光参数采用德国PAM-2500叶绿素荧光仪进行测定[15];丙二醛(MDA)含量采用硫代巴比妥酸显色法进行测定[16];超氧化物歧化酶活性(SOD)采用试剂盒 WST-8 法测定;可溶性蛋白(SP)含量采用BCA法蛋白定量试剂盒测定;可溶性糖(SS)含量采用植物可溶性糖含量检测试剂盒测定。上述试剂盒均由苏州科铭公司生产。
1.4统计分析
采用Excel 2020、SPSS 22.0软件进行数据的方差分析、多重比较(Duncans)和皮尔逊(Pearson)相关性分析。利用Origin 2018进行图表绘制,并利用隶属函数法综合评价2个树种苗木对NaCl和NaHCO3耐受性。隸属函数值的计算公式[15]为:
Xu=(X-Xmin)/(Xmax-Xmin)。(1)
当所测定指标与树种抗盐性呈负相关时,则利用反隶属函数计算。
Xu反=1-(X-Xmin)/(Xmax-Xmin)。(2)
式中:X为各处理某一指标测定值;Xmin为所有处理中该指标的最小值;Xmax为所有处理中该指标的最大值。
2结果与分析
2.1不同处理对野鸦椿和圆齿野鸦椿叶绿素荧光参数的影响
由表2可知,在不同质量浓度NaCl胁迫处理(处理Ⅰ~处理Ⅲ)下,野鸦椿叶片光系统Ⅱ的最大光化学效率(Fv/Fm)、实际光化学效率[Y(Ⅱ)]以及光化学淬灭系数(qP)3个指标均以CK最大,与处理Ⅰ(NaCl施用量2.920 g/盆)差异均不显著,与处理Ⅱ(NaCl施用量5.256 g/盆)和处理Ⅲ(NaCl施用量7.008 g/盆)差异均达显著水平;而在不同质量浓度NaHCO3胁迫处理(处理Ⅳ~处理Ⅵ)下,上述3个指标各胁迫处理与CK之间差异均不显著。而各胁迫处理非光化学淬灭系数(NPQ)与CK之间差异均不显著。
由表3可知,不同处理之间圆齿野鸦椿叶片最大光化学效率、实际光化学效率以及光化学淬灭系数3个指标差异均不显著;非光化学淬灭系数以处理Ⅵ(NaHCO3施用量10.080 g/盆)最小,为0.88,除与CK和处理Ⅴ(NaHCO3施用量7.560 g/盆)差异不显著外,与其他处理差异均达显著水平。
2.2不同处理对野鸦椿和圆齿野鸦椿叶片丙二醛含量的影响
由图1可知,野鸦椿叶片丙二醛含量在不同质量浓度NaCl处理(处理Ⅰ~处理Ⅲ)与CK之间差异均不显著;而圆齿野鸦椿叶片丙二醛含量,以处理Ⅲ(NaCl施用量7.008 g/盆)最高,显著高于CK和其他2个质量浓度处理。在不同质量浓度NaHCO3胁迫处理下,2个树种叶片丙二醛含量均以处理Ⅵ(NaHCO3施用量10.080 g/盆)最高,且各处理间无显著差异。
2.3不同处理对野鸦椿和圆齿野鸦椿叶片超氧化物歧化酶活性的影响
由图2可知,在不同质量浓度NaCl和NaHCO3胁迫处理下,野鸦椿与圆齿野鸦椿叶片超氧化物歧化酶活性均随着盐浓度升高而呈先升高后降低的变化趋势。在不同质量浓度NaCl胁迫处理(处理Ⅰ~处理Ⅲ)下,野鸦椿叶片超氧化物歧化酶活性以处理Ⅲ(NaCl施用量7.008 g/盆)最低,且与其他2个胁迫处理相比差异达显著水平;而圆齿野鸦椿叶片超氧化物歧化酶活性在各处理之间差异不显著。在不同质量浓度NaHCO3胁迫处理(处理Ⅳ~处理Ⅵ)下,野鸦椿叶片超氧化物歧化酶活性在各处理之间差异不显著;圆齿野鸦椿叶片超氧化物歧化酶活性以处理Ⅴ(NaHCO3施用量7.560 g/盆)最高,除与处理Ⅵ(NaHCO3施用量10.080 g/盆)差异不显著外,与处理Ⅳ(NaHCO3施用量4.200 g/盆)和CK差异均达显著水平。
