
南五味子属叶绿体基因组密码子偏好性分析
2023-10-16 12:17尹登攀金吉芬彭志军李兴忠汪得凯
西北林学院学报 2023年5期
刘 涛,尹登攀,金吉芬,彭志军,李兴忠,汪得凯*
(1.贵州省果树科学研究所,贵州 贵阳 550027;2.浙江理工大学 生命科学与医药学院,浙江 杭州 310018;3.贵州省农作物品种资源研究所,贵州 贵阳 550027)
叶绿体在植物光合作用、胁迫响应及生长发育中发挥重要作用,叶绿体基因工程应用于提高作物产量、品质、抗性及生物固氮等多个方面[1-3]还用于生产疫苗等高价值的生物医学产品[4]。叶绿体高度保守,进化率较低,也是开发植物条形码的理想对象[5]。植物在长期的进化和自然选择压力下,由于对密码子使用频率的不同造成密码子的使用偏好性(codon usage bias,CUB),其偏好性受基因组组成、GC含量、基因长度和表达水平、基因中密码子的位置、重组率、mRNA折叠以及tRNA丰度等多种因素的影响[6]。密码子使用偏好性影响RNA加工、蛋白质翻译和折叠等多种过程,进而影响基因表达和细胞的功能[7]。因此,对密码子偏好性分析有助于理解生物的进化、系统发育关系、宿主-病原体共同进化关系和环境适应、个体基因的分子进化、物种之间的水平基因转移事件等[8]。此外通过筛选最优密码子,可以优化密码子而提高基因表达水平,进而提高叶绿体基因工程的效率[9]。
南五味子属(Kadsura)为五味子科(Schisandraceae)药用植物[10]。该属有10多种,如绯红南五味子(K.coccinea)常称“黑老虎”“冷饭团”等[11];异形南五味子(K.heteroclita)常称为“海风藤”“血通”等[12];凤庆南五味子(K.interior)称为“大鸡血藤”“顺宁鸡血藤”等[13];中泰南五味子(K.ananosma)被称为“消血藤”等[14]。南五味子属植物作为传统药用植物,具有活血化瘀、行气活血止痛、祛风散寒祛湿等功效[15]。从中分离到200多种化合物,主要成分为木脂素类、三萜类等[16-17]。现代药理学研究发现,南五味子属植物还具有抗炎、抗氧化、抗乙肝病毒及抗肿瘤等功能[18-20]。此外,果实富含20多种氨基酸和维生素C等营养成分,优于龙眼、荔枝等常见热带水果[21]。