
海拔和籼粳细胞质背景对水稻F2代群体杂种优势和自交衰退的影响
2023-07-22 13:50王石华
南方农业学报 2023年2期
关键词:杂种优势

摘要:【目的】分析海拔引起的環境温度变化对籼粳稻细胞质背景下的F1代配子基因型的选择效应及其对后代分离群体优势和衰退的影响,为水稻育种及全球气温变化对水稻遗传变异的效应研究提供理论参考。【方法】以具有籼稻细胞质背景的改良粳稻品种南34与耐冷性极强的粳稻老品种小麻谷为亲本,将正反交F1代种植在3个海拔条件下产生6个F2代群体,对其11个生物学性状的杂种优势及自交衰退进行分析。【结果】株高、剑叶长、剑叶宽、有效穗数、主穗长、穗颈长、实粒数和结实率8个性状的自交衰退在6个F2代群体中均达显著(P<0.05,下同)或极显著(P<0.01,下同)水平,播始历期、1~2穗节长和着粒密度等3个性状的自交衰退在部分F2代群体中达显著或极显著水平。正反交组合F2代群体中播始历期、株高、剑叶宽、有效穗数、实粒数和结实率6个性状的杂种优势和自交衰退均随海拔的升高而减小,剑叶长、主穗长、1~2穗节长、穗颈长和着粒密度5个性状的杂种优势和自交衰退在正反交组合中具有明显的差异。F2代群体生物学性状的中亲优势率、超亲优势率和自交衰退率明显受到产生F2群体的海拔和细胞质背景的影响。【结论】海拔和细胞质背景对水稻F2代群体杂种优势及自交衰退产生的效应,分别是由于海拔导致的环境温度变化对杂种F1代的雄配子选择以及不同细胞质背景下由雄配子和雌配子共同选择造成。此外,海拔对F2代群体当代的孢子体同样具有非常明显的效应。
关键词:籼粳稻细胞质;海拔变异;F2代群体;杂种优势;自交衰退
中图分类号:S511.035.1 文献标志码:A 文章编号:2095-1191(2023)02-0434-10
Abstract:【Objective】To study influence of environmental temperature changes due to different altitudes on selection effect of F1 generation gamete genotype under indica-japonica rice cytoplasmic background and advantage and depression of progeny separation population, in order to provide theoretical reference for rice breeding and study on effects of global temperature change on rice genetic variation. 【Method】N34, an improved japonica variety with indica-japonica cytoplasmic background, and XMG, an old typical japonica rice variety of strong cold tolerance, were taken as parents. Six F2 populations were generated from reciprocal hybrid of F1 generations at 3 different altitudes, and 11 biological traits of hete-rosis and inbreeding depression were analyzed. 【Result】Inbreeding depression of 8 traits such as plant height,flag-leaf length,flag-leaf width,effective panicle number,main panicle length,panicle neck length,filled grain number and seed setting rate were significant (P<0.05, the same below) or extremely significant (P<0.01, the same below) in 6 F2 populations. Inbreeding depression of 3 traits such as duration from sowing to heading, 1st-2nd panicle node length and grain density were significant or extremely significant in some F2 populations. In reciprocal hybrid combinations of F2 populations, heterosis rates and inbreeding depression of 6 traits in F2 population,such as duration from sowing to heading,plant height,flag-leaf width,effective panicle number,filled grain number and seed setting rate,decreased as altitude of F2 