
增温和氮添加对天山高寒草地生态系统多功能性的影响∗
2023-10-10 07:21谢晨笛任正炜董六文龙剑周小龙
新疆大学学报(自然科学版)(中英文) 2023年5期
谢晨笛,任正炜,董六文,龙剑,周小龙†
(1.新疆大学生态与环境学院,新疆 乌鲁木齐 830017;2.兰州大学生态学院,甘肃 兰州 730000)
0 引言
全球变暖和氮沉降所导致的生物多样性丧失与生态系统功能变化是21世纪人类面临的最大挑战[1].全球变暖与氮沉降会对植物群落物种组成产生复杂的影响,包括物种多度、物种分布变化和物种丧失[2].同时生态系统功能对两者的响应随着空间尺度的变化而呈现出不同的结果[3].这些复杂的变化过程有助于调控与缓解气候变化带来的负面效应,而阐明气候变化条件下生态系统功能和群落多样性的响应机制也成为当前生态学研究中的难题之一[4].
生物多样性是探究生态系统功能的基础[5],当前生态学家分别从基于物种和性状的角度对生物多样性的两个基本组成部分,即物种多样性和功能多样性开展研究[6].多样性稳定性假说认为物种多样性越高的群落其生态系统功能越稳定,越能抵御气候变化所造成的干扰[7-8],但是也有研究得出相反的结论[9].同时植物通过改变其功能性状如高度、比叶面积、种子大小及其传播方式等来适应外界环境条件的改变,并且这种改变最终会影响生态系统功能的变化[10-13].目前对植物功能性状与生态系统功能之间关系开展研究主要通过两种途径:一是基于功能性状的群落加权平均值(CWM traits)来反映群落水平的功能性状,一般采用物种的生物量或相对多度的加权平均来计算[14].二是基于功能多样性(Functional Diversity),即特定群落中植物功能性状值的大小、范围和分布状况[5,11].两种方法的关键之处都在于筛选出与生态系统功能密切相关的植物功能性状.相对物种多样性来说,植物功能多样性更多考虑了共存物种的互补和冗余,能明确反应群落物种间在资源互补利用程度上的差异[11].并且前期研究表明不同的功能性状可以对应于不同的生态系统功能,与生态系统功能变化具有密切关系[15],是生态系统过程的主要决定者,由此可以更为准确地预测生态系统的功能变化.但是迄今为止,相对基于物种的途径,基于功能性状途径所开展的研究依旧较少[16-18].
生态系统通过物质循环和能量流动将各个营养级上的生物与环境因子联系在一起,能为人类提供多种生态系统服务[19].众多的前期研究表明生物多样性与生态系统功能之间存在紧密联系,保护生物多样性对于生态系统功能的维持和适应气候变化具有重大意义[20].早期的研究重点集中于单一生态系统功能,如生产力、凋落物分解、土壤养分等与生物多样性之间的关系.但实际上生态系统可以同时提供多种生态系统功能,这些生态系统功能之间可能存在着促进或拮抗等复杂的相互作用关系,而只关注单一生态系统功能不能对生态系统进行全面的预测评价[21].因此,近年来生态学家们提出了生态系统多功能性(Ecosystem Multifunctionality,EMF)这一概念,并且有关生物多样性与生态系统多功能性之间关系的研究逐渐增多[22-23].从生态系统的角度来看,同时考虑多个生态系统功能能减少生物多样性的多功能冗余[24-25],生物多样性的重要性才得以体现[26].并且考虑生态系统多功能性更贴近不同群体对生态系统服务的需求[27-28].因此,对全球变化背景下生态系统多功能性与生物多样性之间的关系开展研究能帮助我们理解生态系统如何响应环境变化,具有重要的理论与现实意义.
