
酵母多糖对育肥牛生长性能、血清免疫和抗氧化指标以及代谢物的影响
2023-11-10 06:09陈志龙孙文阳梁小军侯鹏霞张恩平
动物营养学报 2023年10期
王 燕 陈志龙 施 安 李 博 孙文阳 李 丹 梁小军 侯鹏霞* 张恩平*
(1.西北农林科技大学动物科技学院,杨凌 712100;2.宁夏农林科学院动物科学研究所,银川 750000;3.宁夏农林科学院固原分院,固原 756000)
酵母多糖是从酵母细胞壁中提取出来的功能性复合多糖,β-葡聚糖和甘露聚糖是其主要的活性物质[1],二者对机体的生命活动具有重要调节作用。研究表明,β-葡聚糖具有良好的免疫调节活性,一方面通过提高免疫球蛋白和白细胞介素水平,增强宿主对细菌、病毒、真菌及寄生虫感染的非特异性免疫和抗氧化能力[2-4];另一方面通过促进抗体的产生,增强机体的特异性免疫功能[5]。甘露聚糖的结构类似病原菌定植在肠道上的特异性受体,可作为高亲和性配体与革兰氏阴性菌的结合位点结合,致使病原菌无法在肠道内附着或定植,从而被清除[6];还可作为免疫刺激辅因子,与细菌、病毒和毒素等外源抗原结合以增加其被吸收的时间,调节机体特异性免疫[2]。酵母多糖则结合了β-葡聚糖和甘露聚糖2大成分的主要优点,具有促进生长、调节免疫和提高抗氧化性能等生物学功能。董金金[7]研究报道,饲粮添加2 g/(头·d)酵母多糖能促进哺乳犊牛胃肠道形态发育,提高胃肠道消化酶活性,促进犊牛生长发育和营养物质消化吸收;Ma等[8]研究报道,饲粮添加适量酵母β-葡聚糖显著提高了荷斯坦犊牛的饲料转化率和日增重;Rozeboom等[9]研究发现,饲粮添加甘露寡糖提高了免疫球蛋白水平,增强了巨噬细胞活性,提升了仔猪免疫力。
目前,酵母多糖在反刍动物生产中的研究相对较少,已有研究报道也主要集中在酵母多糖对哺乳期犊牛胃肠道发育的影响上,关于酵母多糖对育肥牛在生产上的影响鲜有报道。因此,本研究以酵母多糖(富含β-葡聚糖和甘露聚糖)为试验材料,以育肥牛为研究对象,旨在探究饲粮添加酵母多糖对育肥牛生长性能、血清免疫和抗氧化指标以及代谢物的影响,为酵母多糖在肉牛育肥中的应用提供理论依据。
1 材料与方法
1.1 试验设计
本试验于2022年5月至8月在宁夏回族自治区固原市原州区某肉牛养殖场进行。按照单因素随机试验设计,选择健康的16月龄、体重550 kg左右的西门塔尔杂交育肥牛40头,随机分为4组,每组10头。对照组饲喂基础饲粮,试验组在晨饲时在基础饲粮中分别添加5、10和15 g/(d·头)酵母多糖(购自某公司,主要成分及含量:甘露聚糖≥20%,β-葡聚糖≥20%,粗蛋白质≥25%,水分≤6%),并分别记为试验1组、试验2组和试验3组。基础饲粮根据NRC(2016)肉牛饲养标准结合养殖场生产实际配制,其组成及营养水平见表1。预试期10 d,正试期94 d(试验前期50 d,试验后期44 d)。