近年来,由于黑老虎丰富的营养和独特的风味而作为特色水果备受青睐,已成为脱贫致富的林下产业,在贵州东南、湖北恩施、湖南通道等地区广泛种植[22-23]。本研究对4种南五味子属植物叶绿体基因组密码子组成及密码子偏好性进行分析,为南五味子属植物的物种鉴定、种质资源保护以及性状改良提供参考。
1 材料与方法
1.1 材料
检索NCBI数据库,选取南五味子属中测序组装完整且注释良好的4个叶绿体基因组序列,分别为绯红南五味子(K.coccinea,NC_040116.1)、凤庆南五味子(K.interior,NC_050348)、中泰南五味子(K.ananosma,NC_057265.1)和异形南五味子(K.heteroclita,NC_057266.1),叶绿体基因组大小分别为158 573、153 201、145 903和153 289 bp。提取蛋白编码基因的CDS,选取以ATG为起始密码子且长度>300 bp的CDS序列,确定50条蛋白编码CDS用于深入分析。
1.2 方法
1.2.1 南五味子属密码子偏好性分析 叶绿体基因组密码子第1、2、3位及总GC含量采用CUSP (https://www.bioinformatics.nl/emboss-explorer/)分析,并计算同义密码子的相对使用频率(RSCU);有效密码子数(ENC)用在线工具chips(http://emboss.toulouse.inra.fr/cgi-bin/emboss/chips)计算;相关性分析利用SPSS 26软件进行;绘图采用R软件进行。
1.2.2 密码子中性绘图分析 分别统计密码子第1、2和3位碱基GC含量(GC1、GC2和GC3),GC1+GC2的平均值为GC12,以GC3为横坐标,以GC12为纵坐标绘制散点图并进行线性回归,计算GC3和GC12之间的相关系数。相关性绘图在集思慧远云平台(www.genepioneer.com)进行。
1.2.3 相对同义密码子使用度分析 相对同义密码子使用度采用Codon W1.4.2软件进行。RSCU值=密码子实际使用频率/无偏好性时理论使用频率表示。密码子无偏好性时,RSCU=1,密码子使用偏好性高于同义密码子时,RSCU>1。
