苹果酸-天冬氨酸代谢对水稻镉吸收转运特性的影响
2023-11-09 06:49张昕王惠君薛卫杰王常荣张长波黄永春刘仲齐
农业环境科学学报 2023年10期
张昕,王惠君,薛卫杰,王常荣,张长波,黄永春,刘仲齐
(农业农村部环境保护科研监测所农业农村部产地环境污染防控重点实验室,天津 300191)
镉是重金属污染农田中最常见的有害元素。在我国南方镉污染农田中,稻米中的镉含量极易超过国家规定的限量标准(0.20 mg·kg-1,GB 2762—2017)[1-2]。镉在稻米中积累会显著降低稻米中谷氨酸含量,从而影响其他氨基酸的合成[3]。水稻是耐镉能力较强的粮食作物,能将土壤中的镉富集在根系和茎叶等营养器官中,水稻开花后,营养器官中的镉随其他矿质元素一起转运到发育籽粒中,品种类型和灌浆期的气候条件对镉向稻米中的转运有显著影响[3-5]。
在镉胁迫环境中,植物体内活性氧(ROS)代谢很容易受到干扰,镉离子通过扰乱电子传递过程,产生大量的ROS 如超氧阴离子、羟基、H2O2等,进而引发植物氧化损伤[6-7]。叶绿体是合成ROS 的主要细胞器,叶绿体中的烟酰胺腺嘌呤二核苷酸(NADH)通过苹果酸脱氢酶(MDH),将草酰乙酸(OAA)还原为苹果酸,苹果酸从叶绿体转运到细胞质中,在过氧化物酶体或线粒体中又被氧化成OAA,同时产生NADH和ROS[8-9]。苹果酸在叶绿体和线粒体等细胞器间的穿梭和循环称为苹果酸调节(Malate valve),其在维持植物细胞内的能量平衡、pH 和渗透势稳定、清除细胞内过量NADH 和ROS等方面发挥着重要作用[9-10]。外源添加苹果酸能显著提高植物生长量、增加净光合速率、减少过氧化氢积累、增强根系活性等,进而减轻镉的毒害作用[11-13]。由此可见,苹果酸在细胞器间的转运和循环是植物细胞在逆境条件下调控能量平衡的重要机制。
苹果酸循环与氨基酸代谢有密切的关系。三羧酸循环和乙醛酸循环代谢过程中产生的苹果酸,可以通过氧化作用转化成OAA,后者在天冬氨酸氨基转移酶(AST)的催化下转化成天冬氨酸,进而合成谷氨酸、赖氨酸、苏氨酸等一系列氨基酸[9,14-15]。这些氨基酸能够通过螯合作用降低镉的生物活性,抑制镉向稻米中转运[4,16]。此外,苹果酸还具有补充和平衡三羧酸循环其他代谢产物的功能,从而可以保证氨基酸和糖代谢对有机酸的需求[17-18]。植物的根、茎、叶、花和果实等多种器官都能通过苹果酸在液泡中的储存和释放过程来调控有机酸、氨基酸、脂肪酸和可溶性糖之间的转化,同时调控细胞渗透势和含氮化合物的转运等过程[18-19]。
随着植物体内镉含量的增加,根、茎、叶等器官中苹果酸和柠檬酸的含量显著下降[20-21]。外源补充苹果酸,能否促进苹果酸-天冬氨酸的转化过程并消除镉的毒性效应,尚未见相关研究报道。本研究采用根际添加和叶面喷施苹果酸的方法,对苹果酸通过氨基酸代谢缓解镉毒害的生理机制进行了探讨。
1 材料与方法
1.1 试验材料
以湖南大面积种植的籼稻品种中早35 为材料,在农业农村部环境保护科研监测所人工气候室和日光温室分别进行水培试验和盆栽试验。盆栽土壤为来自广西壮族自治区河池市某地的水稻土,其基本理化性质为:pH 6.53,有机质含量34.38 g·kg-1,全氮0.164%,全磷0.024%,全钾1.20%,速效钾含量84.96 mg·kg-1,速效磷含量17.10 mg·kg-1,阳离子交换量7.82 cmol·kg-1,镉含量0.69 mg·kg-1。
1.2 水培试验方法
