
外源茉莉酸对三七根系代谢物的影响及关键代谢物诱导三七抗病性分析
2023-11-02 16:01王正平罗丽芬张林段生双朱书生杨敏
南方农业学报 2023年6期
王正平 罗丽芬 张林 段生双 朱书生 杨敏




DOI:10.3969/j.issn.2095-1191.2023.06.009
摘要:【目的】揭示葉部喷施茉莉酸(Jasmonic acid,JA)诱导三七根系的代谢变化,明确显著变化的代谢物,为三七病害的生物防治提供理论基础。【方法】以三七为试验材料,在叶部接种黑斑病菌(Alternaria panax)菌株SL17,于接种4和8 d时测定三七植株内源JA含量;于三七叶片外源喷施不同浓度的JA后,利用气相色谱—质谱联用仪(GC-MS)分析三七根系代谢物变化,并检测显著变化代谢物对主要根腐病菌的抑制活性及其灌根后诱导三七叶部抗黑斑病的效果。【结果】叶部接种黑斑病菌4 d时,三七叶部JA和茉莉酸亮氨酸螯合物(JA-ile)的含量增加而在根系减少;接种8 d时,叶部JA含量显著增加(P<0.05,下同),而根系中2种物质的含量均极显著增加(P<0.01)。外源喷施不同浓度JA后三七根系代谢变化测定结果显示,三七根系中共检测出125种代谢物,主要分为有机酸类、氨基酸类、酯类、糖类、胺类、核苷酸及其衍生物和酮类等,其中有机酸类、氨基酸类和糖类是根系代谢物中的主要成分。与对照相比,叶部施用JA后三七根系中的草酸和氢化乳清酸含量显著增加。进一步测定显著变化的差异代谢物功能发现,草酸和氢化乳清酸对三七主要根腐病菌具有一定的抑制活性,其中氢化乳清酸在浓度为0.001 μg/mL时对茄腐镰刀菌(Fusarium solani)菌株F3和锈腐病菌(Ilyonectria destructans)菌株RS6的抑制效果最佳,浓度为0.010 μg/mL时对锈腐病菌菌株RS8的抑制效果最佳,浓度为0.100 μg/mL时对恶疫霉(Phytophthora cactorum)菌株D-6的抑制效果最佳;草酸浓度为0.001 μg/mL时对菌株F3的抑制效果最佳,浓度为10.000 μg/mL时对菌株D-6的抑制效果最佳;草酸和氢化乳清酸灌根后还能明显减小三七叶部黑斑病的病斑面积,且均在浓度10.000 μg/mL时诱导抗病效果最佳。【结论】外源喷施JA可改变三七根系代谢,促进氢化乳清酸和草酸的积累,进而增强三七对黑斑病的抗性,并对三七主要根腐病菌具有一定的抑制活性。
关键词:三七;黑斑病菌;茉莉酸;氢化乳清酸;草酸;抗病性
中图分类号:S435.675 文献标志码: A 文章编号:2095-1191(2023)06-1676-13
Effects of exogenous jasmonic acid on the metabolites of Panax
notoginseng (Burk.) F. H. Chen root system and the induction of disease resistance in Panax notoginseng (Burk.) F. H.
Chen by key metabolites
WANG Zheng-ping, LUO Li-fen, ZHANG Lin, DUAN Sheng-shuang, ZHU Shu-sheng, YANG Min*
(Yunnan Agricultural University/Key Laboratory of Agrobiodiversity and Pest Management, Ministry of Education/ State Key Laboratory for Conservation and Utilization Bio-resources in Yunnan, Kunming,Yunnan 650201,China)
Abstract: 【Objective】This study aimed to inresligate the metabolic changes of Panax notoginseng (Burk.) F. H. Chen root system induced by jasmonic acid (JA) spraying on leaves, and clarify the significantly changed metabolites, so as to provide a basis for biological control of P. notoginseng diseases. 【Method】P. notoginseng was used as experiment materials. This study determined the content of endogenous hormone JA in P. notoginseng after inoculation on leaves with Alternaria panax strain SL17 for 4 and 8 d. Metabolic changes in P. notoginseng root system were analyzed by using gas chromatography-mass spectrometry (GC-MS) after exogenously spraying on leaves with different concentrations of JA. This study measured the effects of significantly changed metabolites on resistance against major root rot pathogens and the induction of black spot disease resistance in P. notoginseng after root irrigation. 【Result】The content of JA and Jasmonic acid leucine chelate(JA-ile) in the leaves increased while the content of the both substances in the root system decreased after inoculation on leaves for 4 d. The content of JA in the leaves was significantly increased (P<0.05, the same below), and the contents of two substances in root system were extremely significantly increased (P<0.01) after inoculation for 8 d. After exogenously spraying different concentrations of JA, the metabolism change analysis of P. notoginseng root system showed that 125 metabolites were detected in the root system of P. notoginseng, mainly categorized as organic acids, amino acids, esters, sugars, amines, nucleotides and their derivatives and ketones, of which sugars, organic acids and amino acids were the main components. Compared with the blank control, the content of oxalic acid and dihydroorotic acid in the root system of P. notoginseng increased significantly after exogenously spraying JA on the leaves. Further study on significantly changed differential matebolities revealed that oxalic acid and dihydroorotic acid had certain inhibitory effect against the major root rot pathogens of P. notoginseng. Among them, dihydroorotic acid with the concentration of 0.001 μg/mL had the best inhibitory effect against Fusarium solani strain F3 and Ilyonectria destructans strain RS6. Dihydroorotic acid with the concentration of 0.010 μg/mL had the best inhibitory effect against I. destructans strain RS8. Dihydroorotic acid with the concentration of 0.100 μg/mL had the best inhibitory effect against Phytophthora cactorum strain D-6. Oxalic acid with the concentration of 0.001 μg/mL had the best inhibitory effect against F. solani strain F3. Oxalic acid with the concentration of 10.000 μg/mL had the best inhibitory effect against P. cactorum strain D-6. Oxalic acid and dihydroo-rotic acid significantly reduced the lesion area of black spot disease on leaves of P. notoginseng after root irrigation and represented the most induction of disease resistance in the concentration of 10.000 μg/mL. 【Conclusion】Exogenously spraying JA can alter P. notoginseng root system metabolism and promote the accumulation of dihydroorotic acid and oxalic acid, which enhance plant resistance against black spot disease and show inhibitory effect against main root rot pathogens.