1.2.4 ENC-plot绘图分析 有效密码子ENC计算公式
ENC=2+GC3+29/[GC32+(1-GC3)2]
(1)
然后以横坐标GC3,纵坐标ENC绘制散点图。
1.2.5 PR2-plot绘图分析 分别计算密码子第3位的4个碱基含量,以G3/(G3+C3)为横坐标和A3/(A3+T3)为纵坐标绘制散点图,并进行PR2偏倚分析。
1.2.6 最优密码子分析 以密码子ENC最低和最高10%的基因建立高低偏性库,计算2个库的RSCU和ΔRSCU,筛选标准为:高频率密码子,RSCU>1;高表达密码子,ΔRSCU>0.08;最优密码子,RSCU>1且ΔRSCU>0.08。
2 结果与分析
2.1 密码子偏好性分析
4个物种叶绿体蛋白质编码基因的GC含量(GCall)平均为40.20±0.04,均<50%, 4个物种间的GC含量分布变异范围很小,密码子3个位置的GC含量分布为:GC1>GC2>GC3,多以A/U结尾。ENC分布范围分别为:绯红南五味子,33.29 (rps14)~61.00(rpl22),平均值48.52;凤庆南五味子,33.03(rps14)~60.49(rpl22),平均值48.29;中泰南五味子,33.03(rps14)~61.00(rpl22),平均值48.27;异形南五味子,32.65(rps14)~60.47(rpl22),平均值48.40。4个物种rps14的ENC(33.00±0.26)<40,表现出较强的偏性(图1)。表明南五味子属4个物种叶绿体基因组密码子总体偏性较弱,物种间密码子偏好性较保守。
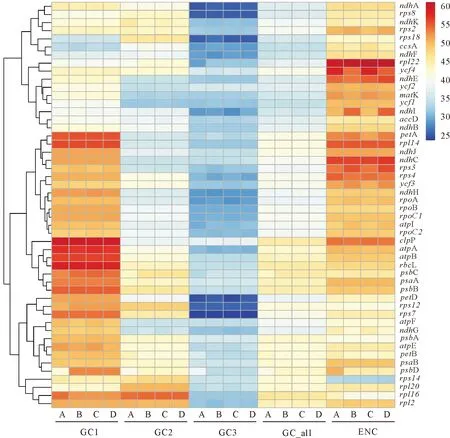
A、B、C、D分别代表绯红南五味子、凤庆南五味子、中泰南五味子和异形南五味子。图1 南五味子属叶绿体基因组密码子不同位置的GC含量和ENC值Fig.1 GC content and ENC value of three codons in the chloroplast genome of Kadsura
