
咖啡果小蠹成虫触角感器的扫描电镜观察
2023-09-25 07:57刘卓凡孟洋李欣蓉田甜宋国敏孙世伟
热带农业科学 2023年8期
刘卓凡 孟洋 李欣蓉 田甜 宋国敏 孙世伟
(1.中国热带农业科学院香料饮料研究所 海南万宁 571533;2.云南农业大学植物保护学院 云南昆明 650201;3.华中农业大学植物科学技术学院 湖北武汉 430070;4.海南省热带香辛饮料作物遗传改良与品质调控重点实验室 海南万宁 571533;5.长江师范学院 重庆 408100)
咖啡果小蠹[Hypothenemus hampei(Ferrari)]属鞘翅目(Coleoptera)象甲科(Curculionidae)小蠹亚科(Scolytinae)咪小蠹属(Hypothenemus),主要分布在埃塞俄比亚、哥伦比亚、巴西、越南、泰国等70多个咖啡种植国家和地区,是一种危害咖啡种子的钻蛀性害虫,每年给全球咖啡产业造成约5亿美元的损失[1-2]。咖啡果小蠹于1985年在海口口岸首次被截获[3],于2019年第一次被发现在海南省万宁市入侵[4],并迅速扩散至海南省主要咖啡产区。咖啡果小蠹在海南多地的种群不断扩大,为害日趋严重,给当地咖啡产业造成了巨大的经济损失,对国内咖啡产业的健康发展构成严重威胁。该虫以雌成虫从果实脐区钻蛀进入内部,蛀食果肉和种子,并产卵于蛀道内。咖啡果小蠹雌雄比约10:1,繁殖能力强,每头雌虫可产卵20~60粒,待幼虫化蛹、羽化交配后,雌成虫飞离继续危害新鲜果实[5-6]。咖啡果小蠹是目前已知的唯一可在咖啡果实内完成完整生命周期的昆虫,其隐蔽的生活方式使得防治困难,化学防治具有局限性。因而,寻找一种绿色高效的防治手段对其进行控制十分关键。昆虫的感器是感知外界环境变化和对刺激产生反应的重要结构,大多富集于触角,其通过辨别不同气味使昆虫产生吸引和驱避反应[7-8]。植食性昆虫可以通过刺激触角上感器中的化学感器细胞,利用寄主植物挥发物来确定寄主、配偶和产卵、冬眠场所,不同类型的感器有不同的功能[9]。因此,研究咖啡果小蠹的触角感器类型对于了解其在寄主选择和定位过程具有十分重要的意义。
近年来,不少学者采用扫描电子显微镜技术研究了小蠹亚科昆虫的触角感器,包括华山松大小蠹(Dendroctonus armandi)[10]、红脂大小蠹(Dendroctonus valens)[11-12]、光滑足距小蠹(Xylosandrus germanus)[13-14]、脐腹小蠹(Scolytus schevyrewi)[15]、云南切梢小蠹(Tomicus yunnanensis)[16-17]、横坑切梢小蠹(Tomicusminor)[16]、短毛切梢小蠹(Tomicusbrevipilosus)[16]、小粒材小蠹(Xyleborus saxeseni)[18]、松六齿小蠹(Ips