Key words: Panax notoginseng (Burk.) F. H. Chen; Alternaria panax; jasmonic acid; dihydroorotic acid; oxalic acid; disease resistance
Foundation items: National Natural Science Foundation of China (32060719); Yunnan Young and Middle-aged Reserve Talent Project (202005AC160045); Yunnan Innovation Team Project (202105AE160016); Yunnan Agriculture and Forestry Joint Key Project (202101BD070001-003)
0 引言
【研究意义】三七[Panax notoginseng (Burk.) F. H. Chen]是我国独具特色的重要药用资源,为五加科(Araliaceae)人参属(Panax)植物。三七主要应用于心脑血管系统疾病的治疗(Watanabe and Tatsuno,2021),具有活血抗栓、止血、补血的药理作用以及调节血压的功效(王小其,2020)。三七主要分布于我国西南部1200~2200 m的高海拔地区(邹石龙,2018),其独特的生长环境易诱发多种病害,且随着三七在各地种植年限的增加,病害的种类、发病面积及严重程度逐年上升,严重影响三七的产量和品质(罗群等,2010;张帅等,2020)。根腐病和黑斑病是目前三七生产中危害较大的病害(蒋妮等,2011),严重制约了三七产业的健康发展(孙萌等,2015)。有研究显示,生产中三七病害的防控措施主要包括农药施用、开垦新地和轮作(张金丽,2022;Wang et al.,2022)等,但这些措施均不能完全减轻病害的发生,农药过量施用还会带来严重的农药残留和重金属含量超标等问题。因此,亟需研究绿色生态的防控措施以减轻三七主要病害的发生危害,对保障三七产业的健康发展具有重要意义。【前人研究进展】茉莉酸(Jasmonic acid,JA)作为一种信号物质可参与植物应对生物和非生物胁迫的防御反應(严加坤等,2019;张明菊等,2021)。研究发现,外源JA处理不仅能减轻水稻稻瘟病和葡萄灰霉病的发病程度(王云锋等,2018;郑素慧等,2022),参与西瓜对根结线虫的系统防御诱导(Yang et al.,2018),还能有效诱导辣椒对青枯病(赵显阳等,2018)、草地早熟禾对白粉病的抗性(赵泽花等,2020)。对拟南芥、大麦、小麦、玉米和珍珠粟进行的几项研究表明,JA调控着植物对干旱胁迫的耐受性(Awan et al.,2020;Wang et al.,2021)。魏婷等(2021)通过外源施用施茉莉酸甲酯(MeJA)处理番茄幼苗提高了其对镉(Cd)的抗性。李颜等(2022)研究发现,外源添加JA不仅可以缓解砷(As)在水稻幼苗根系中的积累,还能提高水稻幼苗的抗性,减少As对水稻幼苗的损害。Han等(2022)研究发现,JA可提高茶树的耐冷性。因此,JA在植物遭受病原菌胁迫时也起着重要作用。另外,JA还可调节植物基因的表达,进而积累次生代谢物,从而增强植物对各种胁迫的抵抗力(Marquis et al.,2022)。Yang等(2019)研究发现,JA和水杨酸(SA)的平衡能促进碳和氮资源的再分配,通过增加次级代谢产物有效增强圆锥铁线莲对紫外光的氧化防御。【本研究切入点】已有研究表明,外源施用JA一方面能使植物根系代谢发生变化,还能提高植株本身的抗性,但三七叶片施用JA后根系代谢是否发生改变,是否影响植株的抗病性等方面的研究目前尚未见报道。【拟解决的关键问题】以三七为试验材料,测定叶部接种黑斑病菌后三七叶片和根系中JA含量的变化,进而利用气相色谱—质谱联用仪(GC-MS)分析外源喷施JA后三七根系代谢物变化,明确显著变化代谢物对主要根腐病菌的抑制活性及其灌根后诱导三七叶部抗黑斑病的效果,为深入解析JA诱导植物根系代谢变化机制及三七黑斑病生物防治提供理论基础。
1 材料与方法
1. 1 试验材料
1. 1. 1 供试材料 健康的一年生三七苗来源于云南省昆明市寻甸县云南农业大学现代农业教育科研基地(东经103°16'41.6",北纬25°31'8.0",海拔1980 m)。供试黑斑病菌(Alternaria panax)菌株SL17、茄腐镰刀菌(Fusarium solani)菌株F3、恶疫霉(Phytophthora cactorum)菌株D-6、锈腐病菌(Ilyonectria destructans)菌株RS6和RS8均由云南农业大学农业生物多样性与病虫害控制教育部重点实验室自三七病株分离培养获得,并经过致病性测定。
1. 1. 2 供试试剂 试验所用化合物JA、氢化乳清酸和草酸,纯度均为98%,购自上海源叶有限公司。
1. 2 试验方法
1. 2. 1 接种黑斑病菌不同时间点对三七植株JA含量的影响 用接种针挑取培养7 d的黑斑病菌菌饼(直径为6 mm)接种于三七叶片伤口处,每株三七接种2片叶片,接种4和8 d后分别收集未接种黑斑病菌的所有剩余叶片和根系,经液氮研磨后,称量150.0 mg于1.5 mL离心管中,委托中国科学院昆明植物研究所进行测定。本研究使用岛津超高效液相色谱仪LC-20A和三重四极杆质谱仪LCMS-8040联用系统,每个样品吸取10 μL注射到ODS柱(1.6 μm,75 mm×2 mm)(Shim-pack XR-ODS III 2.0)中,柱温40 ℃。以0.3 mL/min的流速,由溶剂A(0.05%甲酸,5 mmol甲酸铵)和溶剂B(甲醇)组成的流动相以梯度模式进行洗脱,洗脱程序:1~3 min,B=20%;3~6.5 min,B等速升级到95%,维持0.5 min后,7 min恢复20%。采用电喷雾离子源负离子模式进行检测,离子源接口电压4.5 kV,离子源温度80 ℃;扫描方式为多反应监测(MRM)。