2.4不同处理对野鸦椿和圆齿野鸦椿叶片可溶性蛋白含量的影响
由图3可知,在不同质量浓度NaCl胁迫处理(处理Ⅰ~处理Ⅲ)下,野鸦椿叶片可溶性蛋白含量以处理Ⅰ(NaCl施用量2.920 g/盆)最低,显著低于CK及处理Ⅱ(NaCl施用量5.256 g/盆)和处理Ⅲ(NaCl施用量7.008 g/盆),圆齿野鸦椿叶片可溶性蛋白含量在各处理(处理Ⅰ~处理Ⅲ)间差异不显著,但它们与CK均存在显著差异。在不同质量浓度NaHCO3胁迫处理(处理Ⅳ~处理Ⅵ)下,野鸦椿叶片可溶性蛋白含量以处理Ⅴ(NaHCO3施用量7.560 g/盆)最低,显著低于其他各处理,处理Ⅵ(NaHCO3施用量10.080 g/盆)最高,显著高于其他处理;圆齿野鸦椿叶片可溶性蛋白含量各处理间差异均不显著。
2.5不同处理对野鸦椿和圆齿野鸦椿叶片可溶性糖含量的影响
由图4可知,在不同质量浓度胁迫处理下,各胁迫处理野鸦椿和圆齿野鸦椿叶片可溶性糖含量与CK差异均不显著;各胁迫处理之间,除圆齿野鸦椿叶片可溶性糖含量处理Ⅳ(NaHCO3施用量 4.200 g/盆)显著低于处理Ⅵ(NaHCO3施用量 10.080 g/盆)之外,其他各处理间差异均不显著。
2.6盐浓度与野鸦椿和圆齿野鸦椿叶片各生理生化指标的相关分析
由表4可知,NaCl质量浓度与野鸦椿叶片最大光化学效率(Fv/Fm)、实际光合效率[Y(Ⅱ)]、光化学淬灭系数(qP)之间呈极显著负相关,相关系数分别为-0.78、-0.81和-0.74。NaHCO3质量浓度与野鸦椿叶片各生理生化指标之间相关性均不显著。
野鸦椿叶片各生理生化指标表现如下,叶片超氧化物歧化酶(SOD)活性与丙二醛(MDA)含量呈显著负相关(r=-0.49);叶绿素荧光参数之间表现如下,最大光化学效率与实际光化学效率、光化学淬灭系数呈极显著正相关,相关系数分别为0.82和0.75,与非光化学淬灭系数(NPQ)呈显著负相关(r=-0.50)。实际光化学效率与光化学淬灭系数呈极显著正相关(r=0.98),与非光化学淬灭系数呈极显著负相关(r=-0.81);光化学淬灭系数与非光化学淬灭系数呈极显著负相关(r=-0.74)。
由表5可知,NaCl质量浓度与圆齿野鸦椿叶片可溶性蛋白(SP)含量呈极显著正相关(r=0.68),与叶片丙二醛含量呈极显著负相关(r=-0.81)。NaHCO3质量浓度与圆齿野鸦椿叶片可溶性糖(SS)含量呈显著正相关(r=0.60)。
圆齿野鸦椿叶片可溶性蛋白含量与叶片PSⅡ实际光化学效率呈极显著负相关(r=-0.62),与光化学淬灭系数呈显著负相关(r=-0.55),与非光化学淬灭系数呈显著正相关;叶片叶绿素实际光化学效率与光化学淬灭系数呈极显著正相关(r=0.98),与非光化学淬灭系数呈显著负相关(r=-0.81);非光化学淬灭系数与光化学淬灭系数呈极显著负相关(r=-0.81),与最大光化学效率呈显著正相关(r=0.47)。