草地生态系统是世界上面积最大的生态系统,为牧民提供了众多的生态系统服务[29].但长期以来,在草地开发利用中人们只关注于生产力和载畜量等单一的生态系统功能,而忽视了其它与养分循环有关的生态系统功能,导致草场退化、毒杂草增多等一系列的问题.因此需要整合多种生态系统功能对草地生态系统开展综合评价.高寒草地作为草地中最重要的类型,具有植物生活史短的特点,对全球变暖和氮沉降的响应强烈,是气候变化背景下的敏感区和脆弱区[30-31].我们在位于天山南麓的巴音布鲁克高寒草地开展增温与氮添加的野外控制实验,使用生态系统多功能性的概念对当地草地的响应模式进行评估,并基于物种和性状两种途径对其变化机制进行探究.本研究探索以下三个科学问题:1)在增温与氮添加条件下,天山高寒草地的物种多样性与功能多样性如何变化?2)单一生态系统功能与生态系统多功能性对增温与氮添加处理如何响应?3)在增温与氮添加条件下,生态系统多功能性与各类多样性指标之间的关系如何变化?
1 材料与方法
1.1 样地概况
巴音布鲁克草原位于天山南坡中段腹地,是中国最大的高山草原[32].本文的研究地点位于中国科学院巴音布鲁克草原生态系统研究站(42.53˚N,83.42˚E)附近.研究地点海拔约2 470 m,年降水量265.7 mm,年平均气温-4.8 ℃,年蒸发量1 022.9∼1 247.5 mm,年日照时数2 466∼2 616 h,年均积雪150∼180 d,无绝对无霜期.植被类型为高寒草原,优势种为羊茅(Festuca ovina)、草地早熟禾(Poa pratensis)和溚草(Koeleria cristata),土壤为栗钙土[33].
1.2 实验设计
为了模拟当地温度升高和氮沉降加剧的气候变化趋势,选择长势良好的典型高寒草原开展增温与氮添加实验.实验包括对照、增温和氮添加3个处理,每个处理重复6次,共有18块样区,采用随机区组设计.每块实验样区面积为5 m×5 m,平均分为两个半区,上半区用于植物和土壤的样品采集,下半区用于样方调查.每块样区之间用1 m的缓冲区隔离.氮添加选用尿素(CO(NH2)2),添加浓度为10 g/m2,增温采用实验开顶式增温罩(Open Top Chambers,OTCs)于2018年5月下旬一次性放置于样地中.增温罩由6片透明的有机玻璃组成,底部直径为1.5 m,顶部直径为1.2 m,垂直高度为0.5 m[34].为避免大型家畜放牧影响,实验样地使用围栏进行围封.但当地的小型野生哺乳动物(如旱獭和鼠兔等)可以出入.
1.3 土壤性质、植物性状和群落结构测量
2021年8月即实验开展第4年,使用样方法对不同样地中的群落结构进行调查,样方大小为0.5 m×0.5 m.将植物群落地上部分按照物种采集之后分别装入信封,70 ℃烘箱烘干至恒重,将各物种的生物量相加得到地上生物量.随后在每个样地中使用内径为3.4 cm的土钻分别钻取3个深度为20 cm的土样.通过筛子(2 mm)将植物根与土壤分离,土壤收集之后用于理化性质的测量.植物根系用水充分冲洗干净,然后在70 ℃下烘干,随后称重并计算转换为地下生物量.
对每个样地中的羊茅(Festuca ovina)、溚草(Koeleria cristata)、高山黄耆(Astragalus alpinus)、冰草(Agropyron cristatum)、细果薹草(Carex stenocarpa)、二裂委陵菜(Potentilla bifurca)、紫花针茅(Stipa purpurea)、莓叶委陵菜(Potentilla fragarioides)等常见种进行功能性状的测量,每个物种随机选取6株植株,将完整的植株剪下带回实验室测量高度.从每株植株中选取3片健康成熟的叶片,用扫描仪测量叶面积,然后放置于烘箱中,70 ℃下烘干,用电子天平测量干重,精确至0.000 1 g.同一时期在样地采集足量的常见物种的叶片,在70 ℃的室内烘箱中烘干,磨碎并过筛,测定叶片的碳、氮、磷含量.采用凯氏定氮法分析叶片氮和土壤有效氮,重铬酸钾加热法分析叶片碳和土壤有机质.土壤有效磷含量和叶片磷含量采用钼锑比色法测定.
1.4 数据分析
群落物种多样性采用物种丰富度、香农-维纳多样性指数、辛普森指数和均匀度指数来表征.群落功能多样性选取以下4个指标来表征:功能丰富度(FRic)、Rao的二次熵(RaoQ)、功能离散度(FDis)和功能均匀度(FEve).群落水平的功能性状采用均值加权性状值来表示,计算公式如下:
式中:pi为生物量的相对丰度;traiti为物种i性状值的平均值;s为群落物种丰富度.分别计算高度、比叶面积、叶片碳含量、叶片氮含量、叶片磷含量的群落均值加权值.