1. 2. 2 外源施用JA对三七植株根系代谢的影响
1. 2. 2. 1 样品前处理 分别配制0.001和0.010 mmol/L的JA(分别记为JA_0.001处理和JA_0.010处理)备用。选择18盆长势一致的一年生健康三七苗,每处理6盆,将配制好的JA溶液喷施三七苗叶部,以喷施1%甲醇为对照,24 h后取样。将上述处理三七根系经液氮研磨后取50.0 mg于1.5 mL离心管中,加入1 mL萃取液(甲醇∶氯仿∶水=5∶2∶2),涡旋提取6 min,4 ℃ 14000 r/min离心3 min,取450 μL上清液备用。将上述所有样品放入真空干燥器中干燥。
1. 2. 2. 2 衍生化 在干燥好的样品中加入80 μL盐酸甲氧胺吡啶(20 mg/mL),80 ℃恒温处理60 min,冷却至室温后加入40 μL N-甲基-N-(三甲基硅烷)三氟乙酰胺(MSTFA),75 ℃处理90 min;4 ℃下14000 r/min离心3 min,取上清液80 μL,装入有内补管的GC进样瓶,上机备用。
1. 2. 2. 3 GC-MS 檢测仪器为气相色谱—质谱联用仪(GC-MS-QP2010 ultra,Shimadzu,Japan),GC条件:SH-Rxi-5Sil MS色谱柱(30.00 m×0.25 mm×0.25 μm)。起始柱温40 ℃,以3 ℃/min升温至80 ℃后以5 ℃/min升温至260 ℃,保持30 min。载气为氦气,进样口温度250 ℃,进样量2 μL,进样方式为分流进样,分流比为10∶1。MS条件:EI电离源,离子源温度230 ℃,接口温度250 ℃,扫描范围35~500 m/z,采集方式scan,扫描间隔0.3 s。采用岛津脱机软件、Analysis Base File Converter软件、MSDIAL ver 3.52软件和NIST 14数据库进行峰值的识别和相关数据生成。数据归一化及后续分析在Metaboanalyst 4.0上完成。
1. 2. 3 差异代谢物氢化乳清酸和草酸对三七根腐病病原菌菌丝生长的影响 参考罗丽芬(2019)的方法进行。分别将利用二甲基甲酰胺(DMF)溶解后的氢化乳清酸和利用无菌水溶解后的草酸添加到已灭菌的PDA培养基中制成终浓度为0.001、0.010、0.100、1.000和10.000 μg/mL的平板待用,分别以含有1% DMF和无菌水的PDA培养基为对照。在培养基中央分别接种直径为6 mm的菌株F3、D-6、RS6和RS8菌饼,25 ℃暗培养,待对照菌落长至培养皿的2/3时采用十字交叉法测量各处理菌落直径,每处理6次重复。
1. 2. 4 差异代谢物氢化乳清酸和草酸对三七抗黑斑病的诱导效果测定 分别配制0.001、0.010、0.100、1.000和10.000 μg/mL的氢化乳清酸和草酸溶液备用。选择72盆长势一致的一年生健康三七苗,分别用上述配制好的氢化乳清酸和草酸溶液灌入根部,氢化乳清酸以含1% DMF的无菌水为对照,草酸以等体积的无菌水为对照,每处理6盆。灌根24 h后用接种针挑取培养7 d的黑斑病菌菌饼(直径为6 mm)接种于三七叶片伤口处,每棵苗接种2片叶片。接种后将所有盆栽放入透明塑料箱中,在26 ℃、光照强度2000 lx、12 h光暗交替条件下保湿培养,4 d后取发病叶片,用扫描仪(Epson perfection V850 Pro scanner)扫描病斑后,利用Adobe Photoshop CS6测量病斑面积。
1. 3 统计分析
试验数据采用SPSS 18.0进行分析;通过Excel 2010和GraphPad Prism 9.4.0进行统计学绘图;通过Metaboanalyst平台(https://www.metaboanalyst.ca/)对差异代谢物进行热图聚类和代谢通路分析。
2 结果与分析
2. 1 接种黑斑病菌后不同时间点对三七内源JA的影响
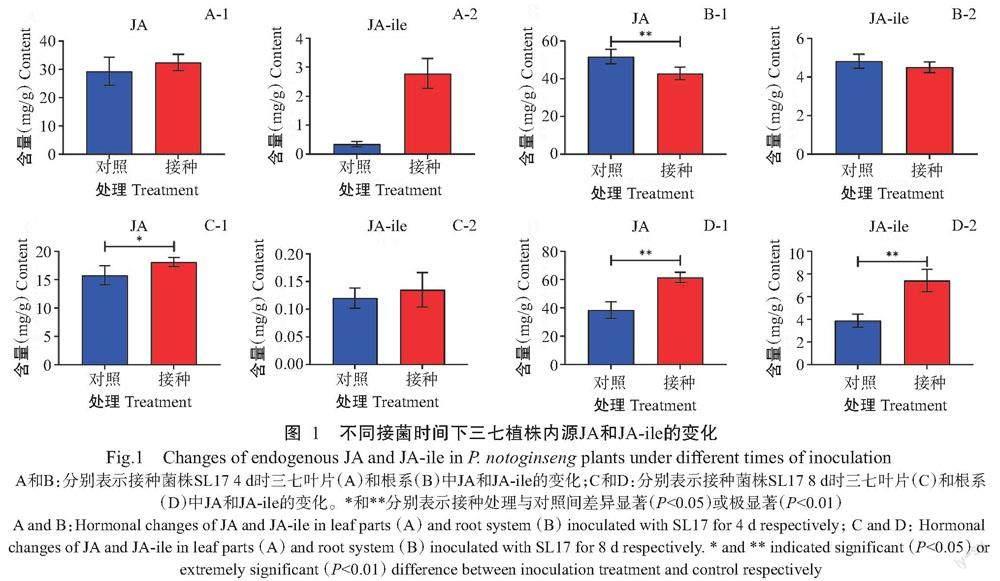
由图1可知,叶部接种菌株SL17后不同时间点对三七植株内源JA含量存在影响。与对照相比,接种4 d后,三七叶片中JA和茉莉酸亮氨酸螯合物(JA-ile)的含量增加(图1-A),但根系中JA和JA-ile含量降低,其中JA含量较对照极显著降低(P<0.01,下同)(图1-B);接种8 d后,叶片中JA和JA-ile含量增加(图1-C),同时,根系中的JA和JA-ile含量极显著增加(图1-D)。
2. 2 外源施用JA后三七根系代谢的变化
2. 2. 1 三七根系代谢物分析 经GC-MS检测所有处理后得到125种代谢物,其中包括糖及衍生物33种、有机酸26种、氨基酸23种、酯类7种、酮类5种、胺类5种、核苷酸及其衍生物4种、烷类3种、多元醇类2种、其他化合物17种。
2. 2. 2 外源施用JA后三七根系差异代谢物分析
2. 2. 2. 1 主成分分析 从PCA得分图可看出(图2),所有样品均处于95%置信区间,说明模型拟合效果较好,且同一处理组中的6组重复相对聚拢,试验重复性相对良好。不同浓度JA处理与CK在主成分上明显分离,并解释了样品总变量的58.98%,表明与对照相比,JA处理明显改变了三七的根系代谢。