2.7野鸦椿与圆齿野鸦椿对NaCl和NaHCO3耐受性的综合评价
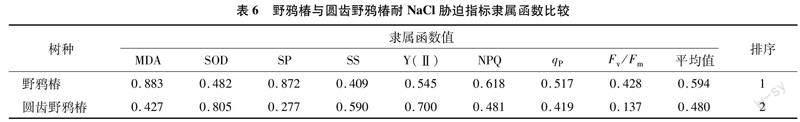
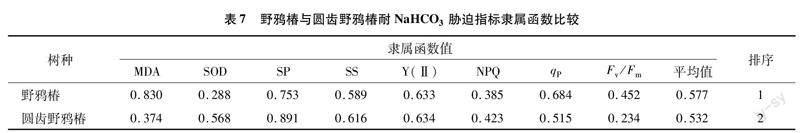
利用模糊数学中隶属函数法,对各处理野鸦椿和圆齿野鸦椿叶片超氧化物歧化酶活性、丙二醛含量、可溶可溶性蛋白含量和可溶性糖含量,以及实际光化学效率、光化学淬灭系数、非光化学淬灭系数、最大光化学效率等8个指标进行综合分析,综合评价野鸦椿与圆齿野鸦椿对NaCl和NaHCO3的耐受性。
由表4可知,野鸦椿叶片丙二醛含量、超氧化物歧化酶活性、实际光化学效率、光化学淬灭、最大光化学效率这5个指标与NaCl质量浓度呈负相关,故野鸦椿在NaCl胁迫下的这5个指标则求其反隶属函数值;野鸦椿叶片丙二醛含量、超氧化物歧化酶活性、可溶性糖含量、非光化学淬灭系数、最大光化学效率这5个指标与NaHCO3质量浓度呈负相关,故野鸦椿在NaHCO3胁迫下的这5个指标则求其反隶属函数值。
由表5可知,圆齿野鸦椿叶片丙二醛含量、超氧化物歧化酶活性、最大光化学效率、光化学淬灭系数这4个指标与NaCl质量浓度呈负相关,故圆齿野鸦椿在NaCl胁迫下的这4个指标则求其反隶属函数值;圆齿野鸦椿叶片丙二醛含量、可溶性蛋白含量、非光化学淬灭系数、最大光化学效率等4个指标与NaHCO3质量浓度呈负相关,故圆齿野鸦椿在NaHCO3胁迫下的上述4个指标求其反隶属函数值。
将每个处理各项指标的隶属函数值(Xu)累加,求平均值,平均隶属函数值越大,则抗性越强。由表6和表7可知,野鸦椿对中性盐(NaCl)和碱性盐(NaHCO3)的耐受性均强于圆齿野鸦椿。
3討论
3.1不同盐胁迫对野鸦椿和圆齿野鸦椿叶片叶绿素荧光参数的影响
叶片叶绿素荧光参数的变化能直接或者间接反映出植物叶片光合作用的原初反应和电子同化过程,常被用来研究植物体的光合作用与其作用环境之间的关系[17]。孙云飞等研究发现,茅苍尔(Atractylodes lancea)叶片中叶绿素含量、叶片PSⅡ最大光化学效率等随着盐浓度的增加呈逐渐下降的趋势[18];包颖等研究发现,随着盐浓度的升高,金娃娃萱草(Hemerocallis fulva ‘Golden Doll)叶片最大光化学效率、实际光化学效率等均呈下降趋势[19]。本研究中在NaCl和NaHCO3胁迫下,野鸦椿和圆齿野鸦椿叶片最大光化学效率、实际光化学效率等均随胁迫浓度的增加而降低,这与大多数研究结果相符。
3.2不同盐胁迫对野鸦椿和圆齿野鸦椿叶片丙二醛含量的影响
当植物处于逆境或者遭受胁迫时,细胞膜首先会受到损伤,细胞膜的破坏程度可以反映出植物受盐害程度,而丙二醛作为膜脂过氧化的最终产物,是衡量细胞膜损伤的常用指标之一[20]。研究表明,植物体内的丙二醛含量会随着胁迫时间的延长、胁迫程度的加深表现出持续变高的趋势[21-23]。本研究中,野鸦椿叶片丙二醛含量随着NaCl和NaHCO3胁迫浓度的增加而升高,这与大多数研究结果相符。圆齿野鸦椿叶片丙二醛含量随着NaCl浓度的升高呈现出先升高后降低再升高的趋势,这与缪珊等对狼尾草(Pennisetum alopecuroides)的研究结果[24]相近,在盐胁迫浓度较低时,植物细胞膜对土壤含盐量的变化较为敏感,当细胞膜受到损伤时,丙二醛含量上升;随着土壤含盐量的进一步增加,细胞内抗氧化保护酶系统在盐胁迫诱导下活性增强,并对质膜进行修复,叶片丙二醛含量降低,但当土壤含盐量进一步上升,超过抗氧化保护酶系统耐受阈值时,细胞膜防御系统再次受到损坏,使其丙二醛含量再度升高。
