
油松不同径向深度与方位树干液流特性研究
2023-10-16 12:23王梦蕾袁红娟孙照斌
西北林学院学报 2023年5期
王梦蕾,袁红娟,白 宇,孙照斌*
(1.河北农业大学 林学院,河北 保定 071000;2.河北省文安县自然资源和规划局,河北 文安 065800)
树干液流是植物进行蒸腾作用时,因蒸腾拉力所产生的树木水分流动过程[1-3]。树干液流的空间差异主要表现在径向深度、方位和垂直高度3个方面,目前研究主要集中在不同径向深度和不同方位,并发现大部分树种边材径向不同深度处和不同方位处的树干液流存在变化[4-5]。
由于林木间水分传输结构差异,距形成层不同深度的树干液流速率存在较大差异,如二白杨[6](Populusgansuensis)边材液流速率最大值在距形成层30 mm深处;尾巨桉(Eucalyptusurophylla×E.grandis)[7]、青海云杉(Piceacrassifolia)[8]树干液流速率在距形成层0~20 mm深处液流最大;四川山矾[9](Symplocossumuntia)在形成层10~30 mm处树干液流速率最大,杉木(Cunninghamialanceolata)[9]在距形成层30~50 mm处树干液流速率最大;黄土区刺槐(Robiniapseudoacacia)和辽东栎[10](Quercuswutaishanica)树干液流速率随径向深度增加而递减。
研究发现,不同方位间的树干液流速率也存在很大差异,如西伯利亚落叶松(Larixgmelinii)[11]和樟子松(Pinussylvestrisvar.mongolica)[12]液流速率在南侧较高,但枣树(Ziziphusjujuba)液流速率在西侧较高[13],胡杨(P.euphratica)的液流速率在东侧最高[14]。旱柳(Salixmatsudana)与小叶杨(P.simonii)的方位间差异随季节而变化等[15]。党宏忠等[16]研究发现,苹果(Maluspumila)不同方位液流速率大小排序为北侧>南侧>东侧>西侧;韩辉等[17]研究发现,沙地赤松(P.densiflora)的不同方位液流速率大小一般为南侧>西侧>北侧>东侧。可见,在树干液流速率测定过程中必须考虑到径向深度和方位差异,以免造成较大测算误差。
目前部分研究认为,树干液流的空间差异可能与树木边材水力结构的非均质性、树冠几何形状或根系分布的空间不对称性等有关[18]。对于影响树干液流的研究主要集中在外界因素(环境因子),其中温度、风速、湿度、光合有效辐射、水汽压亏缺等因素均会造成树干液流的变化[19-21],而树木本身的一些生物特征(木材解剖)对树干液流的影响研究并不多见[22]。