2. 2. 2. 2 差异代谢物分析 基于VIP≥1和P<0.05筛选差异代谢物,并对差异代谢物进行热图聚类分析。结果(图3)显示,与CK相比,JA_0.001处理中有1种代谢物表达上调,43种代谢物表达下调;JA_0.010处理中有12种代谢物表达上调,47种代谢物表达下调。当JA浓度为0.001 mmol/L时,有机酸中的草酸含量增加(图3-A);当浓度为0.010 mmol/L时,表现上调的代谢物数量增多,有机酸类(戊酸、十七烷酸、草酸、抗坏血酸)、多元醇(肌醇)、糖类[L-(-)-阿拉伯糖醇、来苏糖]、氨基酸(天门冬氨酸、脯氨酸、瓜氨酸)、核苷酸及其衍生物(双甘肽、氢化乳清酸)在根系中的含量均增加(图3-B)。
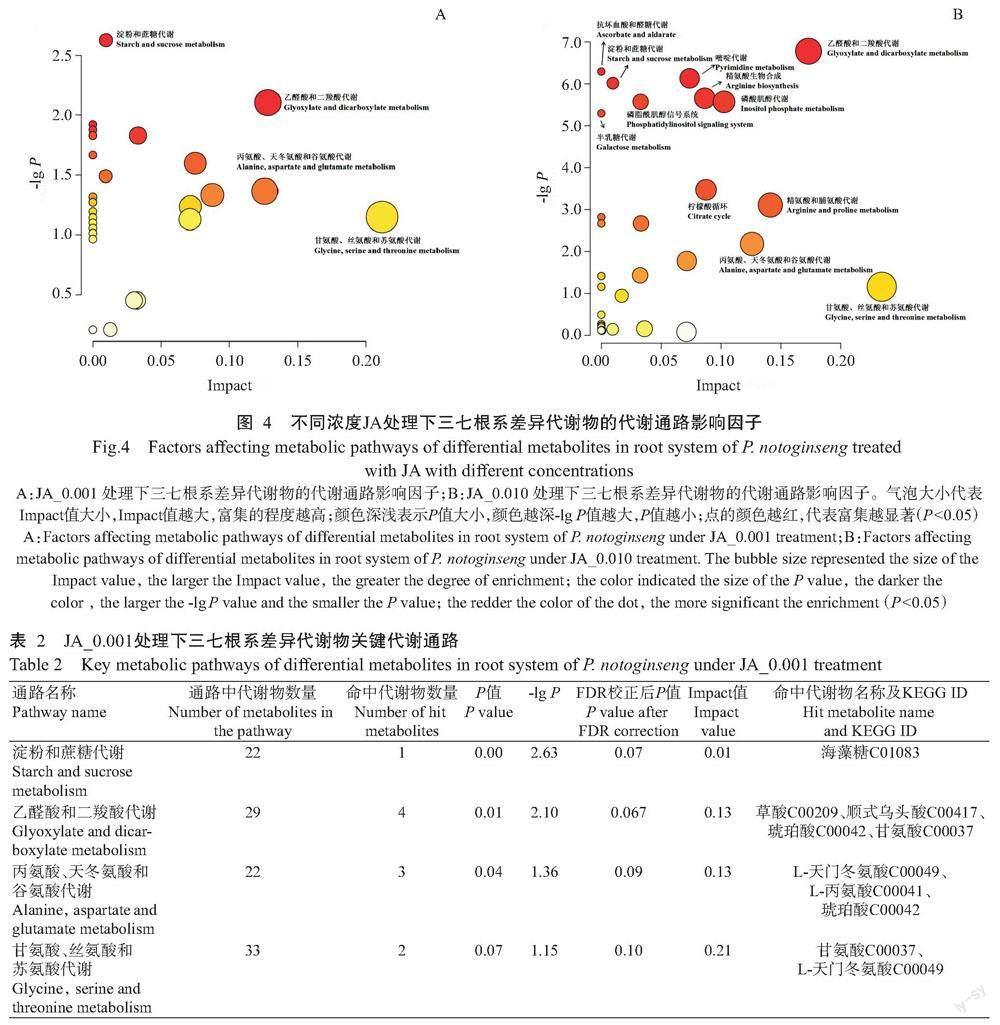
2. 2. 2. 3 差异代谢物代谢通路富集分析 为更直观地反映不同处理下三七根系差异代谢物的变化情况,对JA_0.001和JA_0.010处理的差异代谢物进行代谢通路分析,得到代谢通路影响因子图(图4),JA_0.001处理后发生变化的代谢通路共32条,筛选出Impact>0.1或-lg P值较大的通路共4条,分别为淀粉和蔗糖代谢,乙醛酸和二羧酸代谢,丙氨酸、天冬氨酸和谷氨酸代谢,甘氨酸、丝氨酸和苏氨酸代谢(图4-A)。JA_0.010处理后发生变化的代谢通路共37条,筛选出Impact>0.1或-lg P值较大的通路共12条,分别为抗坏血酸和醛糖代谢,淀粉和蔗糖代谢,磷脂酰肌醇信号系统,半乳糖代谢,嘧啶代谢,精氨酸生物合成,磷酸肌醇代谢,柠檬酸循环,乙醛酸和二羧酸代谢,精氨酸和脯氨酸代谢,丙氨酸、天冬氨酸和谷氨酸代谢,甘氨酸、丝氨酸和苏氨酸代谢。以上结果表明,不同浓度JA喷施三七叶片后,主要影响了根系的糖代谢、氨基酸代谢和有机酸代谢(图4-B)。
富集到JA_0.001处理中4条通路的代谢物有7种,分别是海藻糖、草酸、顺式乌头酸、琥珀酸、甘氨酸、L-天门冬氨酸和L-丙氨酸;其中,琥珀酸、甘氨酸和L-天门冬氨酸参与了2条代谢通路(表2)。富集到JA_0.010处理中12条通路的代谢物有15种,分别是肌醇、抗坏血酸、海藻糖、甘油、氢化乳清酸、5,6-二氢脲嘧啶、草酸、顺式乌头酸、琥珀酸、甘氨酸、D-甘油酸、L-瓜氨酸、L-天门冬氨酸、L-脯氨酸和1,4-丁二胺;其中,肌醇参与了4条代谢通路,L-天门冬氨酸和琥珀酸参与了3条代谢通路(表3)。
2. 2. 2. 4 显著差异代谢物筛选结果 进一步将JA_0.001和JA_0.010处理中显著变化的差异代谢物(VIP≥1,P<0.05,Fold Change>1.5或Fold Change<0.5)进行分析,发现2个浓度中下调的代谢物较多,但随着浓度的增加,显著上调的差异代谢物数量逐渐增多。JA_0.001处理中,显著上调的代谢物为草酸(Fold Change>1.5),JA_0.010处理中,显著上调的代谢物有氢化乳清酸、草酸、瓜氨酸、脯氨酸和抗坏血酸(Fold Change>1.5),其中氢化乳清酸和草酸为极显著上调代谢物(Fold Change>2.0)(图5-A和图5-B)。
2. 3 差异代谢物氢化乳清酸和草酸对三七主要根腐病菌菌丝生长的影响
2. 3. 1 氢化乳清酸对三七主要根腐病菌菌丝生长的影响 由图6可看出,氢化乳清酸对茄腐镰刀菌菌株F3的生长具有一定抑制作用,浓度为0.001、0.100和1.000 μg/mL时能显著抑制菌丝的生长(P<0.05,下同)(图6-A);在0.001和0.010 μg/mL时对恶疫霉菌株D-6无抑制效果,浓度为0.100和10.000 μg/mL时对菌株D-6的菌丝生长具有显著的抑制活性(图6-B);氢化乳清酸对锈腐病菌表现出不同程度的抑制作用,当浓度为0.001、0.010和1.000 μg/mL时能显著抑制菌株RS6的菌丝生长(图6-C),浓度为0.010 μg/mL时对菌株RS8的菌丝生长具有显著的抑制活性(图6-D)。综上所述,氢化乳清酸对三七主要根腐病菌具有一定的抑制作用。
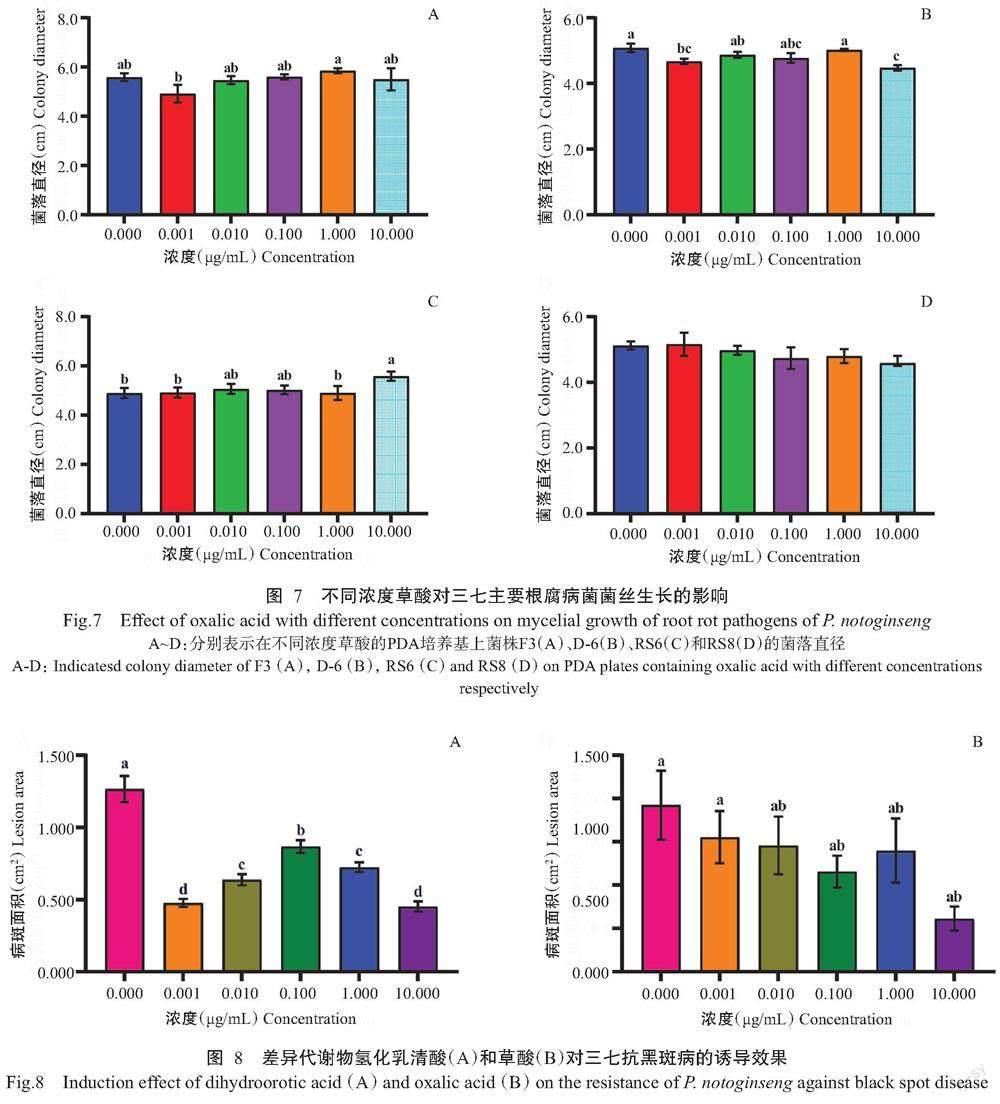
2. 3. 2 草酸对三七主要根腐病菌菌丝生长的影响 由图7可看出,草酸在浓度为0.001 μg/mL时对菌株F3菌丝生长存在明显的抑制作用(图7-A);当浓度为0.001和10.000 μg/mL时对菌株D-6菌丝生长表现出显著的抑制作用(图7-B);草酸对菌株RS6和RS8的菌丝生长无明显的抑制效果。表明草酸对部分三七根腐病菌具有一定的抑制作用。
2. 4 差异代谢物氢化乳清酸和草酸对三七抗黑斑病的诱导效果
经不同浓度氢化乳清酸处理后,三七黑斑病的病斑面积均显著减小。接种4 d后,对照的病斑面积为1.263 cm2,0.001、0.010、0.100、1.000和10.000 μg/mL氢化乳清酸处理后的病斑面积分别比对照小62.32%、49.59%、31.43%、42.83%和64.31%,其中,10.000 μg/mL氢化乳清酸处理后病斑面积最小(图8-A)。
不同浓度草酸处理后,三七黑斑病的病斑面積均减小,但差异不显著(P>0.05)。0.001、0.010、0.100、1.000和10.000 μg/mL草酸处理后的病斑面积分别比对照小19.25%、24.21%、39.87%、27.30%和68.16%,其中,10.000 μg/mL草酸处理后病斑面积最小(图8-B)。以上结果表明,氢化乳清酸和草酸灌根后均能诱导三七对黑斑病菌产生抗性,且浓度为10.000 μg/mL时诱导效果最好。
3 讨论
三七由于其独特的生长环境容易发生多种病害,三七病害的发生直接影响其品质与产量,造成严重的经济损失(崔丹丹等,2020;张雪玲等,2021)。JA是一种重要的植物内源激素,在植物生长、发育和抗逆诱导等方面具有重要的生理功能(毛佳昊等,2021)。有研究发现,病原菌Pseudomonas syringae pv. tomato (Pst)侵染拟南芥后能显著增加植株内JA含量(Yuan et al.,2018)。本研究也发现三七叶部接种黑斑病菌4 d时,JA和JA-ile含量在叶部上调而在根系下调;接种8 d时,不仅叶部JA的含量显著上调,且根系中JA和JA-ile的含量极显著上调,说明三七叶部被黑斑病菌侵染后,也能使JA的含量上调,且随着接种时间的延长,JA在根部被显著积累。