3.3不同盐胁迫对野鸦椿和圆齿野鸦椿叶片超氧化物歧化酶活性的影响
植物遭受盐胁迫时,细胞内氧化还原平衡被破坏,促使超氧化物歧化酶产生,超氧化物歧化酶活性在一定程度上可以反映植物的抗逆性[25]。多数研究表明,随着盐胁迫程度的加深,植物体内超氧化物歧化酶的活性呈现出先上升后下降趋势[26-27]。在本研究中,野鸦椿和圆齿野鸦椿叶片超氧化物歧化酶活性随着NaCl和NaHCO3质量浓度的增高,均呈先升高后降低的变化趋势,这与前人研究结果相一致。
3.4不同盐胁迫对野鸦椿和圆齿野鸦椿叶片可溶性蛋白含量的影响
植物体内的可溶性蛋白含量可以衡量植物在遭受胁迫时的代谢强度。同时,通过可溶性蛋白的累积来降低植物体内渗透势,缓解盐害[28]。罗燕杰等研究发现,在NaCl胁迫下元宝枫(Acer truncatum)叶片中的可溶性蛋白含量随着盐浓度的升高呈现出先升高后下降的变化趋势[29];田晓艳等对牧草的研究发现,叶片可溶性蛋白含量与盐胁迫处理浓度呈显著正相关[30]。在本研究中,随着NaCl质量浓度的增高,圆齿野鸦椿叶片的可溶性蛋白含量呈先升高后降低趋势,这与大多数研究相符。而野鸦椿叶片可溶性蛋白含量随着NaCl和NaHCO3质量浓度升高呈先下降后上升趋势,其原因可能是野鸦椿对于盐胁迫更敏感,胁迫初期细胞膜系统受到损伤,使得可溶性蛋白含量下降,但经过一段时间的适应,植物体为抵御胁迫和缓解伤害,使可溶性蛋白含量上升。
3.5不同盐胁迫对野鸦椿和圆齿野鸦椿叶片可溶性糖含量的影响
可溶性糖是光合作用最终产物之一,是植物合成有机物碳架的能量来源,可以维持植物蛋白质稳定和参与糖类合成运输的功能,其含量也可以反映盐胁迫强度[31-33]。杨宏伟等在对藜麦(Chenopodium quinoa)的盐胁迫处理中发现,随着盐浓度的升高,藜麦幼苗可溶性糖含量呈升高趋势[32]。张林平等研究发现,甘草(Glycyrrhiza uralensis)在NaCl胁迫下,根、叶中可溶性糖含量随着盐浓度的增加呈上升趋势[34]。在本研究中,在NaCl和NaHCO3胁迫下,野鸦椿和圆齿野鸦椿叶片可溶性糖含量均表现为随着盐浓度升高呈先下降再升高的趋势,这与李雅男的研究结果[35]相近。由于盐胁迫的存在,使得酶活性受到抑制,糖的合成受到抑制,而后野鸦椿对盐胁迫的适应性增强,可溶性糖的合成并参与渗透调节,以缓解盐害。
4结论
本研究结果显示,在NaCl和NaHCO3等2种不同盐质量浓度的胁迫下,野鸦椿和圆齿野鸦椿受盐害损伤程度均随着基质含盐量的升高而加深,且野鸦椿对中性盐(NaCl)和碱性盐(NaHCO3)的耐受性均强于圆齒野鸦椿。
参考文献:
[1]舒开林,赵亚楠,王建鑫,等. 盐胁迫下对植物生长影响的研究进展[C]//华北五省市(区)环境科学学会第二十二届学术年会论文集.2021:18-24.