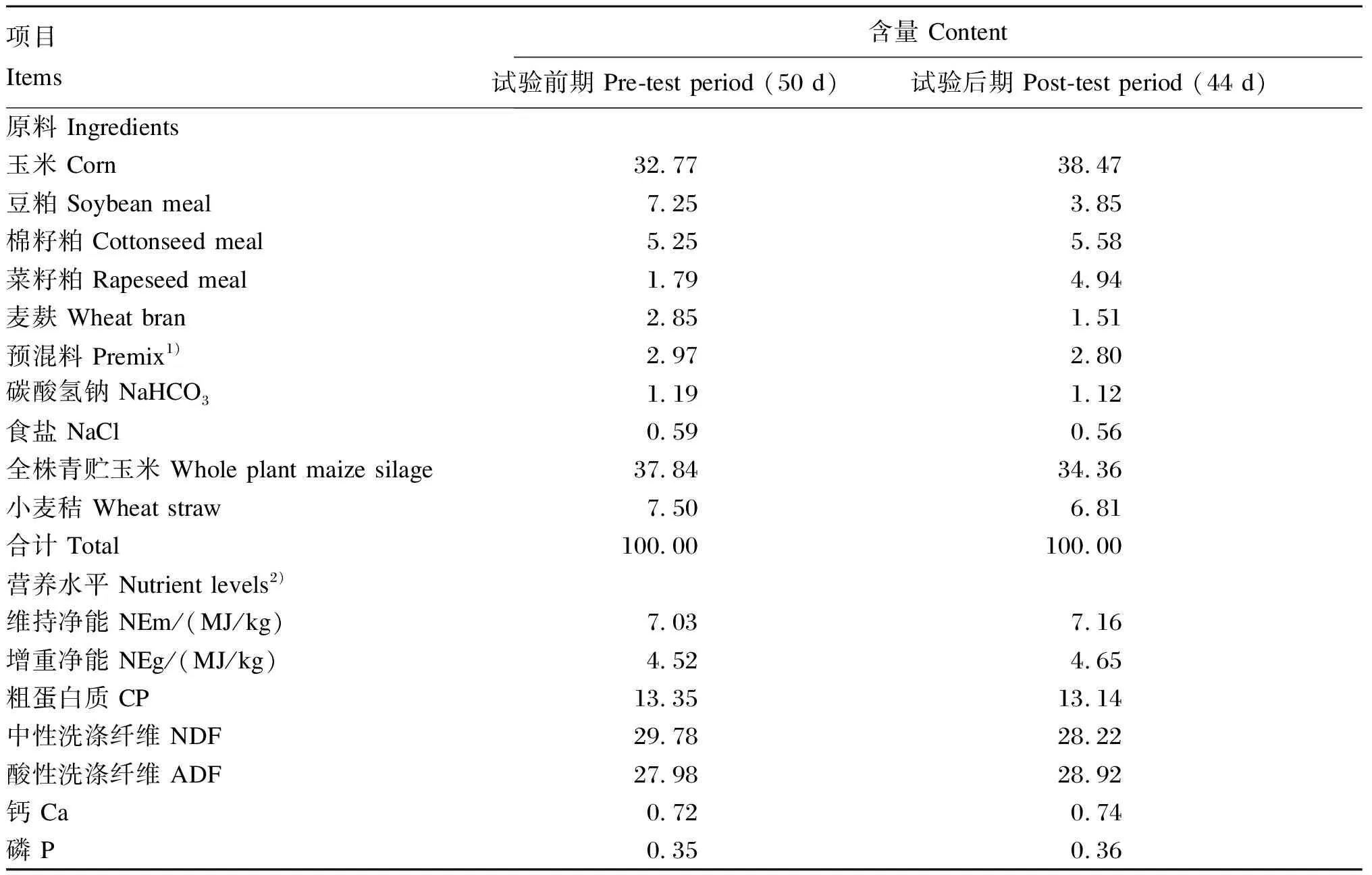
表1 基础饲粮组成及营养水平(干物质基础)
1.2 饲养管理
试验开始前对圈舍进行打扫、消毒。试验牛分圈舍群饲,各组按试验设计饲喂对应饲粮,基础饲粮以全混合日粮(TMR)形式饲喂,每日2次(08:00和16:00),自由饮水。试验组于每日晨饲时,将酵母多糖用水溶解喷洒于饲粮表面(确保育肥牛短时间内采食完)。其他饲养条件保持一致。
1.3 测定指标及方法
1.3.1 生长性能
正试期开始当天、第50天和结束当天(试验第94天)晨饲前,使用地磅对试验牛称重。各组间饲粮添加量保持一致,根据剩料量及时调整添加量,随着体重增加添加量增大(试验前期12.70 kg/d,试验后期14.20 kg/d),计算平均日增重(ADG)和料重比(F/G)。
1.3.2 经济效益
饲粮和添加剂成本均按照试验时该地区的市场价格进行计算。根据育肥牛出栏时的活牛市场价格计算每头牛的增重收益,最后根据每头牛的成本及增重收益计算经济效益。计算公式如下:
利润=增重收益-成本。
1.3.3 血清免疫和抗氧化指标及代谢组学
正试期结束当天晨饲前,试验牛尾静脉采血10 mL,静置30 min,3 000 r/min离心15 min,分离血清。将血清分装2份于1.5 mL离心管中,一份于-20 ℃保存,用于血清免疫和抗氧化指标测定;另一份立即投入液氮,带回实验室于-80 ℃保存,用于代谢组学测定。
血清免疫指标:使用Rayto RT-6100酶标分析仪(美国雷杜)并采用酶联免疫吸附试验(ELISA)法测定血清免疫球蛋白A(IgA)、免疫球蛋白G(IgG)、免疫球蛋白M(IgM)、干扰素-γ(IFN-γ)和肿瘤坏死因子-α(TNF-α)含量,试剂盒购自上海科淘生物科技中心。
血清抗氧化指标:使用UNICO-U7200分光光度计[尤尼柯(上海)仪器有限公司]并采用比色法测定血清超氧化物歧化酶(SOD)、过氧化氢酶(CAT)、谷胱甘肽过氧化物酶(GSH-Px)活性以及丙二醛(MDA)含量,试剂盒购自南京建成生物工程研究所。
血清代谢组学:基于气相色谱-质谱联用(GC-MS)技术进行非靶向代谢组学研究,试验流程主要包括样品收集、代谢物提取、GC-MS检测及数据分析等,以上检测委托北京诺禾致源科技股份有限公司完成。
1.4 数据处理
试验数据先用Excel 2019初步整理,然后采用SPSS 26.0进行单因素方差分析(one-way ANOVA),并采用LSD法进行多重比较,结果用“平均值±标准误”表示,P<0.05为差异显著。
2 结果与分析
2.1 酵母多糖对育肥牛生长性能的影响
由表2可知,各组之间育肥牛初始体重和第50天体重均无显著差异(P>0.05);试验2组和试验3组终末体重显著高于对照组(P<0.05);试验组后44 d ADG显著高于对照组(P<0.05);试验3组全期ADG显著高于其他组(P<0.05),且F/G最低。