许多研究发现JA可诱导植物代谢改变(王春丽等,2012;孙际薇,2014)。丛枝菌根真菌侵染可促进植物体内JA水平上升,进而诱导次生代谢产物增加,增强植物抗病性(吕亚茹等,2022)。本研究对叶部外源喷施JA处理后的三七根系进行代谢组分析发现,采用GC-MS方法共鉴定出125种代谢物,经OPLS-DA分析后从JA_0.001和JA_0.010处理中分别筛选出44和59种发生变化的差异代谢物,将这些差异代谢物进行代谢通路富集分析,发现大多富集到与氨基酸代谢、有机酸代谢及生物合成相关的通路。研究表明,氨基酸代谢与植物调节渗透势、维持细胞结构及蛋白质合成密切相关(刘晨露,2022),非生物胁迫后,植物细胞会激发氨基酸代谢合成亲水渗透物质,调节渗透压,维持正常生长发育(王佳钰,2020);游离氨基酸在受到胁迫后能直接或间接作出响应(Ashraf and Harris,2004)。有机酸在响应逆境胁迫中发挥着重要作用(郭立泉等,2005),董丽娟(2018)发现山定子幼苗在碱胁迫条件下根系会分泌草酸以降低逆境损伤。综上所述,外源施用JA后能增加三七根系代谢产物含量且富集到相关通路,与前人对植物抗逆性的研究相符(田娇等,2015),推测JA可能通过改变代谢来增强植株抗病性。
植物诱导抗性在所有植物中普遍存在(郑家瑞和李云洲,2022),植物在外源蛋白质、糖类、有机酸等处理后,通过受体识别、信号传导等产生防御反应(万宣伍等,2022)。本研究结果表明,叶部JA处理有助于促进三七根系中氢化乳清酸和草酸含量的增加,这些物质可能在JA诱导植物抗性中发挥重要作用。有研究发现,氢化乳清酸主要参与嘧啶代谢,嘧啶代谢在人类疾病控制和嘧啶的胞内循环利用中具有重要作用,有利于免疫细胞的合成,与抗菌活性密切相关(张海彬,2021);但目前尚无关于氢化乳清酸对植株本身的诱导抗性及对病原菌调控相关的报道,本研究结果可为氢化乳清酸在农业方面的应用提供依据。草酸是一种在动植物、真菌中普遍存在的代谢物,主要参与乙醛酸和二羧酸代谢,提高生物体利用乙酰-CoA的能力,对植物的生长发育起着重要作用(Dutton and Evans,1996)。有研究发现,草酸具有很强的抗氧化能力(Kayashima and Katayama,2002)。朱丽琴等(2013)发现草酸处理辣椒果实后会增强其超氧化物歧化酶(SOD)和过氧化氢酶(CAT)活性,延缓辣椒果实衰老;二者的变化可能与植物响应病原菌侵染及调控植物抗性有关,但其作用尚不明确。罗丽芬等(2019)发现外源添加草酸能增强三七根系的抗氧化能力,有效缓解皂苷Rg1对三七生长的自毒活性。徐翠等(2022)发现浓度为25 mmol/L的草酸处理狗尾草种子25 min,可有效促进收获当年和储藏1年种子的萌发及生长。在本研究中,氢化乳清酸和草酸不仅对三七主要根腐病菌具有不同程度的抑制作用,外源灌根后还能提高三七对叶部黑斑病的抗性。但本研究仅测定了三七根系代谢物中氢化乳清酸和草酸在低浓度下对三七抗黑斑病的诱导抗性及对根腐病菌生长的影响,后续还需对这2种物质在根系分泌物中的含量进行检测,以便更好地研究其功能。另外,本研究JA外源喷施三七后,三七根系中许多代谢物被下调,后续亟需对下调的化合物开展深入研究,以明确其功能,以期对外源施用JA后三七根系代谢物的功能进行更全面的解析。
另有研究发现,植物可能利用根部代谢物对土壤中的微生物进行选择(Akhter et al.,2015),并可通过改变代谢和分泌来影响根际微生物的种类和丰度(Turner et al.,2013)。Carvalhais等(2015)发现,外源添加JA可改变土壤微生物的群落结构;Huang等(2019)发现三萜类化合物通过定向富集或抑制特定种类细菌的生长,以调控根际微生物的种群和功能;Voges等(2019)发现香豆素能抑制假单胞菌属生长。植物能在叶部病原体入侵后释放特定的根系分泌物包括长链脂肪酸、氨基酸、短链有机酸和糖以募集有益的根际微生物,从而保护寄主或后代植物(Wen et al.,2020)。有益的根际微生物也能通过诱导植物的系统抗性(ISR)来抑制发病(Yuan et al.,2018)。Kost等(2014)发现,有益菌株Burkholderia phytofirmans PsJN定殖羽扇豆和玉米的能力与其利用草酸为碳源的能力密切相关。本研究中草酸灌根后能提高三七的抗病性,可能是在三七根部募集到了某些有益菌来保护宿主植物。因此,后续亟需深入研究外源施用JA后三七能否通过调控根部代谢物及分泌物中的氢化乳清酸和草酸含量来特异性调控根际土壤微生物群落结构和功能,从而深入解析外源施用JA增强三七植株抗性的机制。
4 结论
外源喷施JA可改变三七根系代谢,诱导氢化乳清酸和草酸的积累,进而增强三七对黑斑病的抗性,并对三七主要根腐病菌具有一定的抑制活性。
参考文献:
崔丹丹,杨巧玲,张俊莲,张峰. 2020. 茉莉酸抑制剂对马铃薯离体块茎形成和发育的影响[J]. 甘肃农业大学学报,55(1):82-90. [Cui D D,Yang Q L,Zhang J L,Zhang F. 2020. Effect of jasmonic acid inhibitor on potato tuber formation and development in vitro[J]. Journal of Gansu Agricultural University,55(1):82-90.] doi:10.13432/j.cnki.jgsau.2020.01.011.