acuminatus)[19]、暗翅足距小蠹(Xylosandrus crassiusculus)[14]、咖啡黑(枝)小蠹(Xylosandrus compactus)[20]、南方松大小蠹(Dendroctonus vitei)[21]、落叶松八齿小蠹(Ips subelongatus)[22]、云杉八齿小蠹(Ips typographus)[23]等,这些昆虫的感器种类和功能有所差异,例如云南切梢小蠹、横坑切梢小蠹、短毛切梢小蠹和南方松大小蠹触角上的锥形感器表面具孔,被认为是一种嗅觉感器[16-17,21];红脂大小蠹和落叶松八齿小蠹触角上的毛形感器顶端具孔,兼具感受化学信息和机械刺激的功能[11-12,22]。咪小蠹属在国内少有分布,对属内特征的了解较为薄弱。前人对咖啡果小蠹的研究极少,主要集中在检疫方面,咖啡果小蠹作为新入侵害虫,目前尚无有关其触角感器的文献报道。本研究利用扫描电镜技术对咖啡果小蠹雌、雄成虫触角外部形态与感器进行观察,明确咖啡果小蠹成虫触角感器的超显微结构,并分析各感器的潜在功能,旨在进一步明确触角感器在寄主选择和定位过程中的作用,为后续开展咖啡果小蠹电生理学、行为学调控研究提供理论依据。
1 材料与方法
1.1 材料
咖啡果小蠹成虫采自位于海南省万宁市的中国热带农业科学院香料饮料研究所咖啡试验基地。
1.2 方法
1.2.1 材料准备在试验地内采集被咖啡果小蠹为害的咖啡果实,置于人工气候箱[T=(26.5±1)℃,RH=(75±5)%,14L:10D)]饲养,在体视镜下解剖果实,获取咖啡果小蠹成虫以备用。
1.2.2 试验设计取咖啡果小蠹雌、雄成虫各20头,用超声波清洗器(ELMA D-78224,德国)清洗1 min,室温干燥后放入4%戊二醛溶液,置于4 ℃冰箱中固定24 h;用0.2 mol/L磷酸缓冲液(pH 7.2)冲洗3次,每次1 min,分别用30%、50%、70%、80%、90%、100%乙醇逐级脱水,每次20 min;脱水后,将样品用乙酸异戊酯置换2次,每次30 min,取出后在CO2临界点干燥;将一小段导电胶粘在样品台上,在体视镜(OLYMPUS SZX16,日本)下,用镊子将干燥好的样品按照腹面、背面、外侧面和内侧面4个角度轻放在导电胶上;将样品台放入离子溅射仪(SBC-12,北京中科科仪股份有限公司)中镀金属膜;将经上述处理后的咖啡果小蠹置于扫描电子显微镜(ZEISS EIGMA,德国)下,观察其触角的形态特征、感器类型及分布特点,电子显微镜工作电压为10 kV。使用Adobe Photoshop CC 2015测量成虫触角各节长度及感器长度,3次重复,取平均值,对图片添加比例尺并进行标注。每类感器统计4个面的总数,3次重复,取平均值。