油松(P.tabuliformis)是中国北方地区的典型树种之一,广泛分布于东北、华北等地区,在中国北方森林群落中具有一定代表性。在外界环境大致相同的条件下,本研究选取河北省冀北山区大力推广的油松,通过测定不同径向深度和不同方位树干液流与木材解剖特征,分析树木构造对水分输送功能的影响,以期为不同树种树干液流的比较、耗水评价、林木水分传输机理提供理论依据。
1 材料与方法
1.1 试验地概况
试验地位于河北省承德市隆化县徐八屋林场(117°62′56″E,41°27′70″N),土壤类型为棕壤土,坡向为东坡,南坡度约13°。地势北高南低,海拔650~1 000 m。属暖温带半湿润大陆性季风型山地气候。全年日照时数为2 777 h,年平均温度5.1 ℃。日照时间随季节的变化与太阳辐射的变化规律一致,日照分布的季节以春季为最高,日照时间在1月和12月最少。全年降水量为480 mm左右,其中3-5月占全年降水量的14%,6-8月占70%左右,9-12月占12.5%,冬季占3.5%。年均有风日数110 d,最多130 d,风沙日数62 d,大风日数38~40 d,风向以西北风为主,春、秋两季多风,空气干燥,蒸发量比较大,多年相对湿度年平均为55%,年蒸发量1 400 mm左右。
该林场主要乔木有油松、落叶松、桦树(Betula)、山杨(P.davidiana)、椴树(Tiliatuan)、蒙古栎(Q.mongolica)、五角枫(Acermono)等。按照树冠圆满、树干通直、长势良好的原则,选择3株近熟油松个体作为研究对象,样树基本情况见表1。
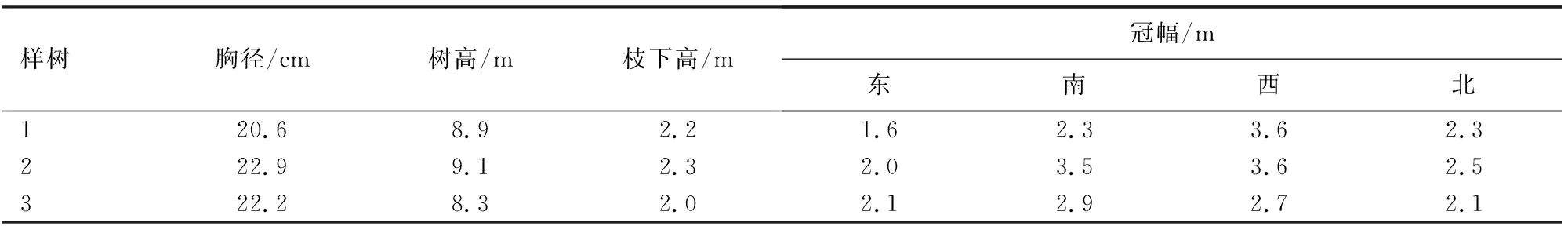
表1 样树基本情况Table 1 Basic information of sample trees
1.2 树干液流测定
测定方法按照马长明等[23]所采用的热扩散树干液流测定试验方法进行。2021年8-9月使用Granier热扩散式茎流计连续监测。树干液流速率按不同径向深度与不同方位分别测定。
测定不同径向深度液流速率时,选取探针直径2 mm,长度规格为10、20、30、50 mm,在样树胸高位置的北侧1/3圆周内,平行安装4组探针,探针长度从左至右分别为10、20、30 mm和50 mm,4组探针之间水平相隔约5 cm,保证探针之间互不干扰。
测定不同方位液流速度时,使用手持罗盘仪,确定东西南北4个方位,在4个方位上均插上直径2 mm、长度为30 mm的探针,保证探针之间互不干扰。
1.3 木材解剖特性测定
待树干液流测定完成后,将3株油松样树伐倒,在树干液流测定的1.3 m左右,截取3个厚度为50 mm圆盘(其中2个分别用于不同径向深度和不同方位解剖特性测定,另1个备用)。
测定不同径向深度解剖特性时,将1个圆盘通过髓心沿南北方向锯成与圆盘厚度相同的2 cm宽长条,在该长条上从形成层到髓心每隔10 mm取样,即将长条分为0~10、10~20、20~30、30~40、40~50 mm 5个部分,在各部分各取火柴杆粗细试样4个用于测定,代表各个径向深度的解剖特性。