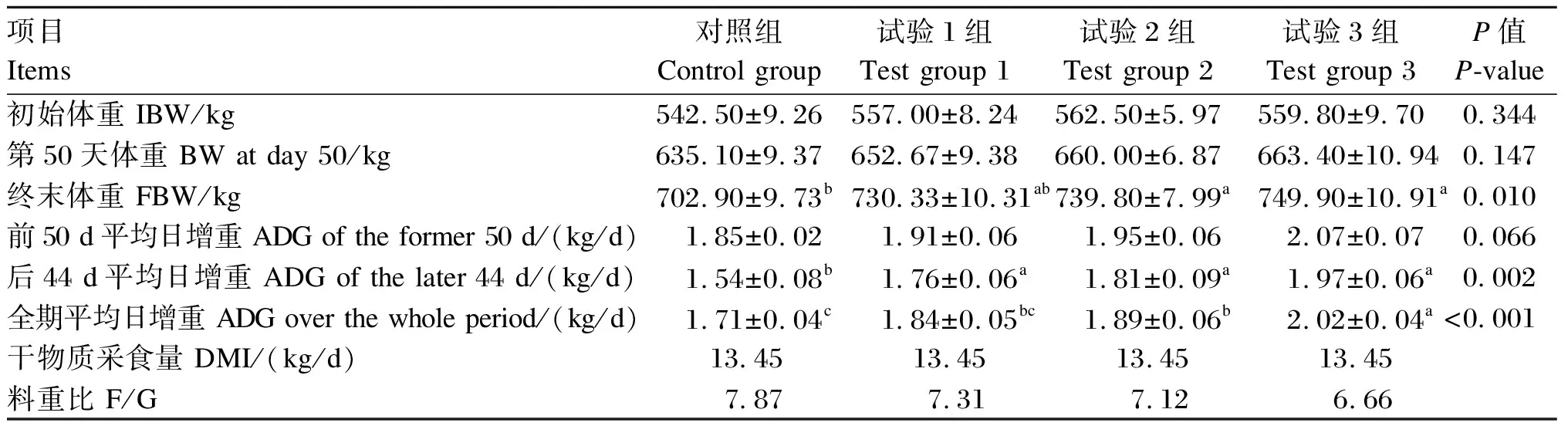
表2 酵母多糖对育肥牛生长性能的影响
2.2 酵母多糖对育肥牛经济效益的影响
由表3可知,对照组、试验1组、试验2组和试验3组育肥牛ADG分别为1.71、1.84、1.89和2.02 kg/d,增重收益分别为5 786.64、6 226.56、6 395.76和6 835.68元/头,总成本分别为3 865.28、3 893.48、3 921.68和3 949.88元/头,其中试验1组、试验2组和试验3组每日每头育肥牛饲料添加剂成本较对照组分别增加了0.3、0.6和0.9元;试验组利润较对照组分别提高了411.72、552.72和964.44元/头。
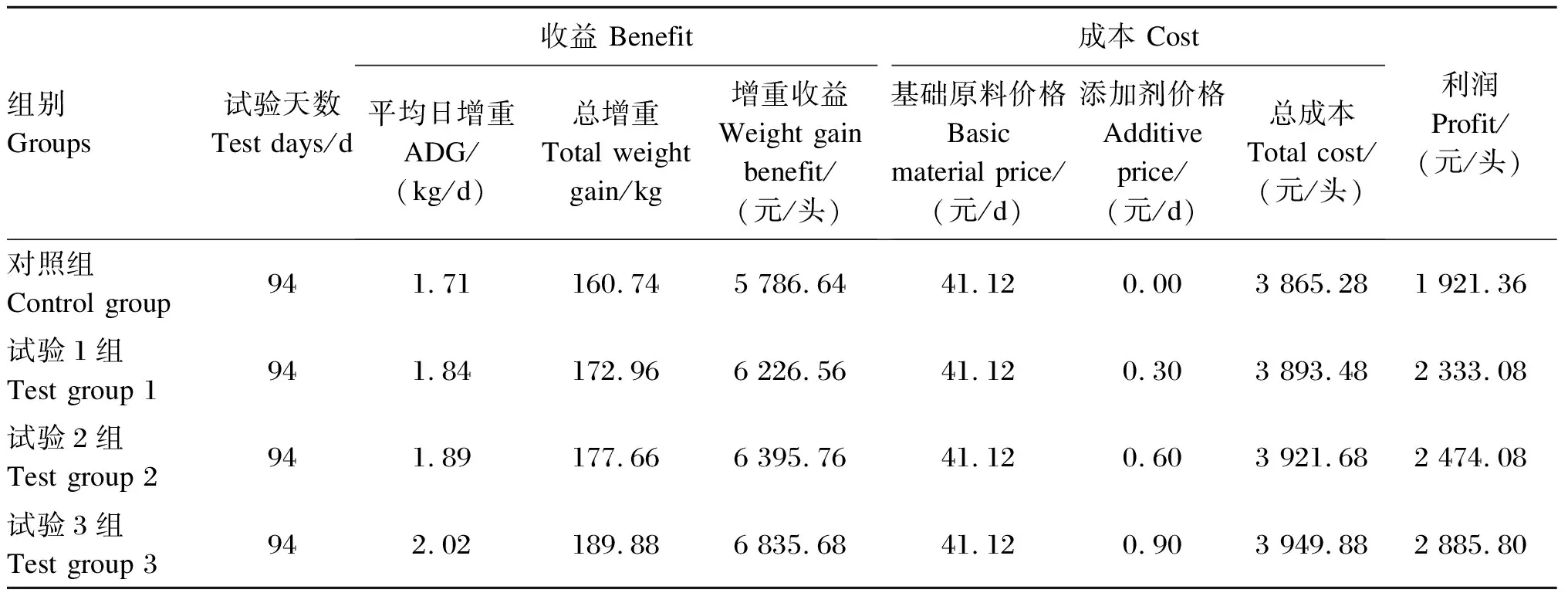
表3 酵母多糖对育肥牛养殖经济效益的影响
2.3 酵母多糖对育肥牛血清免疫指标的影响
由表4可知,各组之间育肥牛血清IgA和IgM含量无显著差异(P>0.05);试验3组血清IgG含量显著高于试验1组和试验2组(P<0.05),血清IFN-γ含量显著高于其他组(P<0.05);各组之间血清TNF-α含量无显著差异(P>0.05),但以试验2组最低。

表4 酵母多糖对育肥牛血清免疫指标的影响
2.4 酵母多糖对育肥牛血清抗氧化指标的影响
由表5可知,试验组育肥牛血清GSH-Px活性高于对照组,其中试验1组显著高于对照组(P<0.05);血清MDA含量随着饲粮中酵母多糖添加水平的提高呈降低趋势,试验2组和试验3组较对照组和试验1组显著降低(P<0.05);试验2组血清SOD活性显著高于其他组(P<0.05);试验3组和试验2组血清CAT活性差异不显著(P>0.05),但试验3组显著高于对照组和试验1组(P<0.05)。
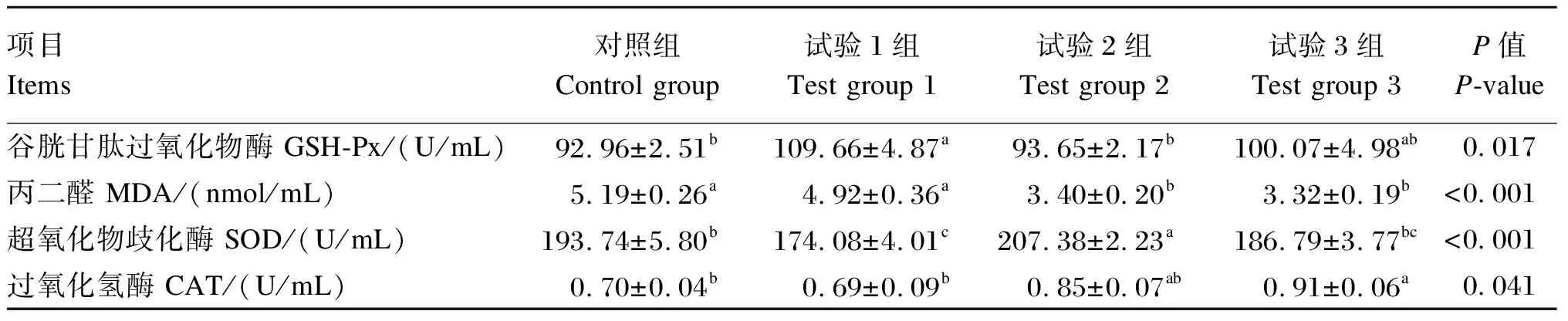
表5 酵母多糖对育肥牛血清抗氧化指标的影响
2.5 酵母多糖对育肥牛血清代谢物的影响
2.5.1 样本质量控制(QC)
基于峰面积值来计算QC样本间的Pearson相关系数。如图1所示,QC样本的相关系数均大于0.98,说明整个检测过程机器稳定性好,数据质量较高。
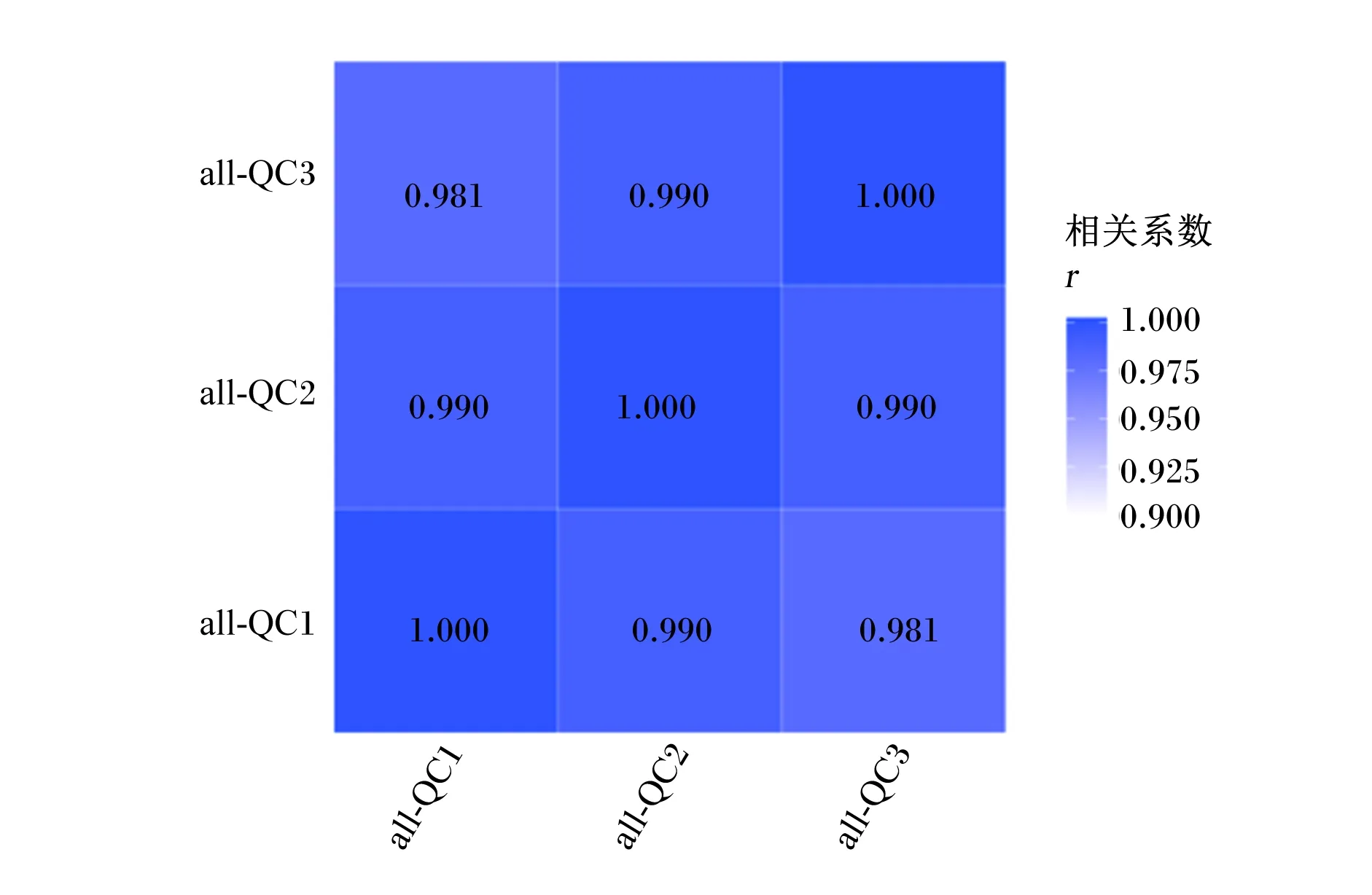
all-QC:所有质量控制样本 all QC samples。
2.5.2 主成分分析(PCA)和偏最小二乘回归分析(PLS-DA)
如图2 所示,从对照组和试验组的血清样本散点图可以看出其主成分1(PC1)、主成分2(PC2)和主成分3(PC3)分别为26.92%、12.41%和9.26%。试验组与对照组样本点较为分散,各组代谢物轮廓并不完全重叠,但由于存在重叠,需进一步采用有监督模式识别方法PLS-DA进行判别分析。