千年生态系统评估(Millennium Ecosystem Assessment,MEA)[35]将生态系统服务功能分为供给服务、文化服务、调节服务和支持服务.草原生态系统多功能性计算时功能指标选取多偏向于调节服务(气候调节、病虫害防治)和支持服务(土壤养分与组成、养分循环、初级生产力)[36]类指标.依照Farnsworth等[37]的观点,选择过程速率相对较快且能直接测量的功能指标进行生态系统多功能性计算,而部分生态系统固有的物理化学特性指标土壤pH、土壤含水率等,因其受非生物因素影响大且过程发生速率缓慢,不列入计算指标.综合考虑下,本次生态系统多功能性计算时,选取9个功能指标分别代表生态系统的气候调节能力(土壤有机质)、土壤特性与肥力(土壤有效磷、有效氮)、初级生产力(地上、地下生物量)、光合作用效率(群落透光率)和生态系统质量(群落水平叶片碳、氮、磷含量)[37-39]如表1所示,功能分类参考文献[36].

表1 用于生态系统多功能性计算的功能指标及其生态系统功能分类
采用平均值法和多阈值法分别计算生态系统多功能性.平均值法首先对生态系统参数指标进行标准化(Zscore标准化),采用单样本Kolmogorov-Smirnov检验是否服从正态分布,Z分数计算公式如下:
式中:Zij为样地i生态系统参数j的Z分数,i介于1∼n之间(n为样地数),j介于1∼9之间;Xij为样地i生态系统参数j的数值;λj为第j种生态系统参数在n个样地间的平均数;δj为第j种生态系统参数在n个样地间的标准差.
最后对每个样地不同生态系统参数指标的Z分数求平均值,即得到各样地的生态系统多功能指数,计算公式如下:
多阈值法评估多样性的增加分别达到多个阈值水平时的功能性变化,设置不同阈值(10%、30%、50%、70%、90%)并且分别计算生态系统多功能性,计算公式如下:
式中:F为总的功能数;fi表示某一生态系统内功能i的值,γi是将fi转化为正值的数学函数;ti表示阈值,计算中只需要根据需要更改阈值ti即可.
在获得上述指标后,首先使用单因素方差分析(ANOVA)和多重比较(HSD)来研究增温和氮添加对生态系统多功能性、物种多样性指数和功能多样性指数的影响.然后对生态系统多功能性与物种多样性指数和功能多样性指数进行线性回归分析,探究物种多样性指数和功能多样性指数与生态系统多功能性变化的关系,找到最能解释生态系统多功能性变化的多样性指标.
使用R语言的multifunc包[21]计算生态系统多功能性指数,vegan包[40]计算物种多样性,FD包[41]计算功能多样性,绘图使用ggplot2包[42].
2 实验结果
2.1 增温和氮添加对生态系统多功能性的影响
为期4年的增温降低生态系统多功能性而氮添加则增加生态系统多功能性(图1A),使用均值法与阈值法计算得到的生态系统多功能性具有同样的变化趋势.当阈值设置大于50%时(图1D∼F),增温有降低生态系统多功能性的趋势但是未达到显著(p>0.05),而氮添加则在阈值为90%时显著地增加生态系统多功能性(p<0.05).对单一生态系统功能的分析结果显示:增温显著降低地上生物量、对地下生物量没有影响,而氮添加则同时增加地上与地下生物量(图2A∼B),但是均未达到显著水平;此外氮添加增加土壤有机质、有效氮和有效磷含量(图2D∼F);增温降低叶片氮含量和磷含量(图2H∼I).