测定不同方位解剖特性时,另取1个圆盘,在东、南、西、北4个方位,距形成层0~10、10~20、20~30 mm处各取火柴杆粗细试样2个,共6个试样,混合后用于测定,代表各个方位的解剖特性。
将上述各木材试样采用富兰克林离析法解离、制片[24],用3%番红水溶液染色1 min,将制好的片放置在数码显微图像电脑分析系统上对木材管胞形态测定。在10×物镜下观察木质部解剖特征,选取30个视野拍照。使用Image-J软件测量管胞宽度、管胞长度和管胞长宽比等数据后取平均值。
1.4 数据分析
运用Execl2013进行数据整理、计算,运用SPSS20.0进行曲线拟合及相关性分析,运用Origin2021做图。
2 结果与分析
2.1 不同径向深度树干液流速率
由图1可知,随着木质部径向深度的增加,油松的液流速率径向变化规律呈先降低后升高的趋势,其中距形成层30~50 mm处液流速率最大,液流速率峰值为0.009 7 g·cm-2·s-1;其次为在20~30 mm和0~10 mm处,液流峰值分别为0.008 4、0.005 0 g·cm-2·s-1;在距形成层10~20 mm处最小,峰值仅为0.002 5 g·cm-2·s-1。其中30~50 mm处液流速率分别为20~30、10~20、0~10 mm处液流速率的1.15、3.88、1.94倍。油松树干液流径向分布格局属于不规则型[25],液流密度由外向内的分布存在2个或者多个峰值,变化规律不明显。
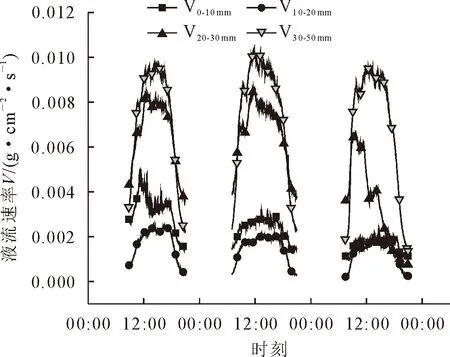
图1 不同径向深度树干液流速率Fig.1 Sap flow rate of trunkat different radial depths
根据油松不同径向深度液流变化规律,建立不同深度处液流速率的数量关系,将单一深度的树干液流速率转化为多深度处树干液流速率。由表2可知,油松V0~10 mm与V10~20 mm液流速率趋势拟合程度较高,线性拟合的决定系数(R2)为0.945;油松V0~10 mm、V10~20 mm与V30~50 mm的线性拟合直线斜率较大,分别为4.999和4.265,说明在0~10、10~20 mm与30~50 mm处液流速率差异较大,V0~10 mm与V10~20 mm的线性拟合直线斜率最小,V0~10 mm是V10~20 mm液流速率的1.153倍,说明在0~10 mm与10~20 mm处液流速率差异较小。

表2 不同径向深度树干液流速率间线性回归方程Table 2 Linear regression equation of sap flowat different radial depths
2.2 不同方位树干液流速率