为进一步阐释各组血清代谢物之间的差异,对样品进行PLS-DA。如图3 所示,对照组与试验组两两区分明显,且同组样品在组内聚集良好,各组血清样品的组内差异小于组间差异,说明试验组与对照组血清代谢物存在差异。各组的PLS-DA图中R2Y值均接近于1,表明该模型的解释率高,且R2Y均大于Q2Y,表明该模型建立良好。进一步对PLS-DA模型进行置换检验,如图4 所示,随着置换保留度的下降,R2和Q2下降,回归线呈上升的趋势,且回归线的Q2在纵坐标上的交叉点均小于0,说明置换检验过关,模型不存在过拟合现象。
2.5.3 差异代谢物的筛选
以变量投影重要性(VIP)>1.0、差异倍数(FC)>1.2或FC<0.833且P<0.05作为筛选差异代谢物的条件,分析结果见表6。与对照组相比,试验组育肥牛血清共筛选出15种差异代谢物,其中试验1组、试验2组和试验3组较对照组分别显著上调1、5和1种差异代谢物(P<0.05),分别显著下调3、5和7种差异代谢物(P<0.05);显著上调的差异代谢物有硫辛酰胺、1,3-丙二胺、α-酮异己酸、β-氨基异丁酸和3-甲胺基-1,2-丙二醇,显著下调的差异代谢物有乳糖、蔗糖、麦芽糖、茉莉酸甲酯、谷氨酰胺、阿特拉津-2-羟基、马来酰亚胺、邻苯二甲酸、十二醇和抗坏血酸。用火山图将筛选的差异代谢物进行可视化演示,结果如图5 所示。与对照组相比,试验组共有的差异代谢物为乳糖,显著下调(P<0.05)。采用MetaboAnalyst 5.0中经典单变量受试者工作特征(ROC)曲线对乳糖进行生物标志物分析,结果发现乳糖在试验组与对照组的比较中曲线下面积(AUC)值均为1,大于0.9,说明乳糖的预测准确性较高,可视为酵母多糖参与调控育肥牛代谢的潜在生物标志物。
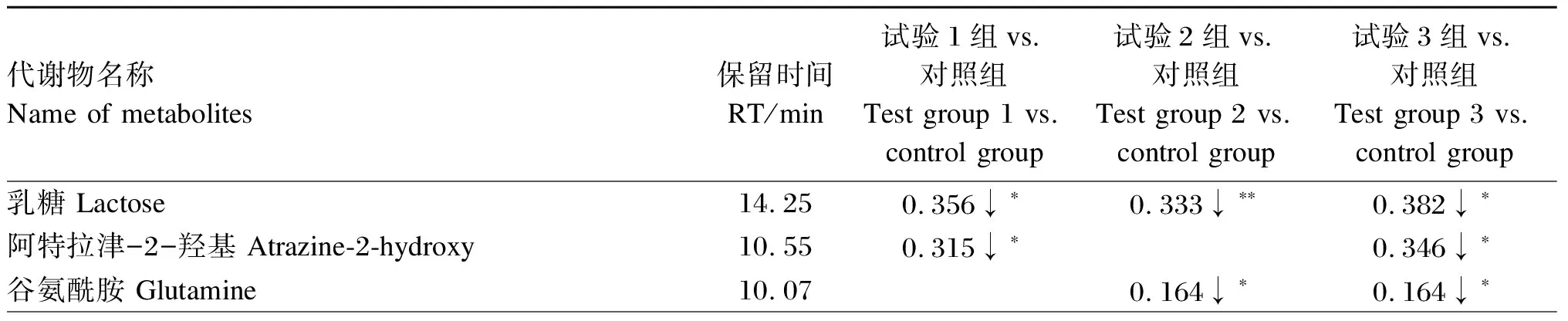
表6 酵母多糖对育肥牛血清代谢物的影响
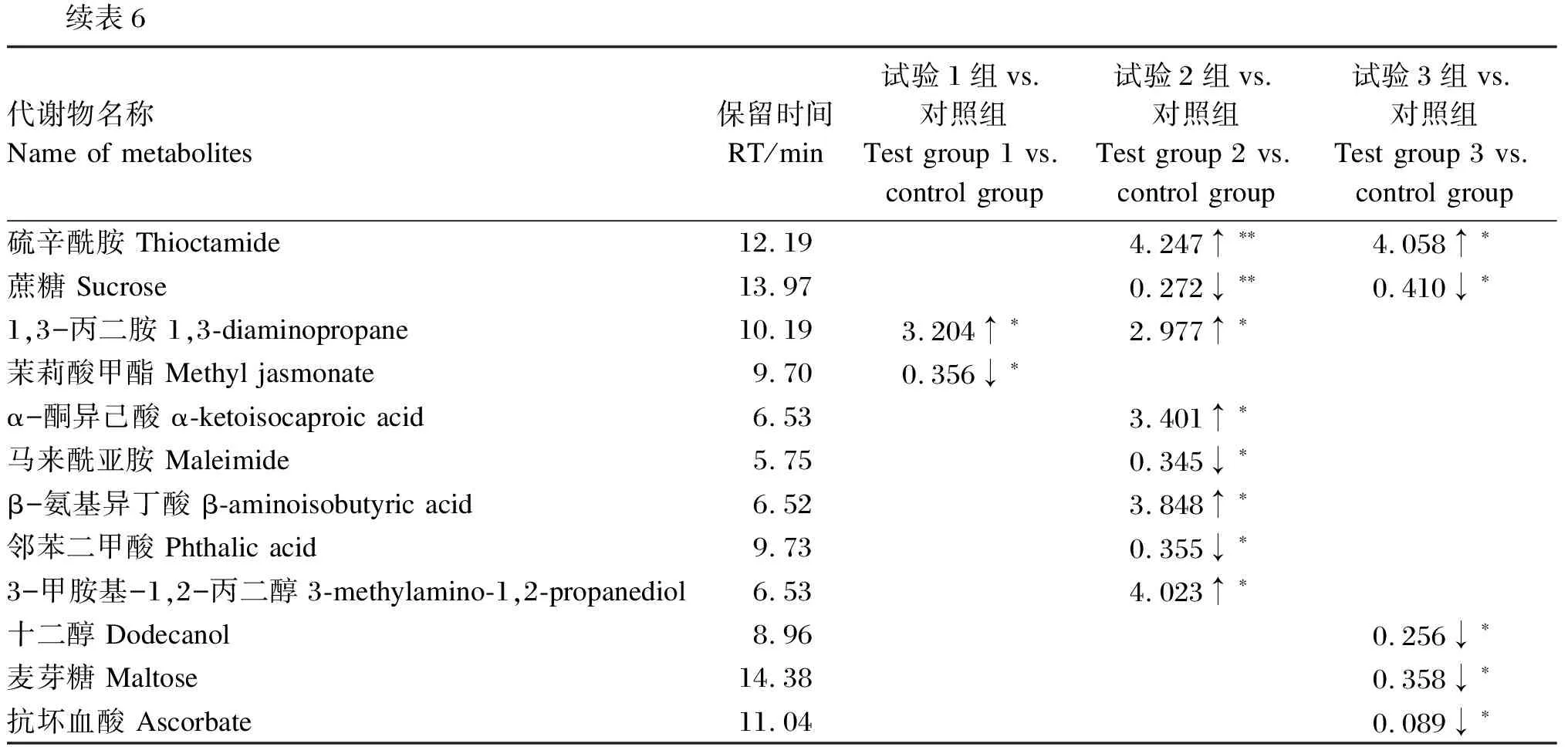
续表6代谢物名称Name of metabolites保留时间RT/min试验1组vs.对照组Test group 1 vs. control group试验2组vs.对照组Test group 2 vs. control group试验3组vs.对照组Test group 3 vs. control group硫辛酰胺 Thioctamide12.194.247↑∗∗4.058↑∗蔗糖 Sucrose13.970.272↓∗∗0.410↓∗1,3-丙二胺 1,3-diaminopropane10.193.204↑∗2.977↑∗茉莉酸甲酯 Methyl jasmonate9.700.356↓∗α-酮异己酸 α-ketoisocaproic