董丽娟. 2018. 不同山定子幼苗耐碱性评价与碱胁迫下有机酸积累的差异机理分析[D]. 杨凌:西北农林科技大学. [Dong L J. 2018. Alkali tolerance of different Malus baccata accessions and the differences of organic acid accumulation of two accessions under alkali stress[D]. Yang-ling:Northwest A & F University.]
郭立泉,石德成,马传福. 2005. 植物在响应逆境胁迫过程中的有机酸代谢调节及分泌现象[J]. 长春教育学院学报,(3):19-24. [Guo L Q,Shi D C,Ma C F. 2005. Regulation of organic acid metabolism and secretion phenomena in plants in response to adversity stress[J]. Journal of Changchun Education Institute,(3):19-24.] doi:10.3969/j.issn.1671-6531.2005.03.007.
蔣妮,覃柳燕,叶云峰. 2011. 三七病害研究进展[J]. 南方农业学报,42(9):1070-1074. [Jiang N,Qin L Y,Ye Y F. 2011. Research advances in disease of Panax notoginseng[J]. Journal of Southern Agriculture,42(9):1070-1074.] doi:10.3969/j.issn.2095-1191.2011.09.010.
李颜,黄益宗,保琼莉,黄永春,张盛楠. 2022. 外源茉莉酸对水稻幼苗根系砷积累及抗逆应答效应[J]. 环境科学,43(9):4831-4838. [Li Y,Huang Y Z,Bao Q L,Huang Y C,Zhang S N. 2022. Effects of exogenous jasmonic acid on arsenic accumulation and response to stress in roots of rice seedlings[J]. Environmental Science,43(9):4831-4838.] doi:10.13227/j.hjkx.202111296.
刘晨露. 2022. 富平楸子根系氨基酸代谢对碱胁迫的响应机制研究[D]. 杨凌:西北农林科技大学. [Liu C L. 2022. Response mechanisms of amino acid metabolism in roots of Malus prunifolia under alkali stress[D]. Yangling:Northwest A & F University.] doi:10.27409/d.cnki.gxbnu. 2022. 000864.
罗丽芬,张俊星,钏有聪,李怡文,郝敏文,顾红蕊,朱书生,杨敏. 2019. 外源草酸对三七皂苷Rg1自毒效应的缓解作用及机制研究[J]. 西北农林科技大学学报(自然科学版),47(4):101-108. [Luo L F,Zhang J X,Chuan Y C,Li Y W,Hao M W,Gu H R,Zhu S S,Yang M. 2019. Effect and mechanism of exogenous oxalic acid on alleviating autotoxicity of Rg1 to Panax notoginseng[J]. Journal of Northwest A & F University (Natural Science Edition),47(4):101-108.] doi:10.13207/j.cnki.jnwafu.2019.04.013.
罗群,游春梅,官会林. 2010. 环境因素对三七生长影响的分析[J]. 中国西部科技,9(9):7-8. [Luo Q,You C M,Guan H L. 2010. Analysis of influences of environmental factors on the growth of Panax notoginseng[J]. Science and Technology of West China,9(9):7-8.] doi:10.3969/j.issn.1671-6396.2010.09.004.
吕亚茹,严俊鑫,王菲,谭怡,蔺吉祥. 2022. 茉莉酸介导的丛枝菌根真菌诱导植物抗病性研究进展[J]. 中国生物防治学报,38(3):726-738. [Lü Y R,Yan J X,Wang F,Tan Y,Lin J X. 2022. Research progress of jasmonic acid-mediated arbuscular mycorrhizal fungi of plant disease resistance[J]. Chinese Journal of Biological Control,38(3):726-738.] doi:10.16409/j.cnki.2095-039x.2022.02.018.
毛佳昊,熊晓辉,卢一辰. 2021. 茉莉酸调控植物应对逆境胁迫作用的研究进展[J]. 生物加工过程,19(4):413-419. [Mao J H,Xiong X H,Lu Y C. 2021. Advances in the regulation of plant stress response by jasmonic acid[J]. Chinese Journal of Bioprocess Engineering,19(4):413-419.] doi:10.3969/j.issn.1672-3678.2021.04.008.
孙际薇. 2014. 茉莉酸甲酯对曼陀罗毛状根的生长及次生代谢产物产生的影响[D]. 重庆:西南大学. [Sun J W. 2014. Effects of methyl jasmonate on the Datura stramonium L. hairy root growth and secondary metabolites[D]. Chongqing:Southwestern University.]
孙萌,叶丽琴,张子龙. 2015. 三七连作障碍成因及其控制研究进展[J]. 山地农业生物学报,34(3):63-67. [Sun M,Ye L Q,Zhang Z L. 2015. Progress on the cause of continuous cropping obstacle Panax notoginseng and its countermeasures[J]. Journal of Mountain Agriculture and Biology,34(3):63-67.] doi:10.15958/j.cnki.sdnyswxb. 2015.03.012.
田嬌,刘园,房敏峰. 2015. 外源茉莉酸类激素对药用植物次生代谢的影响研究[J]. 天然产物研究与开发,27(1):185-190. [Tian J,Liu Y,Fang M F. 2015. Review on the influence of exogenous jasmonates on medicinal plant secondary metabolism[J]. Natural Product Research and Development,27(1):185-190.] doi:10.16333/j.1001-6880. 2015.01.037.
万宣伍,田卉,张伟,杨娟,董义霞,刘昌黎,杜学英,谭康,唐自然. 2022. 植物诱导抗性的机理及应用[J]. 植物医学,1(1):18-25. [Wan X W,Tian H,Zhang W,Yang J,Dong Y X,Liu C L,Du X Y,Tan K,Tang Z R. 2022. Mechanism and application of induced resistance in plant[J]. Plant Health and Medicine,1(1):18-25.] doi:10. 13718/j.cnki.zwyx.2022.01.003.
王春丽,梁宗锁,李殿荣,杨建利. 2012. 水杨酸和茉莉酸甲酯对丹参幼苗叶片显微结构、光合及非结构糖积累的影响[J]. 植物科学学报,30(5):501-510. [Wang C L,Liang Z S,Li D R,Yang J L. 2012. Influence of salicylic acid and methyl jasmonate on leaf microstructure,leaf photosynthesis,and non-structural sugar accumulation of Salvia miltiorrhiza Bunge seedlings[J]. Plant Science Journal,30(5):501-510.] doi:10.3724/SP.J.1142.2012.50501.