由图2可知,树干液流速率值最高的来自于西侧(0.011 1 g·cm-2·s-1),其次为东侧(0.008 6 g·cm-2·s-1)和北侧(0.004 6 g·cm-2·s-1),南侧最低(0.003 0 g·cm-2·s-1)。东、南、西、北各个方位液流速率的峰值时间出现在12:00左右,12:00-16:00开始树干液流速率逐渐降低,16:00后树干液流速率急速下降。
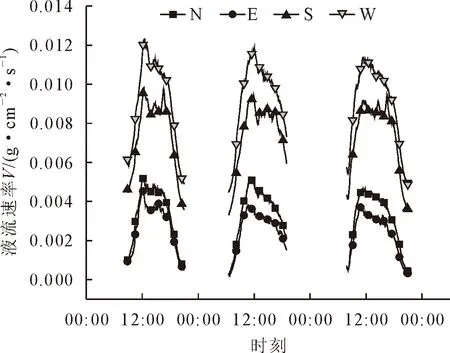
图2 不同方位树干液流速率Fig.2 Sap flow rates in different directions
为了明确不同方位间液流速率间的定量关系,以北侧(通常选择在树干北侧测量,减少阳光直射带来的测量误差)为因变量,建立其他方位与北侧液流速率间的回归关系(图3)。由图3可知,各方位液流速率与北侧液流速率均存在极显著的线性相关(P<0.01)。西侧与北侧间的线性拟合直线斜率为1.50,线性拟合R2为0.983;南侧与北侧间的线性拟合直线斜率为1.25,线性拟合R2为0.956;东侧与北侧间的线性拟合直线斜率为0.81、线性拟合R2为0.973。
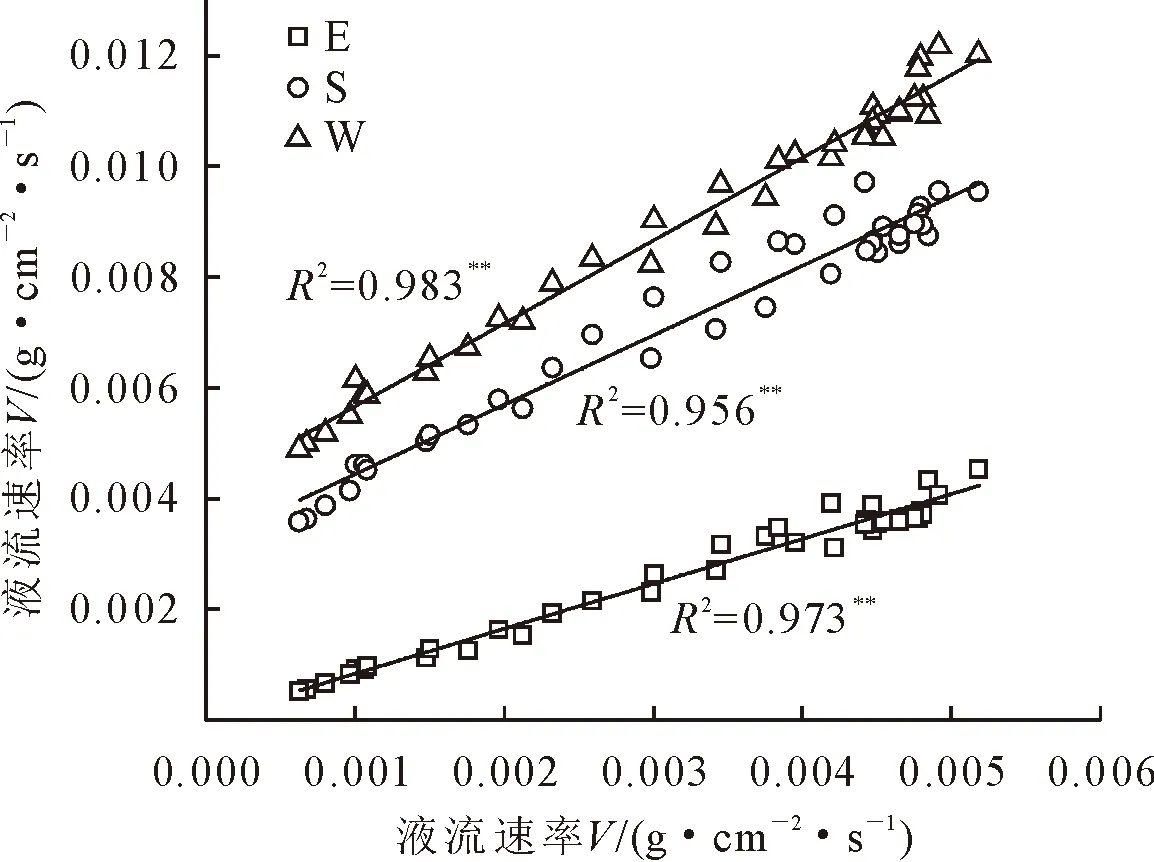
图3 北侧液流速率与其他3个方位液流速率间的相关性Fig.3 Correlationof the flow rate among the north side and the other three sides
2.3 树干液流对解剖特征的响应
树干液流特征反映了木质部不同深度和不同方位导水性的差异,而组成木质部管胞的结构特征(如管胞直径、长度和长宽比)是导致其水分运输能力差异的直接原因,进而反映了不同空间树干液流对木材构造的响应。
由图4可知,管胞长度和长宽比在距形成层40 mm处最大(3 196.256 μm),在20 mm处最小(2 644.93 μm)。管胞宽度在距形成层40 mm处最小(35.56 μm),在30 mm处最大(42.36 μm)。不同径向深度树干液流速率与管胞长度、管胞长宽比变化趋势一致,液流速率与管胞长度径向深度在10~20 mm处先降低,在20~50 mm处逐渐升高;管胞宽度与不同径向深度树干液流速率变化规律相反,在径向深度50 mm处树干液流速率最高,但管胞宽度最小。
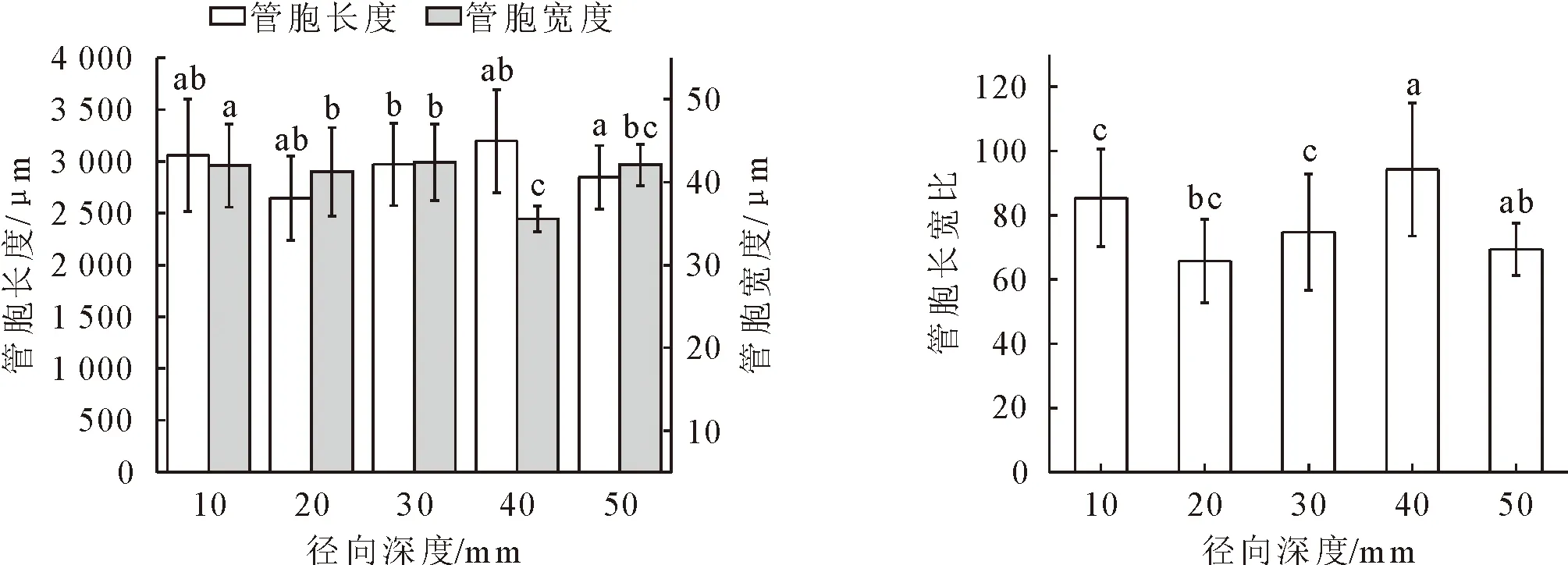
a、b、ab、bc表示不同方位管胞指标之间差异性。下同。图4 油松不同径向深度管胞解剖特征Fig.4 Tracheid length and tracheid width in different radial depths