选取饱满的水稻种子,用H2O2溶液进行种子表面消毒后,将种子均匀铺至育苗盘内,放置在人工气候室中进行发芽培养。水稻幼苗长至两叶一心时,转移至1/2 Hoagland营养液中培养7~10 d。当多数水稻幼苗进入三叶一心期后,从中挑选大小均匀的幼苗,转移到去离子水中饥饿处理24 h,再分别转移至含有2.7µmol·L-1Cd的1/10 Hoagland营养液中处理7 d,在处理液中分别添加0、0.5、1.0 mmol·L-1和1.5 mmol·L-1苹果酸(Cd、Cd+MA1、Cd+MA2 和Cd+MA3),用KOH 和HNO3调节pH 为5.5~6.0,每个处理重复3 次,每个重复15株苗。处理液每隔1 d更换1次。处理结束后,先将幼苗根系浸泡在5 mmol·L-1CaCl2溶液中20 min,去除表面吸附的离子,再用去离子水漂洗干净。从各重复中随机取5 株水稻幼苗,用根系扫描仪(EPSON STD 1600,winRhizo system V.4.0b)扫描根系,测定水稻幼苗的总根长、表面积和根尖数。然后将各处理的水稻根系与地上部分开,杀青(105°C,15 min)后烘干待用。人工气候室条件参数:昼夜时间为16 h/8 h,昼夜温度为25 ℃/20 ℃,白天光照105µmol·m-2·s-1,相对湿度60%。
1.3 盆栽试验方法
水稻种子在1%的次氯酸钠溶液中浸泡15 min后用去离子水洗净,播种于育苗盘中,根据苗龄,依次浇灌1/10~1/2的Hoagland 营养液,幼苗长到五叶一心之后,选取长势均匀的苗,移栽到装有5 kg 土壤的塑料盆中,每盆9 株。水稻开花期进行叶面喷施处理:处理1 喷施去离子水(CK);处理2 喷施2 次5 mmol·L-1苹果酸(MA1);处理3 喷施3 次5 mmol·L-1苹果酸(MA2),每个处理重复4 次,每次喷施总量为30 mL。水稻成熟期收取水稻籽粒、穗颈、旗叶、节和茎秆等样品,参照张雅荟等[22]的方法进行样品前处理后用于测定镉含量、必需元素含量和籽粒氨基酸含量等指标。
1.4 镉含量及亚细胞分布的测定
水培试验的水稻幼苗根系和地上部以及盆栽试验收获的水稻各器官,分别烘干后磨成粉。参照Xue等[16]的方法,分别称取0.5 g 样品于消解管中,加入7 mL HNO3摇匀,室温下静置12 h。将消解管放入电热消解仪ED54上进行消解,温度110 ℃,加热2.5 h后,冷却至室温,在消煮管内加入1 mL H2O2摇匀,110 ℃继续加热1.5 h。将消解管内的液体于170 ℃下赶酸至1 mL以内。再将消解液稀释并转移至25 mL 容量瓶定容,用ICP-MS(Agilent 7500a,美国)测定样品中镉含量。
镉的亚细胞分布方法主要参照Han 等[23]的方法。从收取的水稻幼苗根系和地上部鲜样中,分别称取0.25 g 和0.50 g 置于研钵中,加入提取缓冲液进行充分研磨后使之成匀浆液。然后在3 000 r·min-1下离心15 min,沉淀为细胞壁组分(F1)。取上清液在15 000 r·min-1下离心30 min,沉淀部分为细胞器组分(F2),上清液为细胞液组分(F3)。整个操作过程在4 ℃下进行。提取缓冲液组成为:250 mmol·L-1蔗糖,50 mmol·L-1Tris-HCl(pH 7.5)和1.0 mmol·L-1二硫赤鲜醇。将收集到的3 种亚细胞组分分别放在70 ℃电热板上蒸干至恒质量,然后测定其中的镉含量。
1.5 酶活性测定