王佳鈺. 2020. 高浓度CO2和铅胁迫下水稻幼苗代谢组学响应研究[D]. 沈阳:沈阳师范大学. [Wang J Y. 2020. Metabolomic responses of rice seedlings to elevated CO2 concentration and/or Pb stress[D]. Shenyang:Shenyang Normal University.] doi:10.27328/d.cnki.gshsc.2020.000 026.
王小其. 2020. 中药三七对血液系统的药理药效研究进展[J]. 山西医药杂志,49(10):1231-1233. [Wang X Q. 2020. Advances in the pharmacological and pharmacologi-cal effects of the Chinese medicine Panax notoginseng on the blood system[J]. Shanxi Medical Journal,49(10):1231-1233.] doi:10.3969/j.issn.0253-9926.2020.10.015.
王云锋,王长秘,李春琴,刘林,李晓杰,李晓疆,杨静. 2018. 稻瘟病菌侵染时水稻防御体系对外源茉莉酸的响应分析[J]. 南方农业学报,49(7):1324-1331. [Wang Y F,Wang Z M,Li C Q,Liu L,Li X J,Li X J,Yang J. 2018. Response of rice defense system to exogenous jasmonic acids during rice blast strain infecting rice[J]. Journal of Southern Agriculture,49(7):1324-1331.] doi:10.3969/j.issn.2095-1191.2018.07.10
魏婷,罗辰瑶,李红,李鲜,王雨凌,王雪妍. 2021. 外源茉莉酸甲酯对番茄幼苗生长及镉抗性的影响[J]. 陕西科技大学学报,39(4):15-20. [Wei T,Luo C Y,Li H,Li X,Wang Y L,Wang X Y. 2021. Effects of exogenous methyl jasmonate on the growth and Cd tolerence of tomato seedlings[J]. Journal of Shaanxi University of Science & Technology,39(4):15-20.] doi:10.19481/j.cnki.issn2096-398x.2021.04.003.
徐翠,姜娜,旷思萍,蒋金娟,罗富成,祖艳群. 2022. 外源草酸对不同年限狗尾草种子萌发及生理生化的影响[J]. 草地学报,30(9):2381-2390. [Xu C,Jiang N,Kuang S P,Jiang J J,Luo F C,Zu Y Q. 2022. Effect of exogenous oxalic acid on germination and physiological biochemistry characteristics of Setaria sphacelate ‘Narok’ seeds of different years[J]. Acta Agrestia Sinica,30(9):2381-2390.] doi:10.11733/j.issn.1007-0435.2022.09.018.
严加坤,严荣,汪亚妮. 2019. 外源茉莉酸甲酯对盐胁迫下玉米根系吸水的影响[J]. 广东农业科学,46(1):1-6. [Yan J K,Yan R,Wang Y N. 2019. Impact of exogenous methyl jasmonate on water absorption of maize roots under salt stress[J]. Guangdong Agricultural Sciences,46(1):1-6.] doi:10.16768/j.issn.1004-874x.2019.01.001.
張海彬. 2021. 微生物中嘧啶代谢途径的研究进展[J]. 生物技术,31(6):619-624. [Zhang H B. 2021. Research progress of pyrimidine metabolism in microbes[J]. Biotechnology,31(6):619-624.] doi:10.16519/j.cnki.1004-311x. 2021.06.0095.
张金丽. 2022. 三七根腐病及生物防治研究进展[J]. 农村经济与科技,33(2):50-52. [Zhang J L. 2022. Progress of Panax notoginseng root rot disease and biological control[J]. Rural Economy and Science-Technology,33(2):50-52.] doi:10.3969/j.issn.1007-7103.2022.02.016.
张明菊,朱莉,夏启中. 2021. 植物激素对胁迫反应调控的研究进展[J]. 湖北大学学报(自然科学版),43(3):242-253. [Zhang M J,Zhu L,Xia Q Z. 2021. Research progress on the regulation of plant hormones to stress responses[J]. Journal of Hubei University (Natural Science Edition),43(3):242-253]. doi:10.3969/jjssn.1000-23750202103003.
张帅,罗文才,符成悦,陈斌,张立敏. 2020. 不同种植年限和生育期三七上桃蚜体内蛋白含量及主要酶活性研究[J]. 南方农业学报,51(8):1985-1991. [Zhang S,Luo W C,Fu C Y,Chen B,Zhang L M. 2020. Protein content and the activities of main enzymes in Myzus persicae (Sulzer)feeding on Panax notoginseng at growth period at diffe-rent planting periods[J]. Journal of Southern Agriculture,51(8):1985-1991.] doi:10.3969/j.issn.2095-1191.2020. 08.025.
张雪玲,臧春华,郭汝悦,段毅超,王延刚,郭中信,张振中,任雪玲. 2021. 生物菌肥对连作障碍三七根际和根茎内细菌群落的影响[J]. 河南农业科学,50(12):78‐91. [Zhang X L,Zang C H,Guo R Y,Duan Y C,Wang Y G,Guo Z X,Zhang Z Z,Ren X L. 2021. Effects of biological fertilizer on rhizosphere and endophyticbacterial community of Panax notoginseng with continuouscropping disorder[J]. Journal of Henan Agricultural Sciences,50(12):78-91.] doi:10.15933/j.cnki.1004-3268.2021.12.009.
赵显阳,盘柳依,陈明,付永琦,向妙莲,曾晓春. 2018. 茉莉酸甲酯对辣椒抗青枯病的诱导效应及抗氧化酶活性的影响[J]. 植物保护学报,45(5):1103-1111. [Zhao X Y,Pan L Y,Chen M,Fu Y Q,Xiang M L,Zeng X C. 2018. Inductive effect of methyl jasmonate to bacterial wilt and the effects on the activities of antioxidant enzymes in pepper seedlings[J]. Acta Phytophylacica Sinica,45(5):1103-1111.] doi:10.13802/j.cnki.zwbhxb.2018.2018128.
赵泽花,马祥,董文科,孙国,周姝婧,田丽华,马晖玲. 2020. 外源茉莉酸甲酯诱导草地早熟禾对白粉病抗性的研究[J]. 草原与草坪,40(2):59-66. [Zhao Z H,Ma X,Dong W K,Sun G,Zhou S J,Tian L H,Ma H L. 2020. Study on resistance of Poa pratensis to powdery mildew induced by exogenous methyl jasmonate[J]. Grassland and Turf,40(2):59-66.] doi:10.13817/j.cnki.cyycp.2020. 02.009.
郑家瑞,李云洲. 2022. 植物诱导抗性研究进展[J]. 山地农业生物学报,41(2):51-58. [Zheng J R,Li Y Z. 2022. Research progress on induced resistance in plants[J]. Journal of Mountain Agriculture and Biology,41(2):51-58.] doi:10.15958/j.cnki.sdnyswxb.2022.02.008.
郑素慧,何庆,张健,关军锋,秦南南,杨怡靖铭,刘雪艳,吴斌. 2022. 茉莉酸甲酯对红地球葡萄采后贮藏品质和病害的影响[J]. 新疆农业科学,59(1):190-198. [Zheng S H,He Q,Zhang J,Guan J F,Qin N N,Yang Y J M,Liu X Y,Wu B. 2022. Effects of exogenous methyl jasmonate on quality and diseases of grapefruit[J]. Xinjiang Agricultural Science,59(1):190-198.] doi:10.6048/j.issn. 1001-4330.2022.01.022.
朱麗琴,张伟,汪伟,陈宇欢. 2013. 外源草酸对辣椒保鲜效果和抗氧化防御系统的影响[J]. 江西农业大学学报,35(3):521-524. [Zhu L Q,Zhang W,Wang W,Chen Y H. 2013. Effects of exogenous oxalic acid on preservation and antioxidant defense system of pepper fruits[J]. Acta Agricultural Universitatis Jiangxiensis,35(3):521-524.] doi:10.13836/j.jjau.2013093.