1.2.3 数据统计感器的类型主要参考Schneider[24]的方法进行命名和归类。记录触角各部位触角感器的数量及分布,触角感器百分比=(某种感器数量/感器的总数量)×100%。用Excel 2019记录获得的数据,通过SPSS 25.0软件进行计算与分析,并采用独立样本t检验方法(p<0.05)对雌、雄虫触角各节长度、感器长度和感器数量进行差异显著性分析。
2 结果与分析
2.1 雌、雄成虫触角的基本形态
咖啡果小蠹雌、雄成虫的触角(图1)结构无明显差异,均由柄节(Scape)、鞭节(Funicle)和锤头部(Club) 3部分组成。柄节1节,粗长,呈棒锤状,有少量感器,与头部连接处有一凹陷;鞭节共5节,第1节呈念珠状,第2、3、4、5节呈套筒状,且逐渐变粗,其中,第1节、第3节、第5节上均有少量感器,其余各节无感器;锤头部侧面扁平,正面椭圆形,感器排列成3条带状,带与带之间相互平行。触角各节表面上均有明显的瓦棱状花纹。雌、雄成虫触角及各节长度见表1,雌、雄成虫间触角长度、柄节长度和鞭节长度差异显著,锤头部长度二者之间不显著。

表1 咖啡果小蠹雌、雄成虫触角及各节长度 单位:μm

图1 咖啡果小蠹雌、雄成虫触角形态特征
2.2 触角感器类型
雌、雄成虫各类感器长度见表2,雌、雄成虫间锯齿形感器S.z.2与锥形感器S.b.1的长度存在差异,而其他类型感器长度雌、雄成虫间无显著差异。咖啡果小蠹雌、雄成虫触角上着生4类7种感器(图2、3),分别为:毛形感器(S.tr.1、S.tr.2)、锯齿形感器(S.z.1、S.z.2)、锥形感器((S.b.1、S.b.2)、芽形感器(S.bs.),其中毛形感器、锯齿形感器和锥形感器存在不同亚型。此外,在锤头部表面还观察到一种表皮孔。
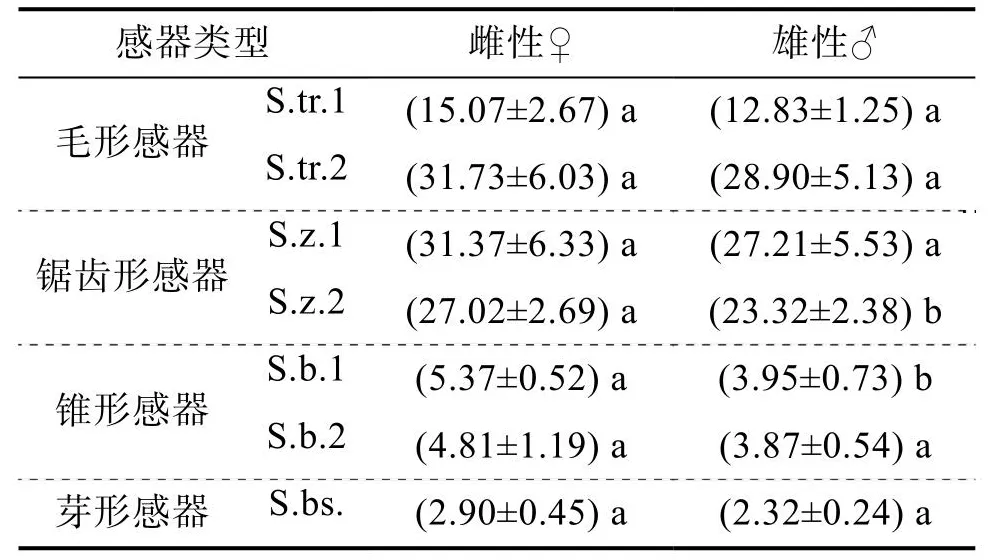
表2 咖啡果小蠹雌、雄成虫触角感器长度 单位:μm
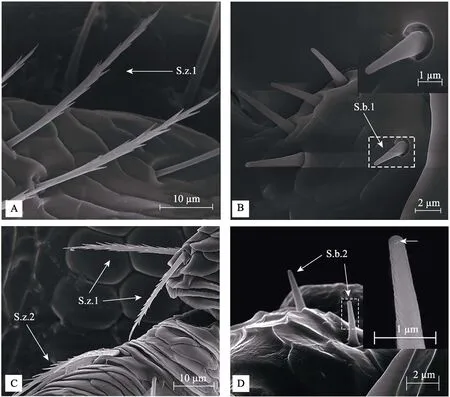
图2 咖啡果小蠹成虫触角柄节和鞭节上感器的形态特征

图3 咖啡果小蠹成虫触角锤头部上感器的形态特征
2.2.1 毛形感器(Sensilla trichodea, S.tr.)触角上数量最多的一类感器,约占总感器数量的52%。集中着生在锤头部,直立或微微弯曲。根据毛形感器的长度、弯曲程度、顶端情况分为2种亚型:第1种为毛形感器Ⅰ型(S.tr.1),大多数存在于锤头部的节间带上,无基底窝,弯曲程度不明显,逐渐变细至钝尖,感器表面密集分布凹陷小孔,数量最多,占毛形感器的90.3%;第2种为毛形感器Ⅱ型(S.tr.2),绝大多数散生在锤头部端部,毛发状,较S.tr.1长,数量少于S.tr.1,从底端向上逐渐变细,整体细长且顶端略尖,弯曲幅度略大,感器顶端带有小孔,占毛形感器的9.7%。