population generation increased. Variation trend of heterosis performance and inbreeding depression rate of other 5 traits including flag-leaf length,main panicle length,1th-2nd panicle node length,panicle neck length and grain density with altitude were greatly different in reciprocal hybrid combinations. Among biological traits of the F2 population, mid-parent hete-rosis rate,super-parent heterosis rate and inbreeding depression were greatly affected by altitude and cytoplasmic background of F2 population. 【Conclusion】Effects of altitude and cytoplasmic background on heterosis and inbreeding depression of F2 population in rice are caused by hybrid F1 male gamete selection due to environmental temperature change at different altitudes,and co-selection of male and female gametes under different cytoplasmic backgrounds,respectively. In addition,altitude also greatly affects contemporary sporophytes of F2 population.
Key words: Indica-japonica rice cytoplasm; altitude variation; F2 population; heterosis; inbreeding depression
Foundation items: Yunnan Key New Products Development Plan Project(2015BB015)
0 引言
【研究意义】生物的整个生活史均处于自然条件的选择压力中,由于全球气候的变异加剧,生物正面临着以气温变化为主要特征的气候条件变化的新选择。气温对植物孢子体世代具有非常明显的影响(Tanto and Demissie,2000;赵利锋等,2001;马丹炜等,2006;严学兵等,2006;张锦文等,2017)。高海拔极端环境的强选择压力能够极大地促进物种表型和遗传适应发生变化(郝艳和雷富民,2022),然而生物对环境的遗传反应仍知之甚少。水稻作为我国第一大粮食作物,其杂种优势利用最为成功,对世界粮食增产贡献巨大。然而,由于水稻的生长、遗传分化、种质资源分布等均受到以气温为主的生态环境的严重影响,故气候变化对其产生的影响直接关系我国的粮食安全。从水稻的分布和前人研究结果可推断出由海拔引起的温度变化是导致水稻籼粳分化的一个重要因素(Nagamine et al.,1992;李自超等,2001;曾亚文等,2001)。尽管Rood等(1988)研究认为杂种优势与环境温度无关,但后续的研究表明不同生态环境条件下F1代产量性状的杂种优势或生活力表现程度有明显的差异(徐美兰,2011;姜高伟等,2021)。因此,研究海拔引起的环境温度变化对籼粳稻细胞质背景下的F1代配子基因型的选择效应及其对后代群体优势的影响,对水稻育种研究和全球气温变化对水稻遗传变异的效应探讨具有参考意义。【前人研究进展】在高等植物的生活史中,二倍体的孢子体世代和单倍体的配子体世代均处于自然条件的选择压力中。在配子产生过程中及配子与配子结合之前,适者生存的法则就对配子发挥着选择作用,只有最适于环境条件的配子才能通过竞争性受精从而产生合子,具有完整遗传信息的配子体经选择后可影响新一代孢子体的表现(Mulcahy,1979;Wang et al.,2009)。海拔环境差异对水稻杂合体雌配子和雄配子发生的选择,会使其后代基因型频率、遗传分化、生物学性状表现等方面发生明显变化(Wang et al.,2009;雷伟等,2015)。籼型和粳型2个亚种无论在形态和生理上还是对环境的适应性方面均存在巨大的差异,由于籼稻和粳稻基因型在不同生态条件下的繁殖速度和死亡率的差异以及由于花粉传播特性不同,籼粳杂种半不育造成的配子选择致使籼粳在遗传上发生明显的分化(杨忠义等,2007)。海拔与云南稻种的籼粳分化密切相关,是籼粳分化的驱动力(曾亚文等,2001;邰丽梅等,2006)。自袁隆平(1987)提出籼粳杂种优势利用的战略构想以来,我国水稻遗传学家长期致力于籼粳杂种不育、广亲和、杂种优势利用等方面的研究(米甲明和欧阳亦聃,2022)。籼粳杂交水稻品种的杂种优势很明显(刘杰和黃学辉,2021),籼粳杂交组合的双亲形态学与基因组的籼粳分化差异均是产生籼粳杂种优势的基础(孙传清等,2000)。