水稻开花期标记单穗的开花时间,按照开花后5、10、15、20、25、30 d的间隔,从每个重复中分别剪取发育正常的稻穗3 个,在液氮中研磨成匀浆,从中取0.20 g 左右的匀浆转移到离心管中,加入1.8 mL 缓冲液(pH 7.4,0.05 mol·L-1Tris-HCl)在冰上静置5 min,然后离心10 min(4 ℃,12 000 r·min-1),上清液即为酶的粗提液。MDH 和AST 的活性用试剂盒(北京索莱宝科技有限公司)测定。MDH 的活性单位用每小时催化产生1µmol·L-1NADH 的酶量来表示,AST 的活性单位用每小时催化产生1 µmol·L-1丙酮酸的酶量来表示。
1.6 氨基酸和巯基化合物的测定
样品中的游离氨基酸依据Xue 等[5]的方法提取。将植物样品晒干磨碎后过筛,称取0.20 g 加入2.0 mL去离子水,然后超声处理30 min,离心(4 ℃,10 000 r·min-1)10 min 后取上清液,残渣再用2.0 mL 去离子水复提,将两次上清液混合均匀过0.22µmol·L-1滤膜后待测。
巯基化合物测定参照Han 等[23]的方法。分别称取0.20 g 根系和地上部鲜样,充分研磨后,加入1.8 mL 的提取缓冲液转移到离心管中,离心(4 ℃,12 000 r·min-1)10 min 后,将上清液轻轻地倒入新的离心管中,加入650 µL HEPES 缓冲液和25 µL TCEP 溶液,混合均匀后,在室温(25 ℃)下预培养5 min,再加入20 µL mBBr 溶液,在黑暗条件下(25 ℃)衍生反应30 min,用1.0 mol·L-1的MSA 100 µL 终止衍生反应,液体用0.22µm 过滤膜过滤后,用高效液相色谱仪(Agilent 1200,美国)测定谷胱甘肽(GSH)和植物螯合肽(PCs)的含量。
1.7 数据统计及分析
采用Excel进行相关数据的计算、统计与处理、制图。用SPSS 20.0 进行统计分析,新复极差法(Duncan′s)进行多重比较、差异显著性检验。
转移因子(TF)为相邻器官间镉含量的比值[23],如TFa/b为a器官镉含量与b器官镉含量的比值。
2 结果与分析
2.1 苹果酸对水稻幼苗根系形态和镉积累特性的影响
苹果酸能显著促进根系的生长发育并抑制镉的吸收和转运。在含有2.7 µmol·L-1镉的培养液中加入0.5~1.5 mmol·L-1的苹果酸使水稻幼苗单株平均根尖数增加43.6%~96.1%(图1A),总根长增加31.5~44.3 cm(图1B)。随着培养液中苹果酸浓度的增加,水稻根系和地上部中的镉含量显著下降。根系中的镉含量主要分布在F1 中,F3、F2 中的镉含量很低。根系F1中的镉含量对苹果酸最为敏感,添加0.5、1.0、1.5 mmol·L-1的苹果酸使水稻根系F1 中的镉含量分别下降7.9%、35.9%、45.5%,而根系F3 中镉含量仅分别下降23.8%、34.8%、39.6%(图1C)。与根系不同,地上部F1和F3中的镉含量比较接近,且F3中镉含量对苹果酸最为敏感。添加0.5、1.0、1.5 mmol·L-1的苹果酸使水稻幼苗地上部F3的镉含量分别下降29.5%、37.1%、39.7%,而F1 的镉含量仅分别下降6.7%、16.4%、13.9%(图1D)。

图1 苹果酸对水稻幼苗根尖数、根长以及镉在根系和地上部亚细胞分布特征的影响Figure 1 Effects of malic acid on root tips,root length,and the subcellular distribution of Cd in roots and shoots of rice seedlings
2.2 苹果酸对水稻幼苗氨基酸和PCs含量的影响