acid6.533.401↑∗马来酰亚胺 Maleimide5.750.345↓∗β-氨基异丁酸 β-aminoisobutyric acid6.523.848↑∗邻苯二甲酸 Phthalic acid9.730.355↓∗3-甲胺基-1,2-丙二醇 3-methylamino-1,2-propanediol6.534.023↑∗十二醇 Dodecanol8.960.256↓∗麦芽糖 Maltose14.380.358↓∗抗坏血酸 Ascorbate11.040.089↓∗

图5 血清代谢物火山图
2.5.4 差异代谢物富集通路
采用京都基因与基因组百科全书(KEGG)数据库注释试验组与对照组差异代谢物显著富集的代谢通路,结果如图6 所示。与对照组相比,试验2组差异代谢物显著富集通路有半乳糖代谢(galactose metabolism)、味觉传导(taste transduction)以及碳水化合物消化与吸收(carbohydrate digestion and absorption)(P<0.05)(图6-a);试验3组差异代谢物极显著富集通路有味觉传导、碳水化合物消化与吸收以及淀粉和蔗糖代谢(starch and sucrose metabolism)(P<0.01)(图6-b)。
3 讨 论
3.1 酵母多糖对育肥牛生长性能的影响
生长速度是反映家畜生长性能和饲料利用率的重要指标[10]。酵母多糖通过促进新陈代谢、提高免疫力提高了动物机体对营养物质的消化吸收,进而促进生长[11-14]。刘佳等[1]研究报道,饲粮添加0.1%酵母多糖可增强断奶仔猪免疫力和抗氧化功能,改善肠道健康,有效促进生长;许飞龙等[15]、吴宇梁等[16]研究报道,饲粮添加0.1%酵母多糖显著提高了断奶仔猪ADG,显著降低了F/G;王辉田等[17]研究发现,饲粮添加2 000 mg/kg酵母细胞壁多糖提高了22~43日龄肉仔鸡ADG,降低了F/G;董金金[7]研究报道,饲粮添加2 g/(头·d)酵母多糖显著提高了犊牛ADG和干物质采食量。以上研究结果均说明,酵母多糖可不同程度地促进家畜生长。但徐杰等[18]研究报道,饲粮添加酵母细胞壁多糖没有显著促进断奶仔猪的生长,这可能与酵母多糖的加工方式、使用剂量、浓度以及活化程度等有关。本研究中,在初始体重相近的情况下,饲粮添加10和15 g/(d·头)酵母多糖显著提高了育肥牛终末体重,且15 g/(d·头)酵母多糖添加组ADG显著高于其他组,F/G最低,这与刘佳等[1]、许飞龙等[15]的研究结果一致。
已有研究表明,饲粮添加酵母培养物等微生态制剂后,育肥牛ADG、营养物质表观消化率等均得到一定程度的改善,养殖场经济效益得到显著提高[19-20]。本试验中,与对照组相比,饲粮添加5、10和15 g/(d·头)酵母多糖使得育肥牛ADG分别提高了0.13、0.18和0.31 kg/d,利润分别提高了411.72、552.72和964.44元/头,与上述研究结论一致,说明饲粮添加酵母多糖有利于促进育肥牛生长,提高养殖场经济效益,在生产中具有一定的推广应用前景。