然而,籼粳杂种F1代存在明显的不育现象,在F2代及其后代表现有杂种衰败现象,即生殖隔离限制了籼粳杂交后代杂种优势的直接利用(李荣德,2016)。水稻细胞质对F1代存在明显的效应(康公平等,2017),籼型或粳型细胞核中带入的细胞质在进化程度中越低级细胞质效应就越明显(汤述翥等,2003)。前人对水稻(唐雪松和刘平,2013)、玉米(宋方威等,2011;Fan et al.,2014)、菊花(奥妮等,2020)等植物也进行相关研究,结果显示一些性状的杂种优势在正反交组合间存在显著差异,表明细胞质背景对杂种优势有影响。【本研究切入点】杂种优势或自交衰退均是在孢子体世代进行环境胁迫或改变遗传背景所产生,目前未见海拔变化和细胞质背景所产生的配子体选择对后代生物学性状杂种优势及自交衰退影响的研究报道。【拟解决的关键问题】利用籼粳不同细胞质背景的材料进行正反交,并将其F1代种植在高、中、低3个不同海拔条件下得到6个F2代群体,以亲本、F1代和6个F2代群体为材料,种植在中等海拔的同一环境条件,对生物学性状的杂种优势和自交衰退进行比较分析,以明确海拔和细胞质背景对水稻F2代群体杂种优势及自交衰退的影响,进而探究海拔导致的环境温度变化以及不同细胞质背景对杂种F1的雌雄配子选择效应,为水稻育种和探讨全球气温变化对水稻遗传变异的效应提供参考。
1 材料与方法
1. 1 试验材料
试验材料包括小麻谷、南34、小麻谷与南34正反交F1代及其在元江(海拔400 m)、昆明(海拔1860 m)、西山团结(海拔2200 m)3个不同海拔产生的6个F2代群体。小麻谷为云南省宁蒗县永宁乡海拔2650 m高寒稻区种植的地方粳稻老品种,南34为籼粳复合杂交选育出的具有籼稻细胞质背景的偏粳改良品系(洪汝科等,2004)。3个试验点海拔最大相差1800 m,纬度最大相差1°31',经度最大相差0°57',年均温最大相差10.6 ℃,极端最高气温最大相差16.1 ℃;水稻抽穗期的平均温度相差10.1 ℃,平均最高温相差13.9 ℃(Wang et al.,2009)。
1. 2 种植管理
3月13日在云南农业大学稻作研究所试验田(昆明)播种不同海拔下产生的6个F2代群体,5月4日移栽,种植于昆明试点的同一块试验田中,全部材料单本插,株行距为15 cm×20 cm,每行10株,管理条件同大田。
1. 3 调查性状
成熟时,按单株取样调查播始历期(播种至开始抽穗所用天数,d)、株高(cm)、剑叶长(cm)、剑叶宽(cm)、有效穗数(穗/株)、主穗长(cm)、1~2穗节长(cm)、穗颈长(cm)、总粒数(粒/穗)、结实率(%)、着粒密度等11个生物学性状。亲本小麻谷调查了9株,亲本南34和正反交F1代各调查了10株,F2代群体调查全部单株(表1)。
1. 4 统计分析
以中亲优势率和超亲优势率进行F1代和F2代杂种优势分析,以自交衰退率进行F2代群体自交衰退分析。F1代群体和F2代群体的中亲优势率和超亲优势率,以及F2代群体的中亲优势自交衰退率和超亲优势自交衰退率按以下公式计算:
F1代中亲优势率(MHF1,%)= [F1-MPMP][×]100
F1代超亲优势率(HbF1,%)= [F1-BPBP][×]100
F2代中亲优势率(MHF2,%)= [F2-MPMP][×]100
F2代超亲优势率(HbF2,%)= [F2-BPBP][×]100
F2代中亲优势自交衰退率(MIDF2,%)= [F2-F1MP][×]100
F2代超亲优势自交衰退率(HbIDF2,%)= [F2-F1BP][×]100
式中,[F1]、[MP]、[BP]和[F2]分別代表杂交F1代群体性状均值、双亲性状均值、超高亲本性状均值及F2代群体性状均值。
以杂种优势或自交衰退的绝对值进行显著性测验(Soehenei and Srinives,2005)。例如,F1代中亲优势的显著性测验如下:
式中,S、V、P1、P2分别表示标准差、方差、亲本1测定值和亲本2测定值。当t≥t0.05或t≥t0.01时,优势达显著或极显著水平。显著性测验的自由度分别为(n1-1)+(n2-1)+(n3-1),其中n1、n2和n3分别代表杂交组合取样数量、亲本1取样数量及亲本2取样数量。其他类型优势均按此方法更换公式进行估算。
2 结果与分析
2. 1 正反交F1代生物学性状的杂交优势率分析结果
各生物学性状在南34与小麻谷F1代正反交群体之间表现出不同程度的杂种优势或杂种劣势(表2)。在正交组合(南34×小麻谷)中,F1代的株高、剑叶宽、有效穗数、主穗长和实粒数5个性状均显著(P<0.05,下同)或极显著(P<0.01,下同)超过双亲均值;株高、剑叶宽、1~2穗节长、穗颈长、实粒数和结实率6个性状与超高亲本存在达显著或极显著差异,其中,剑叶宽和实粒数显著高于超高亲本,其余4个性状极显著低于超高亲本;播始历期、剑叶长和着粒密度等性状与双亲均值和超高亲本均无显著差异(P>0.05,下同)。在反交组合(小麻谷×南34)中,F1代的播始历期、株高、剑叶宽、主穗长、实粒数和结实率6个性状与双亲均值存在达显著或极显著差异,其中播始历期极显著低于双亲均值,其余5个性状均显著或极显著高于双亲均值;播始历期、株高、剑叶长、1~2穗节长、穗颈长和实粒数6个性状与超高亲本均达显著或极显著差异,其中剑叶长和实粒数显著高于超高亲本,其余4个性状显著或极显著低于超高亲本;有效穗数和着粒密度等性状与双亲均值和超高亲本均无显著差异。正反交组合表现一致的是,F1代的株高、1~2穗节长、穗颈长显著或极显著低于超高亲本,实粒数则显著高于超高亲本,有效穗数、主穗长和着粒密度等性状与超高亲本无显著差异。综上所述,南34与小麻谷杂交F1代各生物学性状的优势表现具有明显差异,并且受到细胞质背景的影响。