图1 增温(W)和氮添加(N)对生态系统多功能性的影响
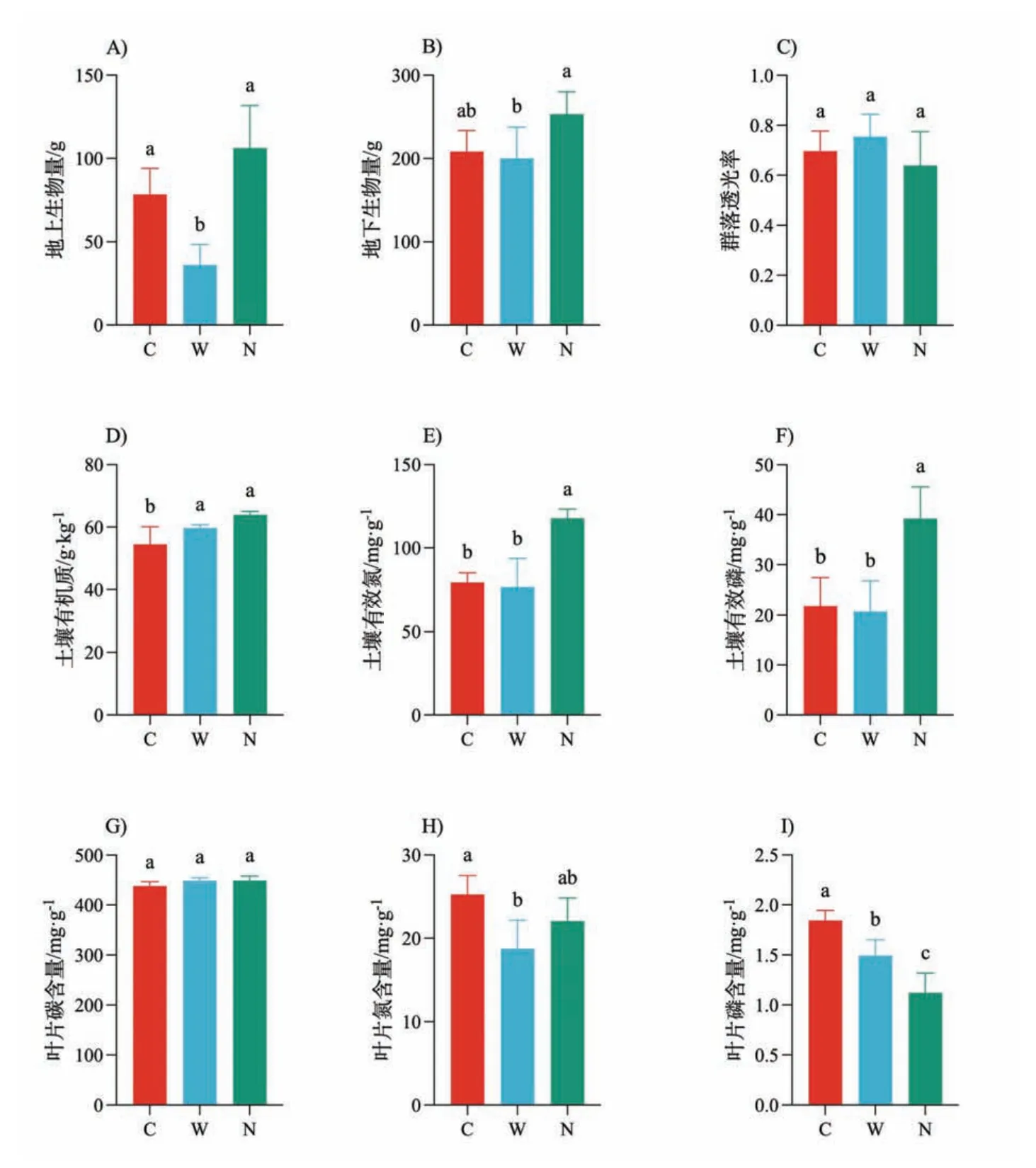
图2 增温(W)和氮添加(N)对单一生态系统功能(指标)的影响
2.2 物种多样性与功能多样性对增温与氮添加的响应
增温与氮添加对物种丰富度没有显著影响(图3A),同时增温增加香农-维纳多样性指数、辛普森指数和均匀度指数(图3B∼D);与此相反,氮添加则导致香农-维纳多样性指数、辛普森指数和均匀度指数显著降低(图3B∼D).功能多样性的变化较为复杂,氮添加对功能丰富度和功能均匀度的影响不显著,增温导致功能丰富度和功能均匀度增加,但是未达到显著水平(图4A,D).同时增温对Rao指数和功能离散度没有显著的影响,但是氮添加导致两者显著降低(图4B∼C).