由图5可知,管胞长度数值大小顺序为:西侧>南侧>北侧>东侧,西侧平均管胞长度(3 190.53 μm)分别为南侧(2 958.27 μm)、北侧(2 913.10 μm)、东侧(2 875.79 μm)的1.09、1.10、1.11倍。管胞宽度数值大小顺序为:东侧>北侧>南侧>西侧,东侧管胞宽度(42.00 μm)分别为北侧(40.56 μm)、南侧(37.09 μm)、西侧(32.09 μm)的1.04、1.13、1.31倍。管胞长宽比大小顺序为:西侧>南侧>北侧>东侧,不同方位管胞长宽比与管胞长度规律一致,西侧管胞长宽比(99.44)分别为南侧(79.78)、北侧(71.82)、东侧(68.47)的1.24、1.39、1.45倍。结合图1和图2的树干液流速率可知,树干液流变化特征与解剖构造之间响应程度较高。不同方位树干液流速率与管胞解剖构造变化趋势一致,西侧液流速率与管胞长度、管胞长宽比均高于其他3个方位,东侧液流速率与管胞长度、管胞长宽比均为最低值;管胞宽度与不同方位深度树干液流速率变化规律相反,管胞宽度越小,液流速率越大。
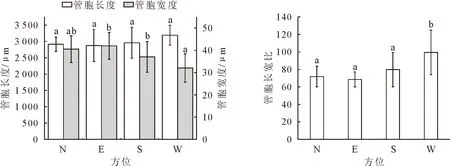
图5 油松不同方位管胞解剖特征Fig.5 Tracheid length and tracheid width in different orientations
3 结论与讨论
3.1 结论
随着木质部径向深度增加,径向液流速率变化规律为V30~50 mm>V20~30 mm>V0~10 mm>V10~20 mm,最大液流速率为0.009 7 g·cm-2·s-1,各深度液流速率之间呈显著线性相关关系;不同方位液流变化规律为西侧>东侧>北侧>南侧。树干北侧方位的液流速率与其他方位间均具有紧密的线性关系。
油松的管胞长度、宽度和管胞长宽比与树干液流的径向和方位变化规律趋势大致相同,即管胞长度越大,管胞宽度越小;管胞长宽比越大,树干液流速率越高;树干液流变化特征与解剖构造之间响应程度较高。
3.2 讨论
本研究中油松树干液流径向变化规律与张璇等[8]研究杉木在形成层30~50 mm处树干液流最大的结果一致。此外,孙守家等[26]、刘超等[5]、孙慧珍等[27]、王华等[28]、张小由等[29]大量学者都探索了树干液流密度沿径向分布的不均匀性,由于不同学者所研究的树木大小和树木种类存在很大差异,其心、边材的划分也不同,进而导致不同树种木质部径向分布规律不一致,所以液流速率最大值出现的位置也存在很大差异。由于本试验条件限制,最大径向深度仅做到50 mm处,并未进行更深层次树干液流的测定,有可能在距离形成层50 mm至心材部分液流速率还有减小,因此并未得出油松树干液流径向变化的完整变化规律,还有待于深入研究。
本研究中树干北侧方位的液流速率与其他方位间均具有紧密的线性关系,油松西侧液流量最大,与黄土区枣树[13]研究结果类似,但与其他学者对于西伯利亚松[11]、樟子松[12]等树种研究结果不一致。造成不同方位树干液流差异的原因可能与不同方位边材宽度、相应方位的冠幅大小、所处地理位置和太阳辐射等因素有关,本研究所测量树干液流的油松西侧冠幅较大,也可能是造成西侧液流量最大的原因之一。