2.2.2 锯齿形感器(Sensilla zigzag, S.z.)直立或弯曲,在柄节、鞭节、锤头部均有分布,约占总感器数量的43.6%。根据感器锯齿排列方式和分布位置分为2种亚型:第1种为锯齿形感器Ⅰ型(S.z.1),存在于柄节和鞭节第1节,麦穗状,两侧有锯齿,占锯齿形感器的10.1%;第2种为锯齿形感器Ⅱ型(S.z.2),存在于鞭节第3节、第5节和锤头部,一侧光滑,一侧有锯齿,占锯齿形感器的89.9%。
2.2.3 锥形感器(Sensilla basiconica, S.b.)直立,形似短锥,数量稀少,占感器总数的3.9%。根据感器外壁和顶端是否具孔分为2种亚型:第1种为锥形感器Ⅰ型(S.b.1),存在于柄节与头部连接处以及锤头部,外壁光滑,顶端无孔,占锥形感器的53.5%;第2种为锥形感器ⅠⅠ型(S.b.2),存在于柄节和鞭节连接处以及锤头部,外壁凹凸不平,顶端有孔,占锥形感器的46.5%。
2.2.4 芽形感器(Sensilla bud-shaped, S.bs.)直立,形似禾谷类种子发芽。所有感器中形态最为短小并且数量最少的一种感器,着生于锤头部顶端,无基底窝,占感器总数的0.6%。
2.2.5 表皮孔(Cuticular pore, CP)分布在锤头部末端不着生感器的空白区域,多着生于感器附近,直径小于1 μm。
2.3 咖啡果小蠹成虫触角感器分布规律
咖啡果小蠹雌、雄成虫触角感器的分布见表3。咖啡果小蠹雌、雄成虫触角的感器类型以及分布特点无明显差异。各类感器的数量上,雌虫均高于雄虫,其中毛形感器S.tr.2的数量中,雌虫显著多于雄虫,其他类型的感器数量二者差异不显著。在所有触角感器中,毛形感器数量最多,约占总感器数量的52%,主要分布在锤头部的带状区域;锯齿形感器数量仅次于毛形感器,约占总感器数量的44%,除在第2~4鞭节未见分布,其余各节均有分布,这2类感器的数量占绝对优势。锥形感器、芽形感器数量合计占总数的4%左右,其中芽形感器仅分布在锤头部,其余感器在触角各部位均有分布。
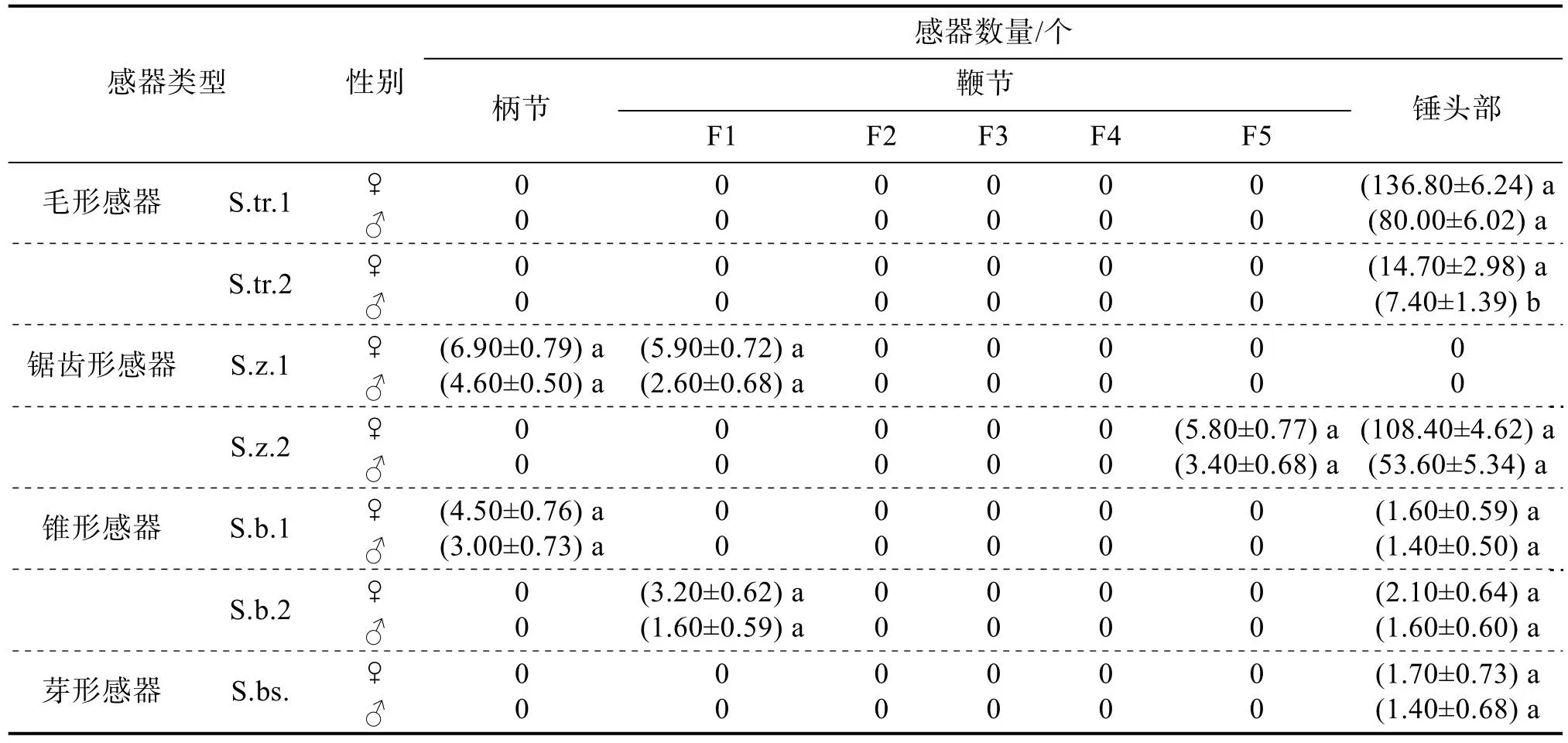
表3 咖啡果小蠹雌、雄成虫触角各节感器的分布及数量
咖啡果小蠹雌、雄成虫触角各节上的感器类型和数量有较大差异,柄节和鞭节感器类型相同,数量少且类型单一,均只有锯齿形感器和锥形感器,其中锯齿形感器都分布在柄节和鞭节表面,锥形感器都分布在连接处。柄节感器数量占总体的3.9%,鞭节感器数量占总体的5.1%。锤头部拥有全部类型的感器,且分布数量也最多,占总体的91.0%,锤头部感器分为3节,每节均由最外层的锯齿形感器S.z.2包裹着排列于内层的其他类型感器,第1节锯齿形感器环状排成1列,围绕节间轮生,毛形感器环状排成1列,主要分布在两侧,中间部分少有分布,其中一侧有1个锥形感器;第2节锯齿形感器和毛形感器均围绕节间轮生,其中锯齿形感器环状排成1列,毛形感器雌虫环状排成2列、雄虫环状排成1列;第3节各类感器密集散生,未有明显的环状排列。雌、雄成虫第2、4鞭节上均无任何感器。咖啡果小蠹与其他种类小蠹感器数量占比如图4所示。可以看出,咖啡果小蠹的锯齿形感器占比明显高于其他种类小蠹,但尚不明确咖啡果小蠹锯齿形感器数量显著高于其他种类小蠹虫的原因。