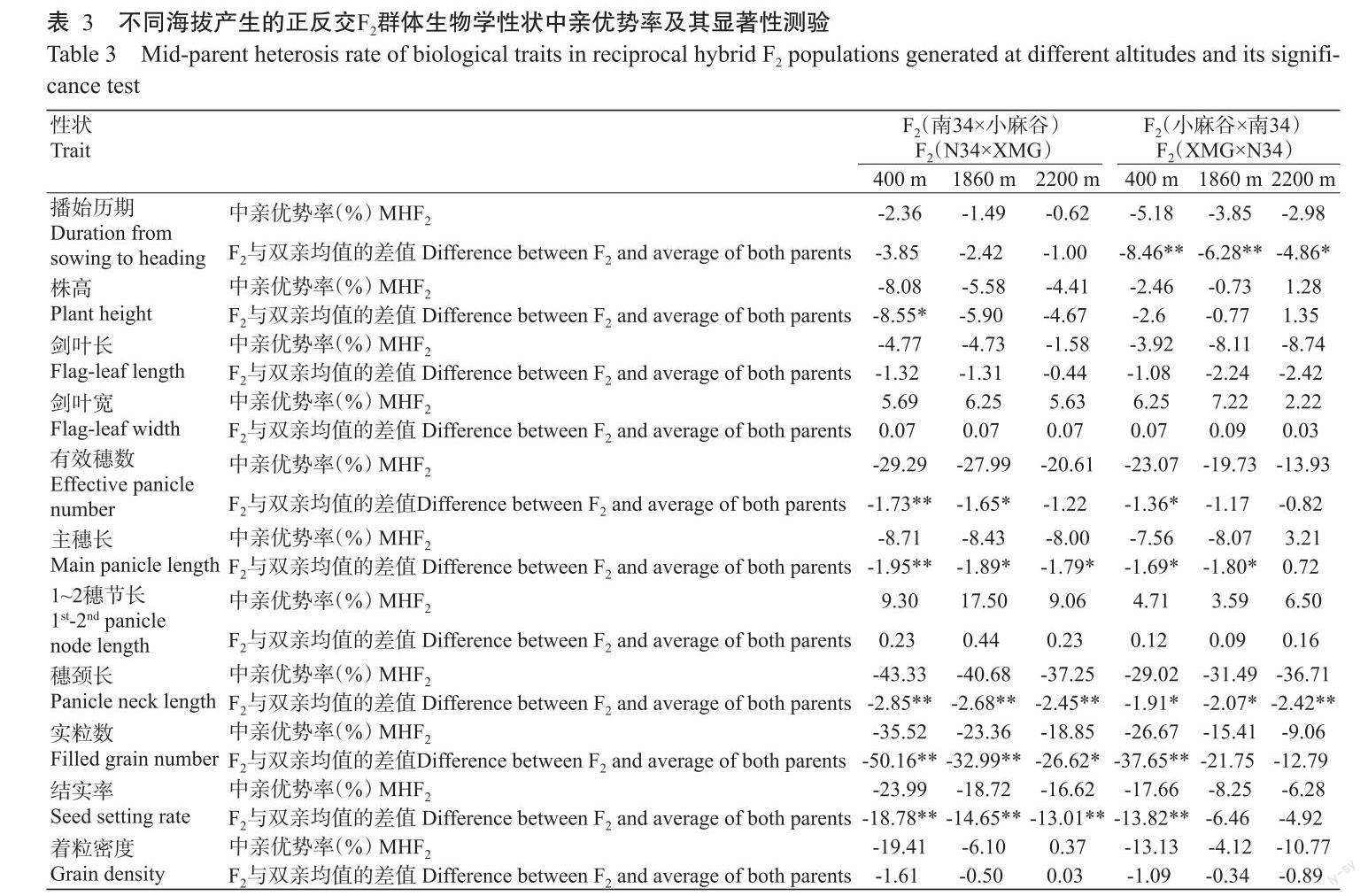
2. 2 不同海拔产生的正反交F2代群体生物学性状中亲优势分析结果
相同组合在不同海拔产生的F2代群体表现出具有明显差异的中亲优势,正反交组合在相同海拔产生的F2群体同样表现出具有明显差异的中亲优势(表3)。
在正交组合(南34×小麻谷)中,3个海拔条件下产生的F2代群体播始历期、株高、剑叶长、有效穗数、主穗长、穗颈长、实粒数和结实率8个性状均表现出负向的中亲优势,剑叶宽和1~2穗节长均表现出正向的中亲优势,着粒密度在400和1860 m海拔下产生的F2代群体中表现出负向的中亲优势,在2200 m海拔产生的F2代群体中表现出正向的中亲优势。3个海拔条件下,播始历期、剑叶长、剑叶宽、1~2穗节长和着粒密度5个性状的中亲优势均未达显著水平,主穗长、穗颈长、实粒数和结实率4个性状的中亲优势均达显著或极显著水平;然而,株高和有效穗数在不同海拔产生的F2代群体中表现出的中亲优势明显不同,株高仅在400 m海拔产生的F2代群体中所表现出的中亲优势达显著水平,在1860和2200 m海拔产生的F2代群体中表现出的中亲优势未达显著水平,有效穗数在400和1860 m海拔产生的F2代群体中表现出的中亲优势分别达极显著和显著水平,在2200 m海拔产生的F2代群体中所表现出的中亲优势未达显著水平。
在反交组合(小麻谷×南34)中,3个海拔条件产生的F2代群体其播始历期、剑叶长、有效穗数、穗颈长、实粒数、结实率和着粒密度等7个性状均表现出负向的中亲优势,剑叶宽和1~2穗节长均表现出正向的中亲优势,株高和主穗长在400和1860 m海拔产生的F2群体中表现出负向的中亲优势,在2200 m海拔产生的F2代群体中表现出正向的中亲优势。其中,株高、剑叶长、剑叶宽、1~2穗节长和着粒密度等5个性状的中亲优势均未达显著水平,播始历期和穗颈长的中亲优势均达显著或极显著水平;然而,有效穗数、主穗长、实粒数和结实率4个性状在不同海拔产生的F2群体中所表现出的中亲优势明显不同,有效穗数、实粒数和结实率仅在400 m海拔产生的F2代群体中所表现出的中亲优势达显著水平,在1860和2200 m海拔产生的F2代群体中表现出的中亲优势未达显著水平,主穗长在400和1860 m海拔产生的F2代群体中表现出的中亲优势均达显著水平,在2200 m海拔产生的F2代群体中表现出的中亲优势未达显著水平。
在正反交组合之间,F2代群体播始历期、株高、剑叶宽、有效穗数、实粒数和结实率6个性状中亲优势率绝对值的变化趋势比较一致,即播始历期、株高、有效穗数、实粒数和结实率5个性状的中亲优势率绝对值随海拔的升高而逐渐减小;剑叶宽的中亲优势率随海拔的升高呈先增大后减小的趋势。然而,剑叶长、主穗长、1~2穗节长、穗颈长和着粒密度5个性状中亲优势率的变化趋势在正反交组合之间具有明显差异。例如,在正交组合(南34×小麻谷)中,F2代群体剑叶长、主穗长、穗颈长和着粒密度4个性状的中亲优势率绝对值随海拔的升高而逐渐减小,1~2穗节长的中亲优势率随海拔的升高呈先增大后减小的趋势;但在反交组合(小麻谷×南34)中,F2代群体剑叶长和穗颈长的中亲优势率绝对值随海拔的升高而逐渐增大,主穗长的中亲优势率绝对值随海拔的升高呈先增大后减小的趋势,1~2穗节长和着粒密度的中亲优势率绝对值随海拔的升高呈先减小后增大的趋势。
综上所述,F2代群体中生物学性状的中亲优势率明显受到产生F2代群体的海拔和细胞质背景的影响,其中细胞质背景对剑叶长、主穗长、1~2穗节长、穗颈长和着粒密度等5个性状的影响非常突出。
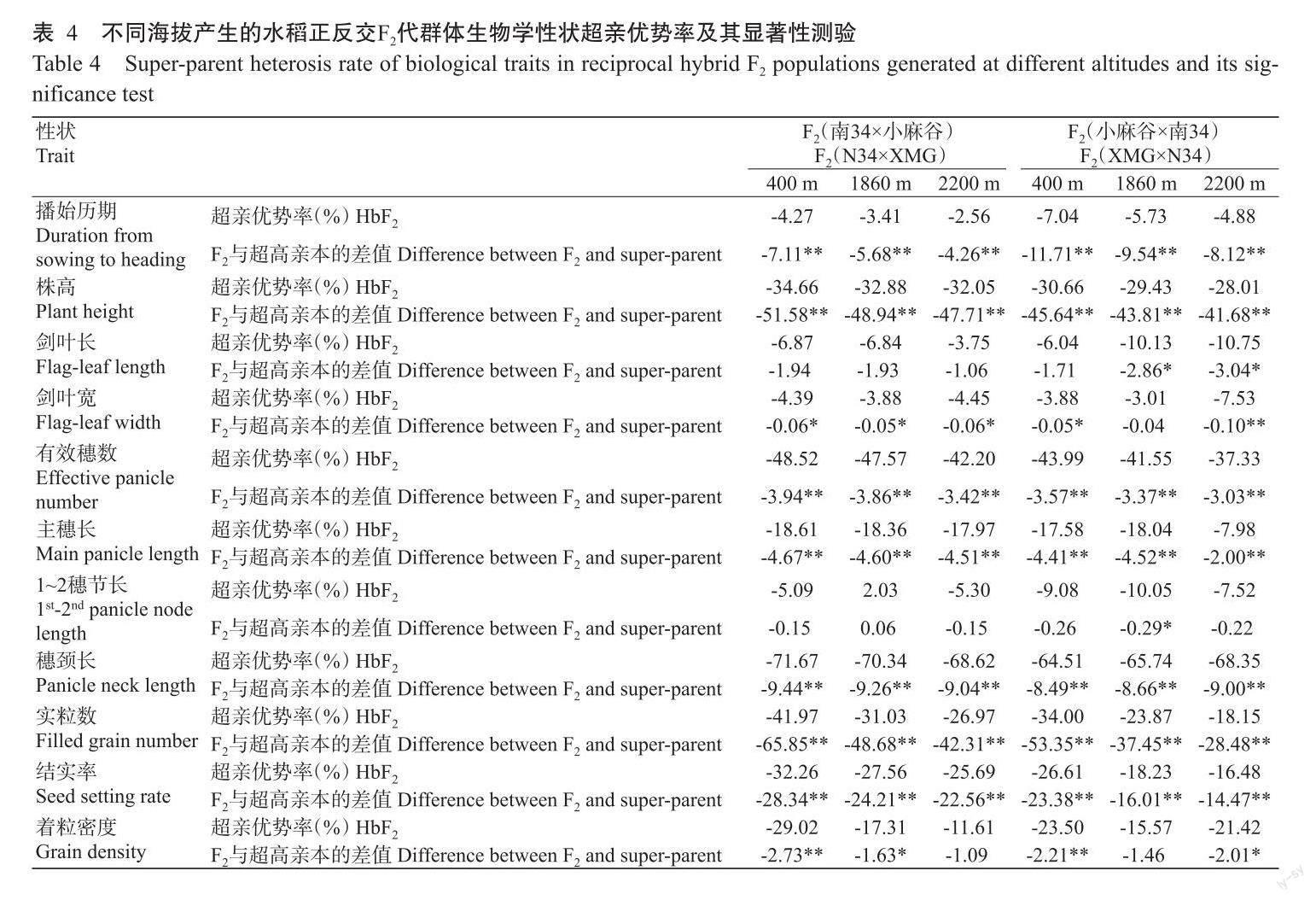
2. 3 不同海拔产生的正反交F2代群体生物学性状超亲优势分析结果
相同组合在不同海拔产生的F2代群体表现出具有明显差异的超亲优势,正反交组合在相同海拔产生的F2代群体同样表现出具有明显差异的超亲优势(表4)。
6个F2代群体中,11个性状几乎均表现出负向的超亲优势,仅在1860 m海拔产生的F2群体中1~2穗节长表现出正向的超亲优势。播始历期、株高、剑叶宽、有效穗数、主穗长、穗颈长、实粒数和结实率8个性状的超亲优势均达显著或极显著水平;然而,剑叶长、1~2穗节长和着粒密度3个性状的超亲优势显著性水平在不同F2代群体中存在明显差异。例如,在正交组合(南34×小麻谷)中剑叶长和1~2穗节长未达显著水平,着粒密度在400和1860 m海拔产生的F2群体中超亲优势分别达极显著和显著水平;但在反交组合(小麻谷×南34)中,剑叶长在1860和2200 m海拔产生的F2群体中超亲优势均达显著水平,1~2穗节长在1860 m海拔产生的F2代群体中超亲优势达显著水平,着粒密度在400和2200 m海拔产生的F2群体中超亲优势分别达极显著和显著水平。
在正反交组合之间,F2代群体播始历期、株高、剑叶宽、有效穗数、实粒数和结实率6个性状超亲优势率绝对值的变化趋势比较一致,即播始历期、株高、有效穗数、实粒数和结实率5个性状的超亲优势率绝对值随海拔的升高而逐渐减小;剑叶宽的中亲优势率绝对值随海拔的升高而减小,在1860 m海拔以上随海拔的升高而明显增大。然而,剑叶长、主穗长、1~2穗节长、穗颈长和着粒密度5个性状超亲优势率的变化趋势在正反交组合之间具有明显差异。例如,在正交组合(南34×小麻谷)的杂交组合中,F2代群体剑叶长、主穗长、穗颈长和着粒密度4个性状的超亲优势率绝对值随海拔的升高而逐渐减小,1~2穗节长的超亲优势率绝对值随海拔的升高呈先减小后增大的趋势;但在反交组合(小麻谷×南34)中,F2代群体剑叶长和穗颈长的超亲优势率绝对值随海拔的升高而逐渐增大,主穗长和1~2穗节长的超亲优势率绝对值随海拔的升高呈先增大后减小的趋势,着粒密度的超亲优势率绝对值随海拔的升高呈先减小后增大的趋势。