相关分析显示,4个物种的GCall与GC1、GC2和GC3都呈极显著正相关(P<0.01),而ENC与GC2和密码子数(Codon No)均呈显著负相关(P<0.01)(图2),提示有效密码子数受第2位密码子GC含量和密码子数影响较大,而受第1位和第3位密码子GC含量影响较小。进一步说明南五味子属4个物种的叶绿体基因组密码子参数高度保守。
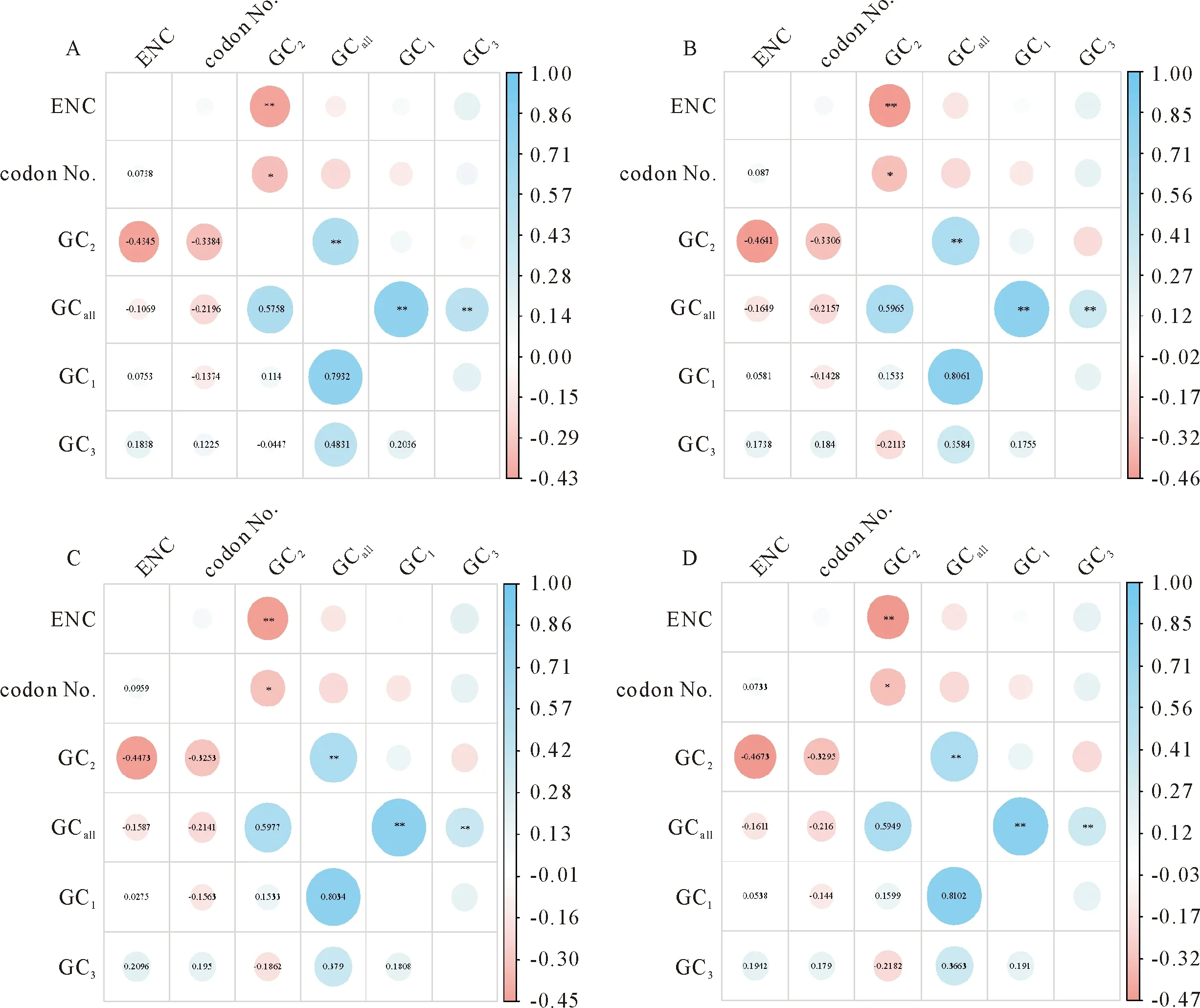
*表示P<0.05,**表示P<0.01。图2 南五味子属叶绿体基因组GC1、GC2、 GC3、GCall、ENC和密码子数量的相关性分析Fig.2 Correlation analysis of GC1,GC2,GC3,GCall,ENC,and codon numbers (cn) in the chloroplast genome of Kadsura
相对使用度(RSCU)分析结果发现,RSCU值在4个物种之间高度保守,有31个密码子的RSCU值≥1,以A和U结尾的密码子有28个,占比为90.3%,这些密码子在南五味子属物种中使用偏性较高,而RSCU值<1的密码子有30个,以A和U结尾的密码子有3个,占比9.7%,这些密码子是南五味子属叶绿体基因的非偏好密码子(图3)。
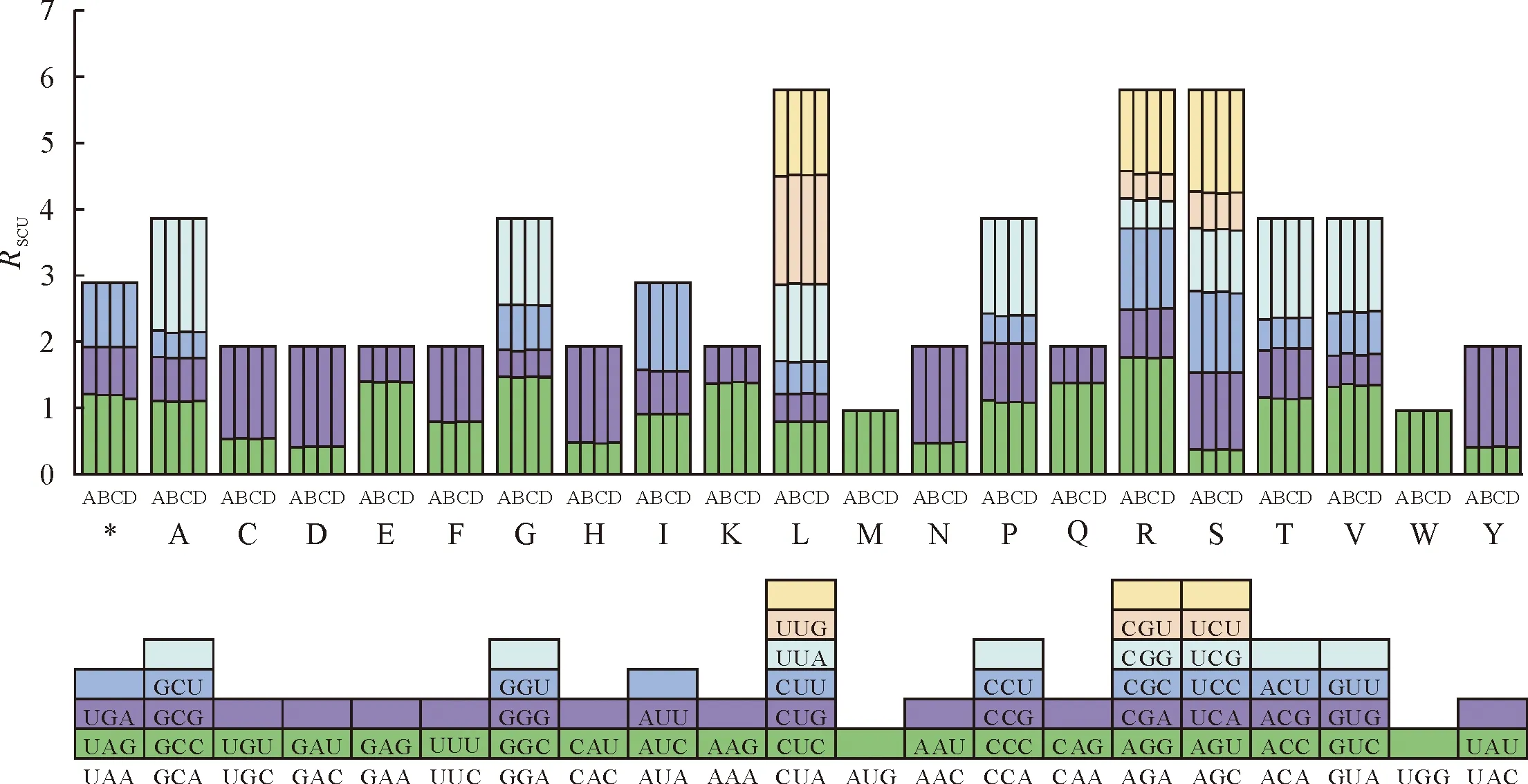
图3 南五味子属叶绿体基因组相对同义密码子使用度(RSCU)Fig.3 Relative synonymous codon usage (RSCU) analysis of genes in the chloroplast genome of Kadsura
2.2 中性绘图分析
中性绘图分析发现,4个物种横坐标GC3的分布范围分别为0.24 (rps7)~0.39(psbN)、0.22(rps11)~0.38(clpP)、0.22(rps11)~0.38(clpP)及0.22(rps11)~0.38(clpP),只有绯红南五味子叶绿体基因组密码子GC3极值的基因与其他3个物种有所差别。而纵坐标GC12的分布范围分别为0.36(ycf1)~0.54(rpl16)、0.36(ycf1)~0.56(rps11)、0.36(ycf1)~0.56(rps11)和0.36(ycf1)~0.56(rps11),其中绯红南五味子GC3最大值的基因与其他3个有所差别(图4)。