3.2 酵母多糖对育肥牛血清免疫指标的影响
免疫系统是保障动物机体健康的重要屏障。酵母多糖具有提高免疫器官指数[21]、T细胞数量[22]、免疫球蛋白及免疫因子水平[18]的能力,可直接激活免疫系统,增强动物机体的先天性和适应性免疫应答[23-24]。IgA、IgG和IgM是体液免疫系统重要成员,其水平上升预示着免疫系统启动,IgG约占血清免疫球蛋白的75%,具有调节机体免疫、抗感染及促进代谢等功能[25]。马吉锋等[10]研究发现,饲粮添加0.1%、0.2%和0.3%酵母细胞壁多糖显著提高滩羊血清IgG、IgA和IgM含量,且添加量为0.2%时,血清IgG、IgA和IgM含量均达到最高;邵亚群等[26]研究表明,饲粮添加酵母细胞壁多糖能显著提高犊牛血清IgG、IgA、白细胞介素-4(IL-4)和TNF-α含量,增强犊牛免疫力;李学俭[27]研究报道,β-甘露聚糖酶显著提高了断奶仔猪血清IgG和IgA含量。本研究中,各组间育肥牛血清IgA和IgM含量无显著差异,15 g/(d·头)酵母多糖添加组育肥牛血清IgG含量显著高于5和10 g/(d·头)酵母多糖添加组,但与对照组无显著差异,这与上述研究结果不一致,说明添加酵母多糖会对育肥牛免疫性能起到一定调控作用,但具体机理有待进一步研究。IFN-γ通过增强巨噬细胞、T细胞和自然杀伤细胞的活力,调节机体免疫。Giang等[28]研究表明,饲喂酵母培养物显著提高了绵羊血清白细胞介素-2(IL-2)和IFN-γ含量。本试验中,饲粮添加15 g/(d·头)酵母多糖显著提高了育肥牛血清IFN-γ含量,与上述研究结果一致。TNF-α是一种多向性促炎因子(由脂肪细胞、活化的单核细胞、巨噬细胞、B细胞、T细胞和成纤维细胞分泌),是转导细菌感染免疫应答的一个重要因子。罗作明[29]研究报道,饲粮添加0.15%~0.20%酵母多糖可降低仔猪血清促炎因子TNF-α和白细胞介素-6(IL-6)含量,提高血清抗炎因子白细胞介素-10(IL-10)含量。本研究中,10 g/(d·头)酵母多糖添加组育肥牛血清TNF-α含量最低,与罗作明[29]研究结果一致,这可能是因为酵母多糖抑制了沙门氏菌、大肠杆菌等有害细菌的生长,从而使机体减少了TNF-α的合成与释放。
3.3 酵母多糖对育肥牛血清抗氧化指标的影响