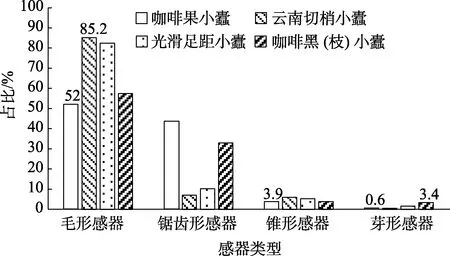
图4 咖啡果小蠹与其他种类小蠹各感器数量占比
3 讨论与结论
3.1 讨论
扫描电子显微镜对咖啡果小蠹雌、雄成虫触角感器的超显微结构观察。结果表明,咖啡果小蠹的触角上有4类7种感器。此外,在一些感器基部周围发现了一种表皮孔,这些表皮孔可能起到分泌润滑剂的作用[25]。
小蠹科成虫触角感器类型较为相似,咖啡果小蠹与足距小蠹属的光滑足距小蠹、咖啡黑(枝)小蠹触角感器类型及分布相似,都具有毛形感器、锯齿形感器、锥形感器和芽形感器,但在数量上有所差异,后2种小蠹虫的毛形感器在锤头部密集散生,而咖啡果小蠹则在节间轮生[13,20]。咖啡果小蠹的毛形感器与松六齿小蠹、华山松大小蠹和云南切梢小蠹分布相似,都分布在锤头部及带与带之间;咖啡果小蠹与松六齿小蠹、云南切梢小蠹的锯齿形感器分布相似,均在其触角的柄节、鞭节以及锤头部分布,但二者也存在一定的差异性,咖啡果小蠹的芽形感器散生于锤头部;华山松大小蠹芽形感器分布于柄节背面末端、鞭节和柄节的连接处或触角锤头部,云南切梢小蠹芽形感器着生于柄节基部的凹陷中[10,17,19]。
触角感器可以感受机械刺激和化学物质信息,是参与完成基本生物学行为的重要部分[26],不同类型的感器执行不同功能。毛形感器是最基础的嗅觉感器,兼具感受性信息素和寄主挥发物的作用,同时毛形感器可以帮助昆虫寻找合适的产卵场所[7,27-28]。不同亚型的毛形感器作用有所差别,咖啡果小蠹触角感器中毛形感器S.tr.1数量最多,这种感器存在于大多数小蠹虫的触角中,如红脂大小蠹、云南切梢小蠹、横坑切梢小蠹、短毛切梢小蠹、光滑足距小蠹、暗翅足距小蠹、落叶松八齿小蠹、云杉八齿小蠹,此类感器外壁上遍布许多细小的孔洞,这些孔洞可能与气味接收、识别和辨别有关[11,14,16,22-23]。因此,S.tr.1被认为是嗅觉感器的证据。毛形感器S.tr.2与Chen等[11]在红脂大小蠹和Shi等[22]在落叶松八齿小蠹发现的一种毛形感器相同,感器弯曲且顶端具孔,这种毛形感器已被发现具有化学感器和机械感器的功能。吴琼等[29]通过对华山松大小蠹触角进行透射电镜观察发现,锯齿形感器中无神经细胞和神经结构,只具有感器的外部形态特征,不具备感器的感受功能。咖啡果小蠹成虫触角上有着丰富的锯齿形感器,在锤头部锯齿形感器沿着毛形感器的密集带排列,为这些嗅觉感器提供了保护作用,并且可以防止虫体受到机械伤害。锥形感器内有丰富的神经细胞,具有识别气味的能力[12]。在其他小蠹上并未发现与咖啡果小蠹类似的顶端带孔的锥形感器S.b.2,光滑足距小蠹和暗翅足距小蠹的触角感器中存在一种顶端具孔的锥形感器,但其表面光滑,并且此感器数量雄性多于雌性[14]。