综上所述,F2代群体中生物学性状的超亲优势率同样明显受到产生F2代群体的海拔和细胞质背景的影响,其中细胞质背景对剑叶长、主穗长、1~2穗节长、穗颈长和着粒密度5个性状的影响非常突出。
2. 4 不同海拔产生的正反交F2代群体生物学性状自交衰退率分析结果
在不同海拔产生的正反交组合F2代群体中除了1~2穗节长外,其余各性状均表现出一定程度的自交衰退现象(表5)。6个F2代群体中,株高、剑叶长、剑叶宽、有效穗数、主穗长、穗颈长、实粒数和结实率8个性状的自交衰退均达显著或极显著水平;1~2穗节长均大于F1代群体,甚至在400和2200 m海拔产生的正交组合(南34×小麻谷)F2代群体中差异达极显著水平,表明1~2穗节长在F2代群体中不仅没有衰退反而还产生了优势;着粒密度均表现出不同程度的自交衰退,在400和1860 m海拔产生的正交组合(南34×小麻谷)F2代群体以及在400和2200 m海拔产生的反交组合(小麻谷×南34)F2群体中自交衰退达极显著水平。播始历期在400 m海拔产生的正交组合(南34×小麻谷)F2代群体中表现值小于F1代群体,但差异不显著;然而,在其余5个F2代群体中均大于F1代群体,并且在2200 m海拔产生的正交组合(南34×小麻谷)F2群体以及在1860和2200 m海拔产生的反交组合(小麻谷×南34)F2群体中差异达显著或极显著水平。在正反交组合之间产生的F2代群体中,播始历期、株高、有效穗数、实粒数和结实率等5个性状的自交衰退率均随海拔的升高而减小;剑叶宽的自交衰退率随海拔的升高呈先减小后增大的趋势。然而,剑叶长、主穗长、穗颈长和着粒密度4个性状的自交衰退率变化趋势在正反交组合之间具有非常明显的差异,例如,在正交组合(南34×小麻谷)中这4个性状的自交衰退率均随海拔的升高而减小;然而,在反交组合(小麻谷×南34)中剑叶长和穗颈长的自交衰退率却随海拔的升高而增大,主穗长的自交衰退率随海拔的升高呈先增大后减小的趋势,着粒密度的自交衰退率随海拔的升高呈先减小后增大的趋势。
综上所述,F2代群体中生物学性状的自交衰退率也明显受到产生F2代群体的海拔和细胞质背景的影响,其中细胞质背景对剑叶长、主穗长、穗颈长和着粒密度等4個性状的影响非常突出。
3 讨论
作物杂种优势除了受到遗传基因的影响外,还明显受到外在的环境条件以及两者相互作用的影响(杨成明等,2009),本研究也得出相同结论。F2代群体中有6个性状的杂种优势和自交衰退在正反交组合中均随产生F2代群体海拔的升高而减小,说明海拔明显影响F2代群体生物学性状的杂种优势和自交衰退表现。由于海拔极显著影响F1代的花粉育性,而对其小穗育性的影响不明显(吕川根等,2002;Wang et al.,2009),因此,这种影响可以判定主要是由于海拔导致的环境温度变化对杂种F1代的雄配子选择造成,而不存在明显的雌配子选择。但本研究发现,F2代群体中另外5个性状的杂种优势表现和自交衰退在正反交组合中具有明显的差异,说明籼粳细胞质也同样明显影响着生物学性状的杂种优势和自交衰退表现。由于在3个海拔条件下籼粳细胞质均显著或极显著影响着花粉育性和小穗育性(Wang et al.,2009),故推测正反交组合F2代群体间生物学性状的杂种优势表现和自交衰退差异除了由雄配子选择引起外,应该还有雌配子的选择所造成,当然由于F1代的小穗育性均在78%以上,雌配子选择程度相对较小。
植物群體的遗传变异在地理分布和海拔差异下明显不相同,海拔引起的以温度为主的生态因子变化对生物遗传多样和遗传分化起着重要作用(冯富娟等,2004;李珊等,2004;姜志磊等,2005;高丽和杨波,2006)。海拔对水稻的遗传分化同样有着非常明显的影响(曾亚文等,2001),由于海拔的不同,稻种资源植株的形态性状发生极为明显的变异。本研究将水稻杂合体置于不同海拔条件下仅繁殖了一代,即可导致不同海拔产生的F2代群体间杂种优势和自交衰退情况有较大变异。当然,若将这6个F2群体体种植在2200 m的高海拔低温条件下(王石华和谭学林,2015),各性状的优势和衰退表现与本研究种植在中海拔较适温度条件下也有非常明显的差异,例如种植在高海拔低温条件下F2代群体株高、有效穗数、主穗长等性状的自交衰退率随海拔的升高而增大,但种植在中海拔较适温度条件下这些性状的自交衰退率却随产生F2代群体海拔的升高而减小。由此可见,海拔差异引起的温度差异除了对产生F2代群体的雌雄配子体世代有非常明显的选择以外,对F2代群体孢子体世代同样具有非常明显的影响。然而,由于本研究仅使用F2代群体进行研究,故难以明确海拔和籼粳细胞质背景对雌雄配子体各自的选择程度,也难以明确海拔对孢子体世代的选择程度。今后可同时选择亲本、F1、F2以及回交群体来进行研究,为探讨全球气温变化对水稻遗传变异的效应提供更有价值的参考。
4 结论
海拔与细胞质背景对水稻F2代群体杂种优势及自交衰退产生的效应,分别是由于海拔导致的环境温度变化对杂种F1的雄配子选择以及不同细胞质背景下由雄配子和雌配子共同选择造成。此外,海拔对F2群体当代的孢子体同样具有非常明显的效应。