在培养液中添加不同浓度的苹果酸后,水稻幼苗根系和地上部中的大部分氨基酸呈现出明显的增长趋势(图2A 和图2B)。在无苹果酸的镉胁迫处理中,幼苗根系和地上部的游离氨基酸总量分别为13.1 g·kg-1和10.1 g·kg-1。添加0.5、1.0 mmol·L-1和1.5 mmol·L-1苹果酸后,幼苗根系中的氨基酸总量分别达到14.9、15.9 g·kg-1和18.2 g·kg-1,地上部的氨基酸总量分别达到11.9、13.1 g·kg-1和15.2 g·kg-1。在检测到的12 种游离氨基酸中,根系中丙氨酸、天冬氨酸、谷氨酸、半胱氨酸、甘氨酸和缬氨酸对苹果酸的浓度特别敏感,添加0.5 mmol·L-1的苹果酸即能使其含量显著增加,添加1.5 mmol·L-1苹果酸使根系中天冬氨酸、谷氨酸、半胱氨酸和甘氨酸的含量增加92.9%~140.0%,丙氨酸和缬氨酸的含量增加41.7%~54.5%(图2A)。地上部丙氨酸、天冬氨酸、谷氨酸、半胱氨酸、甘氨酸和色氨酸6 种氨基酸对苹果酸的浓度特别敏感,添加1.5 mmol·L-1苹果酸使地上部中丙氨酸、天冬氨酸、半胱氨酸和甘氨酸含量增加18.2%~66.7%,谷氨酸和色氨酸含量增加90.9%~100.0%(图2B)。添加苹果酸后,根系和地上部丙氨酸、天冬氨酸、谷氨酸、半胱氨酸和甘氨酸5 种氨基酸的含量呈显著增加的趋势。

图2 苹果酸对水稻幼苗根系和地上部氨基酸及巯基化合物的影响Figure 2 Effects of malic acid on amino acids in roots and shoots,and on sulfhydryl compounds in roots and shoots of rice seedlings
植物体内的GSH 和PCs 能通过螯合作用消除镉的生理毒性。培养液中添加苹果酸后,水稻地上部与根系中的GSH 均显著增加(图2C),其中1.5 mmol·L-1苹果酸使地上部和根系中的GSH 分别增加了68.4%和33.7%。与GSH 含量的变化趋势相反,随着培养液中苹果酸浓度的增加,地上部和根系中的PC2、PC3、PC4的含量呈显著下降的趋势(图2D)。在检测到的植物螯合肽中,PC3的含量明显高于PC2和PC4。添加1.5 mmol·L-1苹果酸使地上部和根系的PC3分别减少了84.7%和81.7%。
2.3 喷施苹果酸对水稻各器官镉含量的影响
在土壤镉含量为0.69 mg·kg-1的盆栽试验中,中早35 稻米中的镉含量高达0.56 mg·kg-1。MA1 和MA2 处理显著降低了成熟期籽粒和穗轴中的镉含量,下降幅度为37.5%~55.4%(图3A)。在苹果酸的作用下,植株顶部营养器官旗叶、穗颈和穗节中的镉含量显著下降(图3B);茎秆基部的镉含量也呈下降趋势,但未达5%显著水平。喷施苹果酸后,相邻器官间镉的TF 也显著下降(图3C)。MA1 和MA2 处理使镉的TF籽粒/穗轴从0.41 分别下降到0.38 和0.32,TF穗轴/穗颈和TF穗颈/穗节也均显著低于喷水的对照处理。穗节和茎基部压缩的分蘖节是营养器官中镉富集能力最强的器官,苹果酸对TF穗节/茎基也有显著影响,从1.37下降到1.16和1.14。

图3 苹果酸对籽粒和穗轴、营养器官镉含量以及相邻器官间镉转移因子的影响Figure 3 Effects of malic acid on Cd content in rice and rachis,vegetative organs,and Cd TF between adjacent organs of rice seedlings
2.4 喷施苹果酸对水稻天冬氨酸和谷氨酸合成的影响