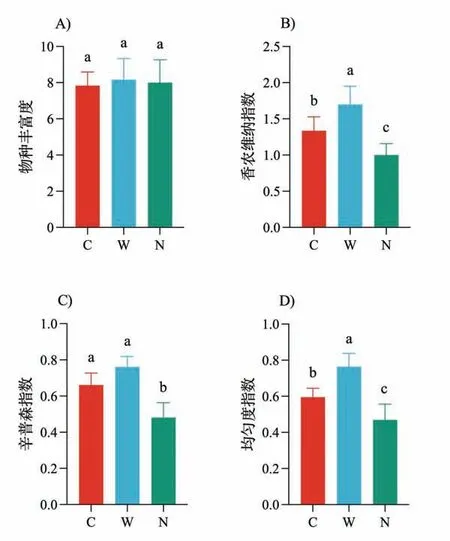
图3 增温(W)和氮添加(N)对物种多样性的影响

图4 增温(W)和氮添加(N)对功能多样性的影响
2.3 生态系统多功能性与多样性指标之间的关系
一元线性回归分析的结果表明:生态系统多功能性与香农-维纳多样性指数(R2=0.64,p <0.001)、辛普森指数(R2=0.54,p <0.001)和均匀度指数(R2=0.51,p <0.001)3个物种多样性指标均呈显著的负线性回归关系(图5B∼D);而对于4个功能多样性指标而言,生态系统多功能性与功能均匀度呈显著的负线性回归关系(R2=0.29,p<0.05),与功能丰富度、Rao指数和功能离散度均未出现显著的回归关系(图6).在所有多样性指标中,香农-维纳多样性指数解释能力最强.
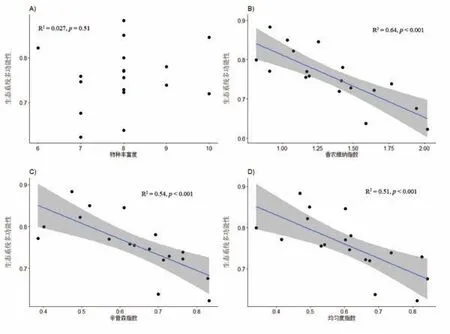
图5 物种多样性与生态系统多样性的关系
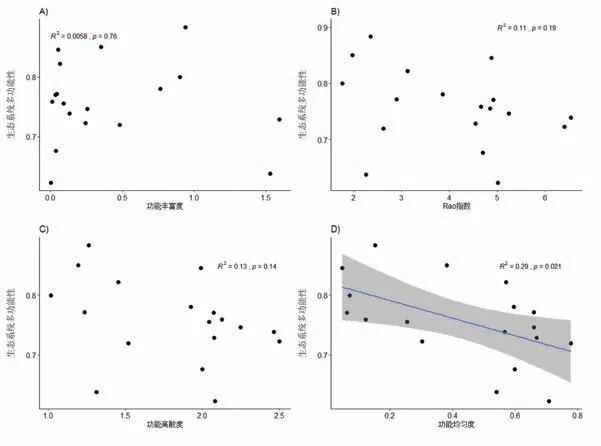
图6 功能多样性与生态系统多功能性的关系
3 讨论
3.1 增温与氮添加对天山高寒草地生态系统功能的影响
通过均值法和阈值法两种途径的计算结果均表明增温降低天山高寒草地的生态系统多功能性而氮添加则增加天山高寒草地的生态系统多功能性(图1).前期研究表明,温度变化对高寒草甸生态系统一系列生物过程影响很大[43],主要通过影响植物与微生物进而影响生态系统多功能性[44].而高寒草甸生态系统也会通过一系列生物化学过程缓解气候变化带来的影响,具体可以通过土壤有机质、植物盖度以及温室气体产出率等指标体现[37].对于高寒草甸生态系统而言,短期增温会增加生态系统多功能性,这可能是由于在较高温度下土壤微生物群落的生化反应速率和代谢需求增加[45],使得土壤功能发生变化.而长期增温,可能导致地上、地下生物多样性减少,从而对生态系统多功能性产生负面影响[46-47].我们的结果也支持该结论,在持续4年增温实验后,生态系统多功能性在增温条件下呈降低趋势.前期研究也表明,氮添加会改变土壤环境条件,导致植物群落组成和土壤微生物群落组成产生变化[48],从而对生态系统多功能性产生影响[49-50].我们的研究表明氮添加后生态系统多功能性增加,这与之前的研究一致[51].