参考文献:
奥妮,仰小东,李晨曦,于瑞宁,房伟民,陈发棣,张飞. 2020. 菊花正反交F1代株高和叶形相关性状的遗传差异[J]. 分子植物育种,18(17):5852-5859. [Ao N,Yang X D,Li C X,Yu R N,Fang W M,Chen F D,Zhang F. 2020. Genetic divergence of plant height and leaf shape related traits in reciprocal F1 progenies of chrysanthemum[J]. Molecular Plant Breeding,18(17):5852-5859.] doi:10. 13271/j.mpb.018.005852.
冯富娟,王凤友,李长松. 2004. 长白山不同海拔条件下红松的遗传分化[J]. 东北林业大学学报,32(3):1-3. [Feng F J,Wang F Y,Li C S. 2004. Genetic differentiation of Pinus koraiensis under different altitude conditions in Changbai Mountains[J]. Journal of Northeast Forestry University,32(3):1-3.] doi:10.3969/j.issn.1000-5382.2004. 03.001.
高丽,杨波. 2006. 湖北野生春兰资源遗传多样性的ISSR分析[J]. 生物多样性,14(3):250-257. [Gao L,Yang B. 2006. Genetic diversity of wild Cymbidium goeringii (Orchidaeae) populations from Hubei based on ISSR analysis[J]. Biodiversity Science,14(3):250-257.] doi:10. 3321/j.issn:1005-0094.2006.03.008.
郝艳,雷富民. 2022. 适应性演化的分子遗传机制:以高海拔适应为例[J]. 遗传,44(8):635-654. [Hao Y,Lei F M. 2022. Genetic mechanism of adaptive evolution:The example of adaptation to high altitudes[J]. Hereditas,44(8):635-654.] doi:10.16288/j.yczz.22-108.
洪汝科,李铮友,王樨,师常俊,黄大军,谭学林. 2004. 抗病优质滇型粳稻恢复系南34的选育及应用[J]. 西南农业学报,17(S):85-87. [Hong R K,Li Z Y,Wang X,Shi C J,Huang D J,Tan X L. 2004. Breeding and application of Dian type restorer Nan 34 with good quality and disease-resistance[J]. Southwest China Journal of Agricultural Sciences,17(S):85-87.] doi:10.3969/j.issn.1001-4829. 2004.z1.020.
姜高伟,李琪,徐成勋,刘士凯. 2021. 温度对长牡蛎和葡萄牙牡蛎及其杂交稚贝生长和存活的影响[J]. 中国水产科学,28(1):29-36. [Jiang G W,Li Q,Xu C X,Liu S K. 2021. Effects of temperature on the growth and survival of reciprocal hybrids of two oyster species,Crassostrea gigas and Crassostrea angulata[J]. Journal of Fishery Sciences of China,28(1):29-36.]
姜志磊,杨欣明,王瑞,高爱农,李立会. 2005. 基于SSR的梭罗草遗传多样性分析[J]. 植物遗传资源学报,6(3):315-318. [Jiang Z L,Yang X M,Wang R,Gao A N,Li L H. 2005. Genetic diversity of Roegneria thoroldiana (Oliv.) Keng populations based on SSR analyses[J]. Journal of Plant Genetic Resources,6(3):315-318.] doi:10.13430/j.cnki.jpgr.2005.03.015.
康公平,戴小军,欧立军,梁满中,陈良碧. 2017. 同核异质两系杂交水稻主要农艺性状的细胞质效应研究[J]. 杂交水稻,32(4):63-66. [Kang G P,Dai X J,Ou L J,Liang M Z,Chen L B. 2017. Cytoplasmic effects on major agronomic traits of isonuclear alloplasmic two-line rice hybrids[J]. Hybrid Rice,32(4):63-66.]