在MDH 催化下,细胞中的苹果酸能高效转化成OAA,后者在AST 的催化下转化成天冬氨酸(4 碳氨基酸);AST 也能够催化α-酮戊二酸转化成谷氨酸(5碳氨基酸)(图4A)。水稻开花后,稻穗中的MDH 活性显著增加,开花后10 d 达到最大值;开花期喷施苹果酸10~30 d 内,MA1 和MA2 处理分别使MDH 的活性提高50.8%~74.5%和63.0%~96.8%(图4B);灌浆中后期(开花后20 d),MDH 活性略有下降,但依然显著高于对照处理(图4B)。稻穗中AST 的活性在开花后5 d 就达到最大值,一直保持到开花20 d,此后开始显著下降(图4C)。在开花后5~30 d 的时间内,MA1 和MA2 处理分别使AST 活性提高5.5%~12.1% 和14.6%~22.6%。
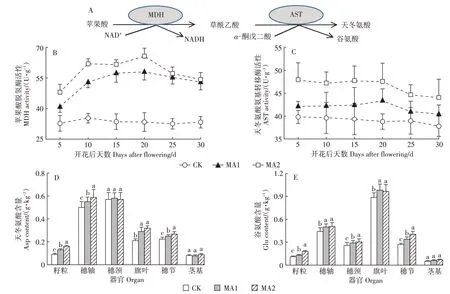
图4 苹果酸-天冬氨酸合成路线,苹果酸对MDH和AST活性以及各器官天冬氨酸和谷氨酸含量的影响Figure 4 Route of malate-aspartic acid synthesis,effects of malic acid on the activities of MDH and AST,and content of aspartic acid and glutamate in rice organs
随着酶活性的增加,籽粒和旗叶等器官中的游离天冬氨酸和谷氨酸含量也显著增加。MA1 和MA2 处理显著提高了籽粒、穗轴、旗叶和穗节中的天冬氨酸含量,对穗颈和茎基部的天冬氨酸含量无显著影响(图4D)。各器官中游离谷氨酸的含量显著高于天冬氨酸,MA1 和MA2 处理显著提高了籽粒、穗轴、旗叶、穗颈和穗节中的谷氨酸含量,仅对茎基部的谷氨酸含量无显著影响(图4E)。
3 讨论
在长期的自然进化过程中,水稻形成了适应逆境胁迫的生理调控机制。当根际环境中的镉进入水稻根系细胞后,许多镉离子被细胞壁中的大分子所固定,进入细胞膜的镉会被转运到液泡中封存起来,以减弱或消除镉对细胞质中各种正常生理活动的干扰[22,24]。本研究发现,根际环境中添加0.5~1.5 mmol·L-1苹果酸,既能显著降低水稻根系细胞壁和细胞液中的镉含量,又能显著增加根尖数目、根长和根表面积。随着根系细胞液中镉含量的显著下降,转运到地上部的镉总量大幅度降低(图1)。这是由于细胞内的小分子有机酸(如苹果酸、柠檬酸等)、氨基酸和PCs 等能与镉和铅等重金属形成螯合物,并清除细胞内过量的ROS等有害物质[9,10,13,25]。在镉胁迫环境中,外源添加苹果酸能显著提高植物生长量、增加净光合速率、减少ROS 积累、增强根系活性等,进而减轻镉的毒害作用[11-12]。不仅如此,水稻开花期叶面喷施苹果酸,也能显著降低成熟期稻米和茎叶中的镉含量(图3)。这说明提高根系和叶片中的苹果酸浓度都能有效抑制水稻细胞对镉的吸收和转运,降低地上部和稻米中的镉含量。
水稻细胞内的谷氨酸和天冬氨酸对镉离子极为敏感,籽粒和营养器官中的谷氨酸和天冬氨酸含量随着镉含量的增加而显著下降[3,5,16]。本研究发现,在水稻开花期喷施苹果酸后稻穗中MDH和AST的活性显著增加,籽粒和茎叶中游离谷氨酸和天冬氨酸含量显著上升(图4),苹果酸是三羧酸循环过程中的重要中间产物,它与OAA之间的氧化还原反应有助于维持植物细胞内NADH 和NAD+的动态平衡[9-10,15]。在细胞质中,苹果酸和α-酮戊二酸通过氨基转移作用,调控天冬氨酸和谷氨酸的合成[10,14]。这说明喷施苹果酸可激发转氨酶的活性,促进苹果酸-草酰乙酸-天冬氨酸以及α-酮戊二酸-谷氨酸的转化过程。提高根际环境中的苹果酸浓度同样能显著提高根系和地上部的天冬氨酸和谷氨酸含量(图2)。由此可见,在镉胁迫环境中提高水稻细胞的苹果酸含量,能显著促进氨基转移过程,有效消除镉对谷氨酸和天冬氨酸合成的抑制作用。