从生态系统功能分类角度[36]进一步对单一生态系统功能的分析表明,增温导致生态系统初级生产力(地上、地下生物量)[46-47]和土壤特性与肥力(土壤有效氮、有效磷)下降,气候调节能力(土壤有机质)[52-56]和光合作用效率(群落透光率)增加,而选取代表生态系统质量功能(养分供给)的3个指标(群落水平叶片碳、氮、磷)变化不一致.光合作用效率提升并不能抵消其它生态系统功能降低造成的负面影响,最终增温使得生态系统多功能性降低.而氮添加虽然导致群落水平叶片氮、磷含量降低,但显著提高生态系统气候调节能力(土壤有机质)和土壤特性与肥力(土壤有效氮、有效磷),初级生产力(地上、地下生物量)也上升,这些生态系统功能的增加可以抵消群落水平叶片氮、磷含量下降所造成的影响,最终生态系统多功能性表现出增加的趋势.
与前期的研究一致[46-47],本研究表明增温降低天山高寒草地地上生物量,地上生物量的降低是由于增温导致群落中植株个体变小所致.增温导致群落水平叶片氮、磷含量均降低,这可能是由于增温阻碍植物氮、磷养分的吸收,影响生态系统养分供给速率.前期研究表明氮是草地生产力的重要限制因子,解除氮限制有助于提高草地生物量[57-58].我们在当地的研究也证明这一观点[59-60].本研究中氮添加有增加地上与地下生物量的趋势,但是未达到显著,这可能与我们的研究时间尚短(4年)有关.普遍的研究认为氮沉降会促进养分释放,增加微生物活性,加快土壤矿化过程,增加土壤养分含量[61],这与我们的研究结果一致.本研究中氮添加导致群落水平叶片磷含量下降,这可能是由于稀释效应,即氮添加导致植物叶片增大而单位面积上叶片磷含量下降.
3.2 增温和氮添加对物种多样性和功能多样性的影响
前期研究普遍证明增温与氮添加将导致物种组成和群落结构发生重大变化,但不同的生态系统类型会出现不同的响应模式[62].前期研究发现,对于亚洲内陆区域而言,春夏温度是影响植被生长的主要环境因素[63],而变暖会降低植物物种的多样性或丰富度[64].对于高寒草甸生态系统而言,植物群落多样性可能会随着温度的升高而降低[65],也可能对变暖没有显著的响应[66].对于高山生态系统而言,变暖对植物物种多样性既有积极影响,也有消极影响[67-68].本研究表明增温对天山高寒草地物种丰富度没有影响,但导致香农-维纳多样性指数、辛普森指数和均匀度指数显著升高.香农-维纳多样性指数和辛普森指数包含丰富度和均匀度两方面的信息[69],本研究中物种丰富度没有发生变化而均匀度显著增加,因此增温导致的香农-维纳多样性指数和辛普森指数的升高应该是由于增温处理中优势种的优势度下降、各物种分布更加均匀所致[70].
大量的研究已证明氮添加可以改变草地植物群落结构并降低物种多样性[71],导致多样性降低的机制包括土壤酸化、生态位维度降低[72]、资源竞争排斥假说[73]、营养失衡假说[74]、氮毒性[75-80].然而,也有研究表明存在其它限制因素(如磷、水分等)的情况下,氮添加可能对物种多样性没有影响[81].我们的研究表明,氮添加对物种丰富度没有影响,但是显著降低了香农-维纳多样性指数、辛普森指数和均匀度指数.与增温对香农-维纳多样性指数和辛普森指数影响机制相反,我们前期研究表明氮添加导致群落中禾草类物种(如羊茅、冰草、溚草和草地早熟禾)的优势度增加[59],进而导致均匀度指数降低,进一步导致了香农-维纳多样性指数与辛普森指数的降低.