邹石龙. 2018. 三七种植现状及其种植技巧分析[J]. 农业与技术,38(4):119. [Zou S L. 2018. Analysis of the current situation of Panax notoginseng cultivation and its cultivation techniques[J]. Agriculture and Technology,38(4):119.] doi: 10.11974/nyyjs.20180233099.
Akhter A,Hage-Ahmed K,Soja G,Steinkellner S. 2015. Compost and biochar alter mycorrhization,tomato root exudation,and development of Fusarium oxysporum f. sp. lycopersici[J]. Frontiers in Plant Science,6:529. doi:10. 3389/fpls.2015.00529.
Ashraf M,Harris P J G. 2004. Potential biochemical indicators of salinity tolerance in plants[J]. Plant Science,166(1):3-16. doi:10.1016/j.plantsci.2003.10.024.
Awan S A,Khan I,Rizwan M,Zhang X,Brestic M,Khan A,El-Sheikh M A,Alyemeni M N,Ali S,Huang L K. 2020. Exogenous abscisic acid and jasmonic acid restrain polye-thylene glycolinduced drought by improving the growth and antioxidative enzyme activities in pearl millet[J]. Physiol Plant,172(2):809-819. doi:10.1111/ppl.13247.
Carvalhais L C,Dennis P G,Badri D V,Kidd B N,Vivanco J M,Schenk P M. 2015. Linking jasmonic acid signaling,root exudates,and rhizosphere microbiomes[J]. Molecular Plant and Microbe Interaction,28:1049-1058. doi:10.1094/MPMI-01-15-0016-R.
Dutton M V,Evans C S. 1996. Oxalate production by fungi:Its role in pathogenicity and ecology in the soil environment[J]. Canadian Journal of Microbiology,42:881-895. doi:10.1139/m96-114.
Han Z L,Zhang C,Zhang H,Duan Y,Zou Z W,Zhou L,Zhu X J,Fang W P,Ma Y C. 2022. CsMYB transcription factors participate in jasmonic acid signal transduction in response to cold stress in tea plant (Camellia sinensis) [J]. Plants,11(21):2869. doi:10.3390/plants11212869.
Huang A C,Jiang T,Liu Y X,Bai Y C,Reed J,Qu B Y,Goo-ssens A,Nutzmann H W,Bai Y,Osbourn A. 2019. A specialized metabolic network selectively modulates Arabidopsis root microbiota[J]. Science,364(6440):546. doi:10.1126/science.aau6389.
Kayashima T,Katayama T. 2002. Oxalic acid is available as a natural antioxidant in some systems[J]. Biochimica Biophysica Acta,1573(1):1-3. doi:10.1016/S0304-4165(02)00338-0.
Kost T,Stopnisek N,Agnoli K,Eberl L,Weisskopf L. 2014. Oxalotrophy,a widespread trait of plant-associated Burkholderia species,is involved in successful root colonization of lupin and maize by Burkholderia phytofirmans[J]. Frontiers in Microbiology,4:1-9. doi:10.3389/fmicb. 2013.00421.
Marquis V,Smirnova E,Graindorge S,Delcros P,Villette C,Zumsteg J,Heintz D,Heitz T. 2022. Broad-spectrum stress tolerance conferred by suppressing jasmonate signaling attenuation in Arabidopsis JASMONIC ACID OXIDASE mutants[J]. The Plant Journal,109(4):856-872. doi:10.1111/tpj.15598.
Turner T R,Ramakrishnan K,Walshaw J,Heavens D,Alston M,Swarbreck D,Osbourn A,Grant A,Poole P S. 2013. Comparative metatranscriptomics reveals kingdom level changes in the rhizosphere microbiome of plants[J]. The ISME Journal,7:2248-2258. doi:10.1038/ismej.2013.119.
Voges M J E E,Bai Y,Schulze-Lefert P,Sattely E S. 2019. Plant-derived coumarins shape the composition of a synthetic root microbiome[J]. Proceedings of the National Academy of Sciences of the United States of America,116:12558-12565. doi:10.1073/pnas.1820691116.
Wang F Y,Zhang X M,Wei M T,Wang Y,Liang Z S,Xia P G. 2022. Appropriate crop rotation alleviates continuous cropping barriers by changing rhizosphere microorga-nisms in Panax notoginseng[J]. Rhizosphere,23:100568. doi:10.1016/J.RHISPH.2022.100568.
Wang X,Li Q,Xie J J,Huang M,Cai J,Zhou Q,Dai T B,Jiang D. 2021. Abscisic acid and jasmonic acid are involved in drought priminginduced tolerance to drought in wheat[J]. The Crop Journal,9(1):120-132. doi:10.1016/j.cj.2020.06.002.
Watanabe Y,Tatsuno I. 2021. Omega-3 polyunsaturated fatty acids focusing on eicosapentaenoic acid and docosahexaenoic acid in the prevention of cardiovascular diseases:A review of the state-of-the-art[J]. Expert Review of Clinical Pharmacology,14(1):79-93. doi:10.1080/17512433. 2021.1863784.
Wen T,Zhao M L,Yuan J,Kowalchuk G A,Shen Q R. 2020. Root exudates mediate plant defense against foliar pathogens by recruiting beneficial microbes[J]. Soil Ecology Letters,3(1):42-51. doi:10.1007/S42832-020-0057-Z.
Yang B X,Chen M,Wang T T,Chen X,Li Y H,Wang X,Zhu W,Xia L A,Hu X J,Tian J K. 2019. A metabolomic strategy revealed the role of JA and SA balance in Clema-tis terniflora DC. response to UVB radiation and dark[J]. Physiologia Plantarum,167(2):232-249. doi:10. 1111/ppl.12883.
Yang Y X,Wu C Q,Ahammed G J,Wu C J,Yang Z,Wan C M,Chen J P. 2018. Red light-induced systemic resistance against root-knot nematode is mediated by a coordinated regulation of salicylic acid,jasmonic acid and redox signaling in watermelon[J]. Frontiers in Plant Science,9:899. doi:10.3389/fpls.2018.00899.
Yuan J,Zhao J,Wen T,Zhao M L,Li R,Goossens P,Huang Q W,Bai Y,Vivanco J M,Kowalchuk G A,Berendsen R L,Shen Q R. 2018. Root exudates drive the soil-borne legacy of aboveground pathogen infection[J]. Microbio-me,6(1):156. doi:10.1186/s40168-018-0537-x.
(責任编辑 麻小燕)
收稿日期:2022-10-05
基金项目:国家自然科学基金项目(32060719);云南省中青年后备人才项目(202005AC160045);云南省创新团队项目(202105AE 160016);云南省农林联合重点项目(202101BD070001-003)
通讯作者:杨敏(1981-),https://orcid.org/0000-0003-2658-3075,博士,教授,主要从事三七连作障碍研究工作,E-mail:yangminscnc @126.com
第一作者:王正平(1999-),https://orcid.org/0000-0002-6636-7927,研究方向为三七连作障碍,E-mail:1273459195@qq.com
猜你喜欢
农业科技通讯(2023年1期)2023-02-12
作物学报(2022年4期)2022-02-10
中国中药杂志(2016年22期)2017-02-13
材料科学与工程学报(2016年4期)2017-01-15
饮食科学(2016年3期)2016-07-04
饮食科学(2016年3期)2016-07-04
养生大世界(2016年5期)2016-05-19
食品工业科技(2014年13期)2014-03-11
食品科学(2013年22期)2013-03-11
食品科学(2013年8期)2013-03-11