[2]罗燕杰. 蜡梅对盐碱胁迫的生理响应研究[D]. 北京:北京林业大学,2019:1-5.
[3]赵可夫,邹琦,李德全,等. 盐分和水分胁迫对盐生和非盐生植物细胞膜脂过氧化作用的效应[J]. 植物学报,1993,35(7):519-525.
[4]王波,宋凤斌,张金才. 植物耐盐性研究进展[J]. 农业系统科学与综合研究,2007,23(2):212-216.
[5]van Zelm E,Zhang Y X,Testerink C.Salt tolerance mechanisms of plants[J]. Annual Review of Plant Biology,2020,71:403-433.
[6]邹双全. 圆齿野鸦椿研究[M]. 北京:科学出版社,2018:1-3.
[7]陆思羽,李悦,陶凌剑,等. 干旱胁迫下不同圆齿野鸦椿家系苗木生理生化指标的变化[J]. 南方农业学报,2020,51(6):1400-1408.
[8]支丽燕,胡松竹,余林,等. 涝渍胁迫对圆齿野鸦椿苗期生长及其叶片生理的影响[J]. 江西农业大学学报,2008,30(2):279-282.
[9]康文娟. 圆齿野鸦椿家系苗期生长特性及抗低温胁迫能力研究[D]. 南昌:江西农业大学,2015:34-35.
[10]刘福辉. 高温胁迫对圆齿野鸦椿幼苗生理代谢的影响[J]. 林业勘察设计,2017,37(4):26-31.
[11]胡钱萍,于志民,涂淑萍.喷施宝对圆齿野鸦椿生长及抗逆性的影响[C]//第四届全国花卉资源、育种、栽培及应用技术交流会论文汇编.呼和浩特,2016:104-109.
[12]康文娟,马晓蒙,涂淑萍,等. 喷施多效唑对圆齿野鸦椿苗木抗旱性的影响[J]. 江西农业大学学报,2014,36(6):1310-1315.
[13]涂淑萍,马晓蒙,游双红,等. 园丰素对圆齿野鸦椿幼苗生长及其抗旱性的影响[J]. 经济林研究,2013,31(2):121-124.
[14]陶凌剑,涂淑萍,金莉颖,等. 光照强度对圆齿野鸦椿叶片光合特性和叶绿素荧光参数的影响[J]. 经济林研究,2022,40(2):225-231.
[15]涂淑萍,黄航,杜曲,等. 不同品种茶树叶片光合特性与叶绿素荧光参数的比较[J]. 江西农业大学学报,2021,43(5):1098-1106.
[16]李合生. 植物生理生化实验原理和技术[M]. 北京:高等教育出版社,2000:260-261.
[17]Genty B,Briantais J M,Baker N R. The relationship between the quantum yield of photosynthetic electron transport and quenching of chlorophyll fluorescence[J]. Biochimica et Biophysica Acta,1989,990(1):87-92.