木材中的水分输送通道是影响树干液流的自身因素,油松的主要水分运输通道是管胞,管胞的数量和直径都会对木质部水分运输产生一定的影响,随着管胞直径的增大,最大导水率和运输水分的效率也随之提高[29]。这是由于不同的木质部直径和长度影响木质部运输水分的阻力,木质部管胞的直径较小,长度较短,则会增加水分输送的阻力,使树木水分运输效率降低。这与本研究中树干液流速率与管胞宽度呈正相关关系结果一致。木质部解剖结构是栓塞抗性和水力导度的基础,与叶琳峰[30]研究的管胞与水力直径对树木输水效率的相关性较高,且管胞直径越大管胞密度越小的结果一致。
由于油松主要由管胞组成,结构较为简单,管胞的主要功能为运输水分和支持作用[32-33]。有学者认为对针叶树而言,具有输导作用的主要是轴向管胞,管胞直径对导水性的影响小于管胞长度对导水性的影响,因为管胞内水分运输的主要阻力来自于纹孔面上的纹孔,短的管胞比长的管胞阻力大[34],这与本文中管胞长度较管胞宽度与树干液流响应程度更高的结果一致。管胞长度和宽度主要是受内部因素(如植物激素等)和外部因素(如温度和降雨量)共同作用的结果[35],单位面积内管胞数量的多少也是影响树木水分运输的重要因素。油松为早晚材区别明显的树种,与早材相比,晚材管胞直径小,细胞壁较厚,管胞腔较窄,晚材水分流速较早材大。
树干液流的差异还可能受解剖构造的其他方面影响(如纹孔)。水分在木质部管孔中的传输阻力主要从管胞中产生的,其中纹孔口面积和纹孔膜面积越小,树干液流速率越大,树干液流率与纹孔结构之间存在着比较复杂的联系,纹孔面积假说中[36],当其上纹孔膜的总面积和数目增大时,在管孔内发生栓塞的可能性增大,本研究只对解剖构造进行了初步探索,还未涉及纹孔膜总面积、管胞上的纹孔密度、纹孔膜厚度等其余指标,尚不足以解释纹孔结构与树干液流速率的关系,尚需深入探究。
树种间树干液流径向分布格局的差异,不仅取决于木质部结构特征,还受气象因素、立地条件等外界的影响,未来对树干液流规律的研究需将内部与外部影响因素相结合,多角度、全面、综合地开展。
猜你喜欢
军事文摘(2021年16期)2021-11-05
山西林业(2021年2期)2021-07-21
现代园艺(2017年22期)2018-01-19
电源技术(2017年1期)2017-03-20
小猕猴学习画刊(2017年1期)2017-02-17
小猕猴学习画刊(2017年1期)2017-02-17
电源技术(2015年7期)2015-08-22
山西农经(2015年7期)2015-07-10
水土保持通报(2014年5期)2014-06-09
中国质量与标准导报(2014年5期)2014-02-28