畜禽新陈代谢、氧化呼吸供能会产生化学性质极不稳定的自由基,畜禽体内自由基与抗氧化酶之间不平衡,会引起氧化应激,导致细胞组织结构受损。SOD和GSH-Px是氧自由基的清除剂,能有效缓解机体氧化应激,在维持细胞的氧化与抗氧化平衡中充当重要角色[30]。MDA是机体脂质过氧化的最终产物(具有细胞毒性),其含量高低可反映氧自由基水平和脂质过氧化的速率和强度,当MDA含量升高时,代表着机体的抗氧化性能减弱。酵母多糖可有效清除羟自由基和超氧阴离子自由基,抑制脂质氧化产物MDA的生成,提高与抗氧化功能相关酶的活性,增强机体的抗氧化能力[31]。Bozkurt等[32]研究发现,饲粮添加甘露寡糖可提高蛋鸡肝脏SOD活性并降低蛋黄MDA含量;Wang等[33]研究发现,饲粮添加酵母细胞壁可提高肉鸡21和42日龄空肠SOD活性和42日龄空肠GSH-Px活性;顾鲲涛等[34]研究发现,饲粮添加10 g/(头·d)酵母β-葡聚糖能显著提高围产奶牛产前和产后血清GSH-Px活性,有降低产前血清MDA含量的趋势;李冬芳等[35]研究发现,饲粮添加β-葡聚糖能提高肉羊血清SOD活性,并降低血清MDA含量;Zhang等[36]研究表明,在母猪妊娠和哺乳期间补充酵母培养物可提高血清和母乳SOD活性,降低血清MDA含量。
本试验中,饲粮添加适宜水平酵母多糖能够提高育肥牛血清GSH-Px和SOD活性,降低血清MDA含量,与上述研究结果一致。其中,酵母多糖添加水平为10 g/(d·头)时显著提高了血清SOD活性,添加水平为10和15 g/(d·头)时显著降低了血清MDA含量。这可能是因为酵母多糖的免疫刺激活性和对病原体的有效清除减少了自由基的产生[37],并因此减少了SOD和GSH-Px的合成。CAT具有清除氧自由基,抑制脂质过氧化,从而减少氧化损伤的功能[38]。Czech等[39]研究发现,饲粮添加酵母甘露寡糖可显著提高后代仔猪血清SOD和CAT活性。本试验中,饲粮添加15 g/(d·头)酵母多糖显著提高了育肥牛血清CAT活性,与上述研究结果一致。这可能是因为外源添加酵母多糖提高了机体内清除氧自由基的抗氧化酶的活性,减少了活性氧对机体造成的损伤,保障了机体健康,具体作用机理有待进一步研究。
3.4 酵母多糖对育肥牛血清代谢物的影响
代谢组学是对代谢机制的研究,是通过测量整个生物样本中的代谢产物并阐明其潜在的代谢通路来实现的[40]。代谢物可通过各种新陈代谢反应合成能量及生物大分子物质,直接满足个体生长发育的物质需求,且不同的代谢物及代谢通路会影响相关组织器官的发育[41]。本试验中,通过代谢组学技术对育肥牛血清中小分子代谢物进行定性和定量分析发现,与对照组相比,试验组育肥牛血清中共筛选出15种差异代谢物,其中乳糖是试验组较对照组共有的差异代谢物且含量显著下调;此外,10 g/(d·头)酵母多糖添加组差异代谢物显著富集通路有半乳糖代谢、味觉传导以及碳水化合物消化与吸收,15 g/(d·头)酵母多糖添加组差异代谢物极显著富集通路有味觉传导、碳水化合物消化与吸收以及淀粉和蔗糖代谢。