4个物种的GC12与GC3的相关系数都基本接近为0,显示GC12与GC3之间的相关性不显著。而绯红南五味子、凤庆南五味子、中泰南五味子和异形南五味子4个物种的回归系数(曲线斜率)(即突变对偏好性的效应占14.91%)分别为0.149、0.008 62、0.016 5和0.001 67,进一步说明南五味子属叶绿体基因组密码子第1、2位和第3位碱基组成不同,基因组GC含量高度保守,除了绯红南五味子叶绿体基因组密码子的使用受突变影响较大(14.9%)外,其他3个物种受突变影响不超过2%,进而表明,南五味子属物种的密码子偏好性受自然选择影响为主,受突变的影响较弱。
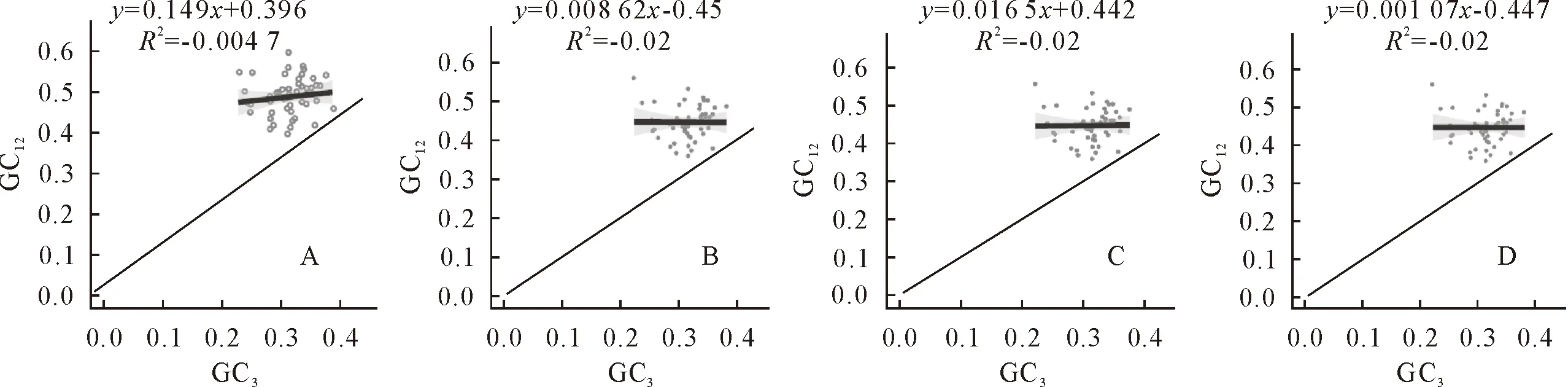
图4 南五味子属叶绿体基因组中性绘图分析Fig.4 Neutrality plot analysis of chloroplast genome in Kadsura
2.3 ENC-plot分析
ENC-plot分析结果显示,4个物种只有rps14的ENC低于40,离标准曲线最远,其偏好性更多地受到自然选择的影响,而突变对这些基因的影响较小(图5)。4个物种中,ENC值在-0.05~0.05的基因分别有12、15、12和13,分别占比24%、29%、24%和25%(表1),表明这些基因的ENC实际值和预期值与标准曲线接近,而其余基因的ENC实际值和预期值与标准曲线较远,表明自然选择是影响南五味子属叶绿体基因密码子的主要因素,与ENC-plot分析绘图分析结果一致。
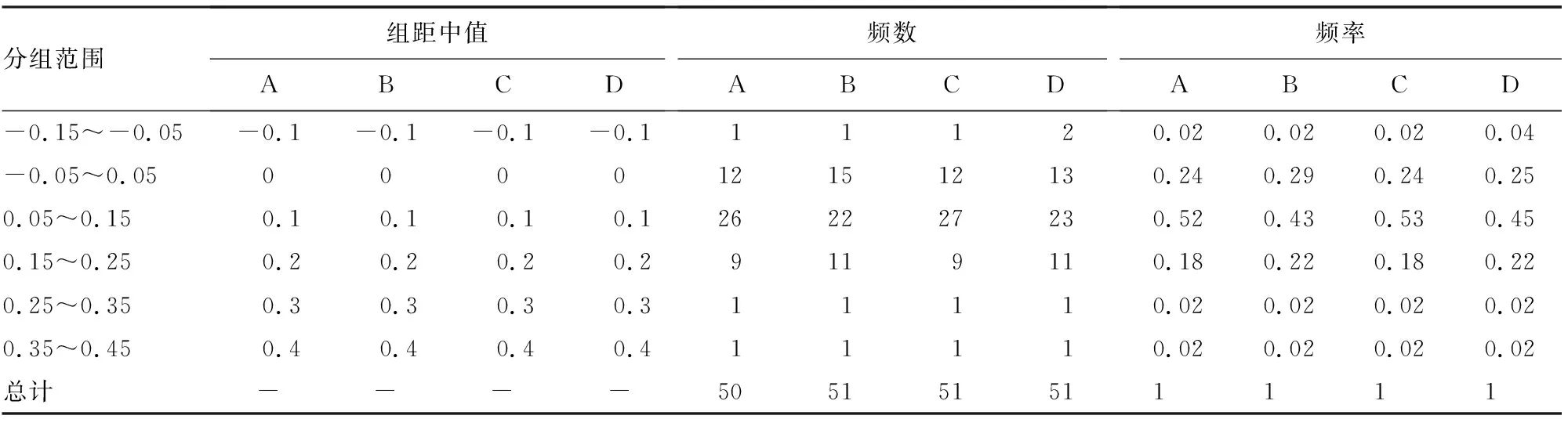
表1 南五味子属4个物种ENC比值分布Table 1 Distribution of ENC ratios in four species of Kadsura
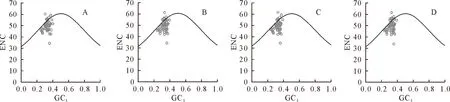
图5 南五味子属叶绿体基因组ENC-plot分析Fig.5 ENC-plot analysis of chloroplast genome of in Kadsura
2.4 PR2-plot分析