[18]孙云飞,张文明,巢建国,等. 盐胁迫对茅苍术叶绿素含量及叶绿素荧光参数的影响[J]. 江苏农业科学,2020,48(4):146-149.
[19]包颖,王嘉欣,陈超,等. NaCl和NaHCO3胁迫对萱草金娃娃光合作用及叶绿素荧光特性的影响[J]. 江苏农业科学,2020,48(3):133-140.
[20]钟旻依,张新全,杨昕颖,等. 植物对重金属铬胁迫响应机制的研究进展[J]. 草业科学,2019,36(8):1962-1975.
[21]王保平,董晓燕,董宽虎. 盐碱胁迫对紫花苜蓿幼苗生理特性的影响[J]. 草地学报,2013,21(6):1124-1129.
[22]李會云,郭修武. 盐胁迫对葡萄砧木叶片保护酶活性和丙二醛含量的影响[J]. 果树学报,2008,25(2):240-243.
[23]王旭明,赵夏夏,陈景阳,等. 盐胁迫下海水稻抗逆生理响应分析[J]. 中国生态农业学报,2019,27(5):747-756.
[24]缪珊,夏振平,李志强. NaCl胁迫对三种狼尾草生长及生理特性的影响[J]. 黑龙江农业科学,2019(6):132-136.
[25]高涵,洪丕征,郭新送,等. 盐胁迫对几种盐生植物幼苗生长及生理生化指标的影响[J]. 山东农业大学学报(自然科学版),2021,52(5):731-738.
[26]张梅茹,马金彪,姚银安,等. 盐角草(Salicornia europaea)对NaCl处理的生理响应[J]. 中国沙漠,2014,34(2):441-447.
[27]贾文庆,尤扬,刘会超,等. 盐胁迫对白三叶茎SOD和MDA的影响[J]. 贵州农业科学,2009,37(3):36-38.
[28]姚佳,刘信宝,崔鑫,等. 不同NaCl胁迫对苗期扁蓿豆渗透调节物质及光合生理的影响[J]. 草业学报,2015,24(5):91-99.
[29]罗燕杰,陈晨,赵靓,等. 元宝枫对盐胁迫的生理响应与耐盐性评价[M]. 北京:中国林业出版社,2018:293-300.
[30]田晓艳,刘延吉,郭迎春.盐胁迫对NHC牧草Na+、K+、Pro、可溶性糖及可溶性蛋白的影响[J]. 草业科学,2008,25(10):34-38.
[31]黄相玲,林妃妃,张明月,等. 盐胁迫对小叶榄仁幼苗生长和渗透调节物质含量的影响[J]. 南方农业学报,2018,49(7):1364-1369.
[32]杨宏伟,刘文瑜,沈宝云,等. NaCl胁迫对藜麦种子萌发和幼苗生理特性的影响[J]. 草业学报,2017,26(8):146-153.
[33]侯江涛,杨雅兰,赵莹菲,等. 海藻精对NaCl胁迫下黄瓜幼苗抗性生理的影响[J]. 江苏农业科学,2022,50(13):165-170.
[34]张林平,刘艳,王洋,等. NaCl胁迫对甘草生长和渗透调节物质积累的影响[J]. 内蒙古农业大学学报(自然科学版),2020,41(3):10-15.
[35]李雅男. 碱性盐胁迫下百合的生理响应机制[D]. 北京:中国农业科学院,2016:16-17.
猜你喜欢
湖南农业大学学报(自然科学版)(2023年2期)2023-05-08
江苏农业学报(2022年6期)2022-02-06
河南农业科学(2020年2期)2020-03-11
山东农业科学(2016年12期)2017-01-21
上海农业学报(2016年2期)2016-10-27
浙江农业学报(2016年7期)2016-06-15
中国糖料(2013年4期)2013-03-02