碳水化合物在机体生长发育和维系健康等方面发挥重要作用[42],是食物中的重要组成部分,是大多数生命机体重要能量来源,碳水化合物代谢在能量代谢上发挥重要作用。一般情况下,人类饮食中碳水化合物含量占比50%以上,而作为畜禽饲粮主要原料的玉米中碳水化合物含量超过70%[43],并且随着育肥的进展,碳水化合物代谢不断提高。碳水化合物主要分为单糖、双糖、寡糖及多糖,其中双糖主要包括乳糖、蔗糖和麦芽糖等。
乳糖主要在空肠、回肠中被消化吸收,通过小肠上皮细胞刷状缘分泌的乳糖酶将其水解为葡萄糖和半乳糖后,依靠细胞的主动转运被吸收。乳糖是分解速度极快的可溶性糖,易被消化吸收分解为单糖并产生大量的能量[44];还可以选择性地刺激嗜酸细菌生长,增加乳酸菌数量,从而调节微生物组成,使动物机体表现出更大的增重和更好的饲料效率[45]。生产中,乳糖常用于过渡仔猪断奶应激,改善肠道健康,促进仔猪采食[46];周旭森[44]研究报道,饲粮添加1%乳糖可显著提高母羊ADG和净增重,提高生长性能。本研究中,饲粮添加酵母多糖显著下调了育肥牛血清乳糖含量,这可能是因为在育肥阶段高精料饲粮的饲喂下,酵母多糖会促进大部分乳糖分解生成葡萄糖,且机体多余的葡萄糖会部分转化为非糖物质,如脂肪,从而促进育肥牛脂肪的沉积,提高生长性能。
蔗糖在动物体内被水解成果糖和葡萄糖[47]。麦芽糖是由2个葡萄糖单糖经由α-1,4糖苷键连接而成的二糖。本试验中,饲粮添加酵母多糖显著下调育肥牛血清中蔗糖和麦芽糖含量。从KEGG通路富集分析可知,蔗糖和麦芽糖主要参与味觉传导、碳水化合物消化与吸收及淀粉和蔗糖代谢通路。由此可推测,酵母多糖促进了育肥牛对碳水化合物的吸收和利用,有效防止了高精料饲粮饲喂可能会导致的亚急性瘤胃酸中毒、瘤胃酸中毒等营养代谢性疾病发生,且酵母多糖添加水平为10~15 g/(d·头)可促进育肥牛健康生长,其具体作用机制有待进一步探究。
4 结 论
本研究条件下,饲粮添加10~15 g/(d·头)酵母多糖提高了育肥牛免疫和抗氧化性能,其中以添加15 g/(d·头)时育肥牛增重最快,经济效益最高。饲粮添加酵母多糖后,育肥牛血清筛选出15种差异代谢物,其中乳糖可能是酵母多糖参与调控育肥牛代谢的潜在生物标志物;差异代谢物主要参与味觉传导、碳水化合物消化与吸收、淀粉和蔗糖代谢以及半乳糖代谢通路。
猜你喜欢
现代临床医学(2022年4期)2022-09-29
动物营养学报(2022年8期)2022-09-01
动物营养学报(2017年8期)2017-08-16
中国调味品(2017年2期)2017-03-20
创新作文(小学版)(2016年16期)2016-11-11
现代检验医学杂志(2016年5期)2016-08-20
分析测试学报(2015年7期)2016-01-13
动物营养学报(2015年9期)2016-01-07
中国科技信息(2015年2期)2015-11-16
质谱学报(2015年5期)2015-03-01