PR2-plot分析结果显示,坐标4个象限内均有基因分布,但4个区域中的基因位点分布不均匀,大多数基因分布在右下方(图6),表明4种植物中T/G密码子使用频率较高。综合分析显示GC3存在T/G偏性,进一步表明自然选择是造成南五味子属植物叶绿体基因组密码子使用偏好性的主要因素,而突变对密码子偏好性影响较弱。
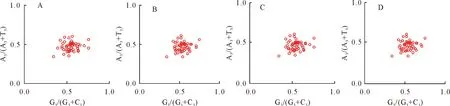
图6 南五味子属叶绿体基因组PR2-plot分析Fig.6 PR2-plot analysis of chloroplast genome in Kadsura
2.5 最优密码子分析
最优密码子筛选发现,在绯红南五味子、凤庆南五味子、中泰南五味子和异形南五味子4个物种中分别确定了18、17、16和18个最优密码子,其中以A/U结尾的密码子有15、12、11和13个,分别占比83.3%、70.6%、68.8%和72.2%,有14个密码子(ACC、ACU、CAU、CGU、CUA、GAU、GCU、GUA、UCU、UGU、UUC及UUG)为4个物种共同的最优密码子。ΔRSCU>0.5的高表达密码子分别有4、4、4和6个,其中共有最优密码子只有编码半胱氨酸(Cys)的UGU(表2)。表明4个物种中,使用频率最高的密码子和其编码的对应氨基酸各不相同。
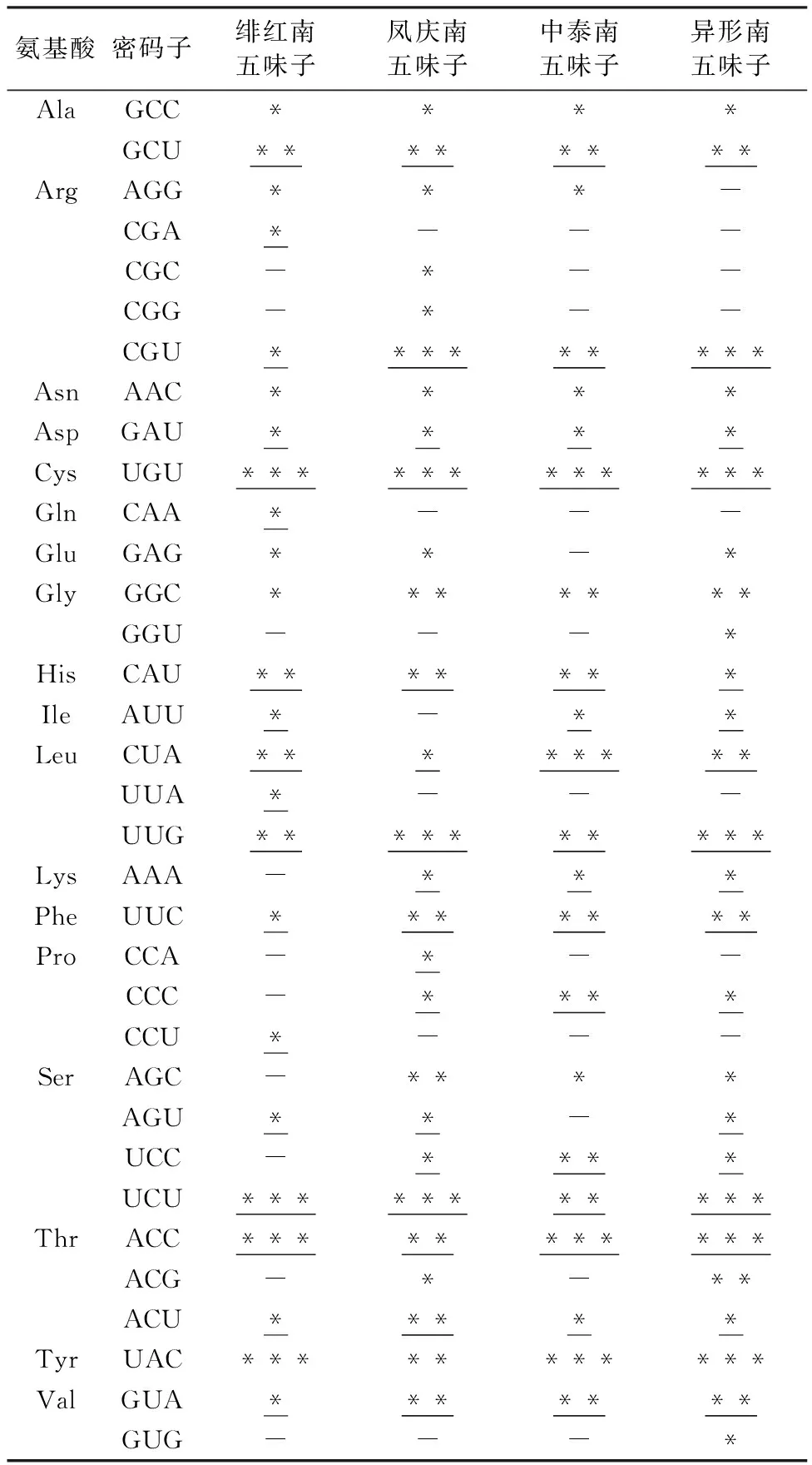
表2 南五味子属叶绿体基因最优密码子Table 2 Optimal codons of chloroplast genome in Kadsura
3 结论与讨论
3.1 结论
通过中性绘图、ENC-plot和PR2-plot 3种方法对密码子使用偏好性的影响因素进行分析,结果显示南五味子属4个物种叶绿体基因组密码子总体偏性较弱,物种间密码子偏好性较保守,自然选择是影响密码子偏好性的主要因素。绯红南五味子、凤庆南五味子、中泰南五味子和异形南五味子4个物种分别确定了18、17、16和18个最优密码子,共有的最优密码子为14个,ΔRSCU>0.5的高表达密码子分别为4、4、4和6个,其中共有最优密码子只有编码半胱氨酸(Cys)的UGU。研究结果为南五味子属植物系统发育和进化研究提供了新的见解,也为遗传育种研究提供了基础。