对于功能多样性而言,我们的研究显示增温有导致功能丰富度和功能均匀度增加的趋势,但未达到显著水平,对Rao指数和功能离散度几乎没有影响,这可能与我们实验时间尚短有关,而氮添加则导致Rao指数和功能离散度显著降低,但对功能丰富度和功能均匀度几乎没有影响,这一结果与我们前期的研究结果一致[60].该结果说明为期4年的氮添加并未导致群落功能空间变化,且不同物种在功能空间内分布的均匀程度也没有显著变化.氮添加导致功能离散度降低,氮添加后群落优势种在功能空间中的分布呈现聚集趋势,这可能是由于氮添加群落中物种的功能性状都朝着相同方向变化,如增加植株高度获取更多光照以适应加剧的光竞争.
3.3 增温与氮添加条件下生态系统多样性与生物多样性之间的关系
前期的大部分研究均证明,物种多样性与生态系统多样性之间存现普遍的正相关关系[82],即植物物种多样性对生态系统多功能性存在限制作用[83],因为物种多样性高的群落能使更多的功能维持在高水平[82],极少研究显示物种多样性与生态系统多功能性之间存在负相关关系,而本研究中表征物种多样性的4个指标中有3个指标与生态系统多功能性呈负的线性回归关系,说明生态系统多功能性并非群落多样性所决定,即多样性高的群落未必生态系统多功能性也高.这可能是因为实验周期较短,部分生态过程速率较慢[36,84].也可能是因为功能性指标的选取未能很好地代表高寒草地实际的生态系统多功能性且未考虑多营养级多样性的作用,从而低估了群落多样性对生态系统多功能性的作用[82].如Díaz等[85]证明不管多样性的高低,生态系统的生产力和抗逆性不可能同时处于高水平,功能之间的权衡作用使任何一个群落不可能同时提供多种高水平的生态系统功能.本次研究只考虑植物物种多样性,可能是物种多样性导致光竞争加剧等原因影响植物对养分的吸收,导致生态系统生产力下降[86],而生态系统功能指标选择又未能很好地体现多功能间的互补性[24],导致群落多样性对生态系统多功能性的作用被低估.
表征功能多样性的4个指标中,生态系统多功能性仅与功能均匀度呈负的线性回归关系,这说明功能均匀度高的群落生态系统多功能性更低.一般而言功能均匀度高的群落可以对各种资源进行更加充分的利用,有助于提高生态系统多功能性,但本研究中氮添加后土壤有机质、有效氮和有效磷都增加(图2),因此地下养分已经不再是限制性因素,而对地上部分光资源的利用更加重要,因此植株高度更高的禾本科植物更加占据优势,这一过程导致群落的功能均匀度下降,但生态系统多功能性增加.在所有的指标中,香农-维纳多样性指数对生态系统多功能性的解释程度最高(R2=0.64)[84],说明生态系统多功能性受到植物多样性的制约[87].总的来说,群落多样性与群落功能多样性都能很好地解释生态系统多功能性的变化,说明保护生物多样性对于生态系统功能的维持有重要意义[82].
4 结论
通过在天山高寒草地进行为期4年的增温与氮添加实验,探究气候变化背景下天山高寒草地生态系统多功能性的变化模式及其与物种多样性和功能多样性之间的关系,结果表明:增温减小生态系统多功能性而氮添加则增加生态系统多功能性,增温导致的生态系统多功能性降低主要是由于初级生产力(地上生物量)和植物养分供给(群落水平叶片氮、磷含量)降低;而氮添加则通过显著增加初级生产力(地上生物量)、气候调节(土壤有机质)、土壤特性与肥力(土壤有效氮、有效磷)导致生态系统多功能性增加;物种多样性与功能多样性均可对生态系统多功能性变化进行解释,其中解释能力最强的指标为香农-维纳多样性指数.
猜你喜欢
作物研究(2023年2期)2023-05-28
昆明医科大学学报(2022年2期)2022-03-29
食品安全导刊(2021年20期)2021-08-30
今日农业(2020年19期)2020-12-14
河南畜牧兽医(2020年21期)2020-01-10
纺织科技进展(2015年1期)2015-11-28
中国科技信息(2015年2期)2015-11-16
水生生物学报(2015年1期)2015-02-28
植物营养与肥料学报(2014年1期)2014-03-11
中成药(2014年10期)2014-02-28