S.b.2上的微孔可能与毛形感器S.tr.2顶端上的微孔作用类似,都可以通过感受外界气体物质形成神经冲动,咖啡果小蠹可能依靠触角上的锥形感器参与了寄主定位。芽形感器也被认为是一种锥形感器。Payne等[30]认为,芽形感器可能具有化学感受功能,但对化学物质的反应类型可能不同于其他锥形感器。感器内树突可沿毛发内腔向上延伸至尖端,并暴露于空气中以检测水蒸气、CO2或其他化学物质[31]。
雌性咖啡果小蠹触角整体长度、柄节长度和鞭节长度都显著高于雄性,而锤头部二者并未有差异,这可能是因为感器大多密集于锤头部,是感受化学物质的重要结构,因此二者差异并不显著。感器长度中,锯齿形感器S.z.2和锥形感器S.b.1雌性和雄性二者差异显著,这可能是由于雄虫一生只存活在果实内部,不存在雄性竞争和寻找食物等压力,因此,其保护型锯齿形感器长度显著小于雌虫;同时,由于生存压力小,雄虫识别气味的锥形感器长度显著小于雌虫。雌虫锤头部的毛形感器S.tr.2数量显著多于雄虫,这可能是因为S.tr.2参与了雌性寻找寄主植物和产卵场所过程。本研究观察了咖啡果小蠹雌、雄成虫触角的形态特征、感器类型和分布规律,明确了咖啡果小蠹雌、雄成虫触角感器的类型和分布规律及各种感器在寄主定位中的作用及功能,但需进一步通过透射电镜试验、电生理试验和行为学研究来证实。
3.2 结论
咖啡果小蠹触角具有4类7种感器,分别为:毛形感器、锯齿形感器、锥形感器和芽形感器,其中毛形感器、锯齿形感器和锥形感器均具有2种亚型。咖啡果小蠹的7种感器在雌、雄成虫上都有发现,不存在性二型现象,所有类型的感器数量雌虫都多于雄虫。雌、雄成虫触角感器的形态、种类和分布位置均无明显差异,以毛形感器数量最多,该类感器在咖啡果小蠹的寄主定位中起到重要作用,毛形感器S.tr.2数量雌雄间存在差异,与寻找寄主植物和产卵场所有关;锯齿形感受器数量与毛形感受器相当,起着感受机械刺激和保护嗅觉感受器的作用。雌雄成虫在触角长度、柄节长度和鞭节长度上有明显差异,锤头部长度无明显差异;雌雄成虫在锯齿形感受器S.z.2和锥形感受器S.b.1长度上有明显差异,可能与咖啡果小蠹的生存方式有关。
猜你喜欢
中国蜂业(2023年2期)2023-04-04
煤气与热力(2021年10期)2021-12-02
飞天(2021年6期)2021-06-28
华人时刊(2020年19期)2021-01-14
电子测试(2018年18期)2018-11-14
西南交通大学学报(2018年5期)2018-11-08
能源(2018年5期)2018-06-15
文学少年(有声彩绘)(2017年1期)2017-02-14
河北林业科技(2016年5期)2016-11-08
山地农业生物学报(2015年2期)2015-12-02