3.2 讨论
不同物种的氨基酸同义密码子的使用频率相差较大,同属植物、同一物种不同个体,甚至同一物种的不同基因对密码子的使用偏好性都存在差异[24-25]。叶绿体基因组GC含量一般<40%,本研究中,南五味子属4个物种叶绿体基因组密码子GC含量为40.20±0.04,与大多数绿色植物类似,4个南五味子属物种的叶绿体基因组密码子GC1、GC2、GC3和GCall平均含量均<50%,密码子均偏好使用A/U结尾,结果与茄属(Solanum)[26]、木兰科(Magnoliaceae)[27]、毛茛科(Ranunculaceae)和芍药科(Paeoniaceae)[28]、豆蔻属(Amomum)[29]、睡莲属(Nymphaea)[30]、榕属(Ficus)[31]等大多数植物一致。有效密码子数(ENC)是判断密码子偏好性强弱的主要指标,本研究南五味子属4个物种只有rps14的ENC(33.00±0.26)<40,其他49条序列ENC均>40,显示4个物种的叶绿体基因组密码子总体偏性较弱,结果与木兰科[27]、毛茛科和芍药科[28]、普通油茶(Camelliaoleifera)[32]、菠萝(Ananascomosus)[33]、樟叶槭(Acercinnamomifolium)[34]、小粒咖啡铁皮卡(Coffeaarabica‘Typica’)[35]等研究结果一致。在其他物种中,如菠萝(38.48)[33]、二乔玉兰(Yulaniasoulangeana)(35.94)和日本辛夷(Yulaniakobus)(35.94)[36]中也发现rsp14基因的ENC<40,提示rsp14基因的强密码子偏差在自然界中可能存在一定的普遍性。通常认为高表达基因的密码子偏好程度大,而其ENC较小,rsp14基因为管家基因,表达量较高。拟南芥中研究表明,叶绿体rsp14基因在热胁迫下起初表达量显著下调,随后恢复至正常水平,显示rsp14基因可能在植物适应环境温度方法发挥重要作用[37]。本研究中,南五味子属4个物种均为常绿植物,主要分布于长江以南省份,为应对不同季节的冷、热等环境胁迫,rsp14基因可能需要维持核糖体翻译机器的高效运转,因而受环境胁迫,受到自然选择影响出现了强密码子偏差。