雷伟,文建成,普世皇,王昌江,刘华萍,孙朝华,苏家秀,李振,徐津,谭亚玲,金寿林,谭学林. 2015. 海拔差异对水稻雌配子体基因型选择及其后代表型遗传分化的影响[J]. 中国农业科学,48(7):1249-1261. [Lei W,Wen J C,Pu S H,Wang C J,Liu H P,Sun C H,Su J X,Li Z,Xu J,Tan Y L,Jin S L,Tan X L. 2015. Female gametophyte genotype selection and its progeny phenotypic genetic differentiation in rice at different altitude condition[J]. Scientia Agricultura Sinica,48(7):1249-1261.] doi:10.3864/j.issn.0578-1752.2015.07.01.
李榮德. 2016. 水稻籼粳亚种间杂种衰败的遗传解析及HB-12基因克隆[D]. 扬州:扬州大学. [Li R D. 2016. Genetic dissection of hybrid breakdown in an Indica/Japonica cross in rice (Oryza sativa L.) and molecular cloning of HB-12[D]. Yangzhou:Yangzhou University.]
李珊,蔡宇良,钱增强,赵桂仿. 2004. 云南金钱槭形态变异与遗传变异的相关性研究[J]. 生态学报,24(5):925-931. [Li S,Cai Y L,Qian Z Q,Zhao G F. 2004. Researches on the relationship between the morphological and genetic variations of Dipteronia dyeriana Henry(Aceraceae)[J]. Acta Ecologica Sinica,24(5):925-931.] doi:10.3321/j.issn:1000-0933.2004.05.010.
李自超,张洪亮,曾亚文,申时全,孙传清,王象坤. 2001. 云南稻种资源表型遗传多样性的研究[J]. 作物学报,27(5):832-837. [Li Z C,Zhang H L,Zeng Y W,Shen S Q,Sun C Q,Wang X K. 2001. Studies on phenotypic diversity of rice germplasm in Yunnan,China [J]. Acta Agronomica Sinica,27(5):832-837.] doi:10.3321/j.issn:0496-3490. 2001.06.023.
刘杰,黄学辉. 2021. 作物杂种优势研究现状与展望[J]. 中国科学:生命科学,51(10):1396-1404. [Liu J,Huang X H. 2021. Advances and perspectives in crop heterosis[J]. Scientia Sinica(Vitae),51(10):1396-1404.] doi:10.1360/SSV-2021-0171.
吕川根,王才林,宗寿余,赵凌,邹江石. 2002. 温度对水稻亚种间杂种育性及结实率的影响[J]. 作物学报,28(4):499-504. [Lü C G,Wang C L,Zong S Y,Zhao L,Zou J S. 2002. Effects of temperature on fertility and seed set in intersubspecific hybrid rice (Oryza sativa L.)[J]. Acta Agronomica Sinica,28(4):499-504.] doi:10.3321/j.issn:0496-3490.2002.04.012.
马丹炜,王胜华,罗通,王文国,庄国庆,陈放. 2006. 环境因子对岩生植物金发草遗传多样性的影响[J]. 中山大学学报(自然科学版),45(2):73-77. [Ma D W,Wang S H,Luo T,Wang W G,Zhuang G Q,Chen F. 2006. Effects of environmental factors on the genetic diversity of Pogonatherum paniceum[J]. Acta Scientiarum Naturalium Universitatis Sunyatseni,45(2):73-77.] doi:10.3321/j.issn:0529-6579.2006.02.018.
米甲明,欧阳亦聃. 2022. 水稻籼粳亚种间杂种优势利用的理论与实践[J]. 华中农业大学学报,41(1):40-47. [Mi J M,Ouyang Y D. 2022. Theory and practice of utilizing Xian-geng inter-subspecific heterosis in rice[J]. Journal of Huazhong Agricultural University,41(1):40-47.] doi:10.13300/j.cnki.hnlkxb.2022.01.004.
宋方威,彭惠茹,刘婷,张义荣,孙其信,倪中福. 2011. 利用三重测交群体剖析玉米株高与穗位高杂种优势的遗传学基础[J]. 作物学报,37(7):1186-1195. [Song F W,Peng H R,Liu T,Zhang Y R,Sun Q X,Ni Z F. 2011. Heterosis for plant height and ear position in maize revealed by quantitative trait loci analysis with triple testcross design[J]. Acta Agronomica Sinica,37(7):1186-1195.]
孙传清,姜廷波,陈亮,吴长明,李自超,王象坤. 2000. 水稻杂种优势与遗传分化关系的研究[J]. 作物学报,26(6):641-649. [Sun C Q,Jiang T B,Chen L,Wu C M,Li Z C,Wang X K. 2000. Studies on the relationship between heterosis and genetic differentiation in hybrid rice (Oryza sativa L.)[J]. Acta Agronomica Sinica,26(6):641-649.] doi:10.3321/j.issn:0496-3490.2000.06.001.
邰丽梅,杜鹃,曾亚文,李自超,张洪亮,普晓英,杨树明,凌青. 2006. 思茅地区核心稻种籼粳特异性标记的聚类分析[J]. 西南农业学报,19(S1):49-53. [Tai L M,Du J,Zeng Y W,Li Z C,Zhang H L,Pu X Y,Yang S M,Ling Q. 2006. The cluster analysis of peculiarity of Indica and Japonica in Simao[J]. Southwest China Journal of Agricultural Sciences,19(S1):49-53.] doi:10.3969/j.issn.1001-4829.2006.z1.009.
汤述翥,张亚东,孙红芹,叶全宝,陈秀花,顾铭洪. 2003. 水稻同核异质广亲和不育系细胞质效应的研究[J]. 作物学报,29(2):202-207. [Tang S Z,Zhang Y D,Sun H Q,Ye Q B,Chen X H,Gu M H. 2003. Effects of CMS cytoplasm in isonuclear alloplasmic CMS lines with wide compatibility in rice[J]. Acta Agronomica Sinica,29(2):202-207.] doi:10.3321/j.issn:0496-3490.2003.02.007.
唐雪松,刘平. 2013. 水稻细胞质效应对杂种优势的影响研究[J]. 种子,32(6):36-42. [Tang X S,Liu P. 2013. Study the influence of cytoplasm effects on heterosis in rice[J]. Seed,32(6):36-42.] doi:10.16590/j.cnki.1001-4705. 2013.06.061.