GSH是由谷氨酸、半胱氨酸和甘氨酸结合而成的三肽化合物(γ-Glu-L-Cys-L-Gly),是PCs 的前体。在植物螯合肽合成酶催化下,GSH可以进一步合成结构为[(γ-Glu-L-Cys)2~11-Gly]的PCs,PCs 能与镉形成无毒、低分子质量的Cd-S-PC 螯合物储存于液泡中[24,26-27]。水稻细胞中的镉含量与PCs 含量呈正相关关系[22,24,26]。本研究发现,提高根际环境中的苹果酸浓度后,水稻根系和地上部组织中的谷氨酸、半胱氨酸和甘氨酸含量以及GSH 含量显著增加,而PC2、PC3、PC4的含量却随着苹果酸浓度的增加而减小(图2)。这说明苹果酸通过促进谷氨酸、半胱氨酸和甘氨酸的合成提高了细胞中的GSH 含量。随着氨基酸和GSH含量的增加,细胞液中的镉含量显著下降,此时,细胞不需要合成更多的PCs 来消除镉的毒害,于是PCs的含量表现出下降趋势。
氨基酸代谢在植物的抗逆反应中发挥着重要作用。天冬氨酸和天冬酰胺、谷氨酸和谷氨酰胺是细胞中最重要的氮源供体,与酶活性和离子转运的调控有密切关系[7,28-30]。增加谷氨酸浓度能够提高阳离子通道对镉的识别和拦截,抑制有害元素从营养器官向籽粒中的转运[5,7,31]。本研究发现,喷施苹果酸后,水稻籽粒、穗轴、穗颈、穗节和旗叶中的游离谷氨酸含量显著增加(图4)。水稻开花期喷施柠檬酸、巯基丁二酸等小分子酸能够激活水稻抗氧化防御体系,提高难溶态镉在茎叶组织中的分配比例,调控水稻体内阳离子通道的活性,促进氨基酸合成和有益元素的转运,有效抑制镉从营养器官向籽粒的转运[16,23,32]。水稻开花期喷施苹果酸能够使镉从茎基向上部茎叶以及籽粒中转运的比率(即TF)大幅度下降,致使水稻各器官的镉含量自下而上呈现显著下降的趋势(图3)。由此可见,喷施苹果酸在促进谷氨酸和天冬氨酸合成的同时,显著提高了水稻营养器官对镉的拦截能力,有效抑制了水稻灌浆期间镉从旗叶向籽粒中的转运,显著降低籽粒中的镉含量。
4 结论
(1)根际环境中添加0.5~1.5 mmol·L-1的苹果酸可显著降低水稻细胞壁、细胞器及细胞液中镉的含量。与此同时,水稻地上部与根系中的谷胱甘肽及其前体谷氨酸、半胱氨酸和甘氨酸含量显著增加,植物螯合肽含量随镉含量的下降而下降。
(2)开花期喷施苹果酸显著降低成熟期籽粒和穗轴以及旗叶、穗颈和穗节中的镉含量,使镉从穗轴向籽粒转运的比例从0.41 下降到0.32~0.38,相邻营养器官间镉的转移因子也显著下降。
(3)喷施苹果酸显著提高灌浆期稻穗中苹果酸脱氢酶和天冬氨酸氨基转移酶的活性,使穗轴、穗茎和旗叶中谷氨酸含量显著上升,从而抑制了镉向顶部营养器官和籽粒中的转运。
猜你喜欢
山东化工(2017年22期)2017-12-20
三门峡职业技术学院学报(2017年1期)2017-06-05
中国洗涤用品工业(2017年2期)2017-04-16
化工设计通讯(2017年10期)2017-03-02
中国比较医学杂志(2017年5期)2017-01-17
分析仪器(2016年2期)2016-09-07
听力学及言语疾病杂志(2015年5期)2015-12-24
医学研究杂志(2015年12期)2015-06-10
中国酿造(2013年5期)2013-01-26
中国果业信息(2013年12期)2013-01-22