叶绿体基因组密码子各参数之间的相关分析显示,4个物种GCall与GC1、GC2和GC3都呈极显著正相关(P<0.01),ENC与GC2和密码子数均呈显著负相关(P<0.01),4个物种的叶绿体基因组密码子参数高度保守。结果与木兰科[27]、毛茛科和芍药科[28]、豆蔻属[29]、睡莲属[30]、樟叶槭[34]、小粒咖啡铁皮卡[35]等植物类似。南五味子属为五味子科植物,与木兰科为亲缘关系较近,但其密码子GC含量及参数之间的相关性差异较明显,表明不同物种密码子各位置的GC含量受环境影响较大。
本研究显示,自然选择是影响密码子偏好性的主要因素,与毛茛科和芍药科[28]、樟叶槭[34]等植物类似。而木兰科[27]、豆蔻属[29]、睡莲属[30]、榕属[31]、小粒咖啡铁皮卡[35]等植物叶绿基因组密码子的使用不仅受自然选择影响,还受突变的影响。因此,不同植物,甚至同一科的植物,由于受到自然选择压力和突变压力不同,都会造成密码子偏好性差异。
通过RSCU分析和高频密码子筛选,绯红南五味子、凤庆南五味子、中泰南五味子和异形南五味子4个物种分别确定了18、17、16和18个最优密码子,共有的最优密码子有14个。其中ΔRSCU>0.5的高表达密码子分别有4、4、4和6个,其中共有最优密码子只有编码半胱氨酸(Cys)的UGU,尽管南五味子属4个物种叶绿体基因组GC含量及偏好性较保守,但在高频密码子及最优密码子的使用上差异较大。研究结果阐明了南五味子属4个物种的叶绿体基因组密码子的使用偏好性及其影响因素,同时也有助于南五味子属种质资源保护、系统发育研究、资源鉴定及叶绿体工程研究。
猜你喜欢
矿产勘查(2020年1期)2020-12-28
生物学通报(2020年11期)2020-10-22
保健医苑(2018年9期)2019-01-10
中成药(2018年7期)2018-08-04
唐山文学(2016年2期)2017-01-15
广西林业科学(2016年1期)2016-03-20
云南林业(2015年6期)2015-02-10
茶叶通讯(2014年4期)2014-02-27
食品科学(2013年6期)2013-03-11
中国烟草学报(2012年2期)2012-04-09