王石華,谭学林. 2015. 不同海拔产生的水稻F2群体杂种优势及自交衰退研究[J]. 西南农业学报,28(1):16-20. [Wang S H,Tan X L. 2015. Study on heterosis and inbreeding depression in rice F2 populations generated from hybrids at different altitudes[J]. Southwest China Journal of Agricultural Science,28(1):16-20.] doi:10.16213/j.cnki.scjas.2015.01.004.
徐美兰. 2011. 寒地粳稻产量性状杂种优势环境变异及遗传机理研究[D]. 哈尔滨:东北农业大学. [Xu M L. 2011. Study on the environmental variation and genetic mechanism of heterosis on yield traits of Japonica rice in cold region[D]. Harbin:Northeast Agricultural University.]
严学兵,郭玉霞,周禾,王堃. 2006. 影响披碱草属植物遗传分化和亲缘关系的地理因素分析[J]. 植物资源与环境学报,15(4):17-24. [Yan X B,Guo Y X,Zhou H,Wang K. 2006. Analysis of geographical conditions affected on genetic variation and relationship among populations of Elymus[J]. Journal of Plant Resources and Environment,15(4):17-24.] doi:10.3969/j.issn.1674-7895.2006.04.003.
杨成明,吴孝波,刘勇强,潘琳,董绍斌,刘育生. 2009. 气温对杂交水稻II优718产量优势的影响分析[J]. 西南农业学报,22(5):1232-1237. [Yang C M,Wu X B,Liu Y Q,Pan L,Dong S B,Liu Y S. 2009. Effect of temperature on yield performance of hybrid rice II You 718[J]. Southwest China Journal of Agricultural Science,22(5):1232-1237.] doi:10.16213/j.cnki.scjas.2009.05.026.
杨忠义,曹永生,苏艳,刘晓利,李华慧,卢义宣. 2007. 中国栽培稻的籼粳分化机理再论[J]. 西南农业学报,20(3):321-326. [Yang Z Y,Cao Y S,Su Y,Liu X L,Li H H,Lu Y X. 2007. Polarization mechanism of Indica and Japonica rice in China cultivated rice[J]. Southwest China Journal of Agricultural Sciences,20(3):321-326.] doi:10.16213/j.cnki.scjas.2007.03.001.
曾亚文,李自超,杨忠义,申时全,张洪亮,王象坤. 2001. 云南地方稻种籼粳亚种的生态群分类及其地理生态分布[J]. 作物学报,27(1):15-20. [Zeng Y W,Li Z C,Yang Z Y,Shen S Q,Zhang H L,Wang X K. 2001. Geographical distribution and cline classification of Indica/Japonica subspecies of Yunnan local rice resources[J]. Acta Agronomica Sinica,27(1):15-20.] doi:10.3321/j.issn:0496-3490.2001.01.003.
袁隆平. 1987. 杂交水稻的育种战略设想[J]. 杂交水稻,(1):1-3. [Yuan L P. 1987. Breeding strategy of hybrid rice[J]. Hybrid Rice,(1):1-3.] doi:10.16267/j.cnki.1005-3956. 1987.01.001.
张锦文,邓伟,年伟,吕永刚,徐雨然,谷安宇,曾彪,李小林,邓华凤. 2017. 杂交籼稻在云南不同海拔和纬度下稻米淀粉RVA谱特征值分析[J]. 杂交水稻,32(3):59-65. [Zhang J W,Deng W,Nian W,Lü Y G,Xu Y R,Gu A Y,Zeng B,Li X L,Deng H F. 2017. Analysis on rice starch RVA spectrum eigenvalues of indica hybrid rice under different altitude and latitude conditions in Yunnan[J]. Hybrid Rice,32(3):59-65.] doi:10.16267/j.cnki. 1005-3956.201703022.
赵利锋,李珊,潘莹,阎桂琴,赵桂仿. 2001. 华山新麦草自然居群沿海拔梯度的遗传分化[J]. 西北植物学报,21(3):391-400. [Zhao L F,Li S,Pan Y,Yan G Q,Zhao G F. 2001. Population differentiation of Psathyrostachys huashanina along an altitudinal gradient detected by random amplified polymorphic DNA[J]. Acta Botanica Boreali-Occidentalia Sinica,21(3):391-400.]
Fan X M,Zhang Y D,Yao W H,Bi Y Q,Liu L,Chen H M,Kang M S. 2014. Reciprocal diallel crosses impact combining ability,variance estimation,and heterotic group classification[J]. Crop Science,54(1):89-97. doi:10.2135/cropsci2013.06.0393.
Mulcahy D L. 1979. The rise of the angiosperms:A genecological factor[J]. Science,206:20-23. doi:10.1126/science. 206.4414.20.
Nagamine T,Xiong J H,Xiao Q. 1992. Genetic variation in several isozymes of indigenous rice varieties in Yunnan Province of China[J]. Japanese Journal of Breeding,(42):507-513. doi:10.1270/jsbbs1951.42.507.
Rood S B,Buzzell R I,MacDonald M D. 1988. Influence of temperature on heterosis for maize seedling growth[J]. Crop Science,28(2):283-286. doi:10.2135/cropsci1988. 0011183X002800020021x.
Soehenei R,Srinives P. 2005. Significance of heterosis and heterobeltiosis in an F1 hybrid of mungbean (Vigna radiata (L.) Wilczek) for hybrid seed production[J]. Sabrar Journal of Breeding and Genetics,37(2):97-105.
Tanto T,Demissie A. 2000. A comparative genetic diversity study for four major crops managed under Ethiopian condition[J]. American Journal of Botany,87:783-792.
Wang S H,Tan Y L,Tan X L,Zhang Z L,Wen J C,Kou S Y. 2009. Segregation distortion detected in six rice F2 populations generated from reciprocal hybrids at three altitudes[J]. Genetics Research,91(5):345-353. doi:10.1017/S0016672309990176.
(責任编辑 陈 燕)
猜你喜欢
蔬菜(2021年7期)2021-11-27
四川农业大学学报(2017年3期)2017-11-10
山西农经(2016年5期)2016-02-28
中国畜牧兽医文摘(2015年3期)2015-01-25
种子科技(2014年7期)2014-05-18
种子科技(2014年8期)2014-05-18
湖北农业科学(2014年4期)2014-05-04
茶叶通讯(2014年2期)2014-02-27
种子科技(2014年9期)2014-01-22
生物学杂志(2013年1期)2013-04-11
