
湖南阳明山山顶矮林群落结构及其与土壤因子的相关性
2023-12-14 05:28陈艳华张文静彭重华龙岳林
安徽农业科学 2023年23期
关键词:群落结构
陈艳华 张文静 彭重华 龙岳林

摘要 [目的]了解湖南阳明山山顶矮林的群落结构及土壤环境状况。[方法]对湖南阳明山的山顶矮林进行野外调查,并对生境土壤进行测定。[结果]阳明山山顶矮林样地内共记录到37科51属68种植物,杜鹃花科杜鹃花属植物是乔木层和灌木层的优势植物,而乔木层的云锦杜鹃(Rhododendron fortunei)在数量、高度及重要值上占据绝对优势地位,呈单优种群落结构特征。乔木层、灌木层和草本层各科重要值与土壤有机质(OM)、全氮(TN)具有显著相关性(P<0.05)。[结论]湖南阳明山山顶矮林是亚热带山地常绿阔叶林在山顶特殊生境下长期发育形成的特殊群落变型,具有单优种群落的结构特征;其中,土壤因子对亚热带山顶植被的发育具有重要意义。
关键词 山顶矮林;群落结构;土壤因子;湖南阳明山
中图分类号 S 718.5 文献标识码 A 文章编号 0517-6611(2023)23-0104-06
doi:10.3969/j.issn.0517-6611.2023.23.024
Community Structure of the Dwarf Forest and Relation with Soil Factors at Ridge of the Yangming Mountain in Hunan Province,China
CHEN Yan-hua1,ZHANG Wen-jing1,PENG Zhong-hua2 et al
(1.Hunan Institute of Science and Technology,Yueyang,Hunan 414006;2.College of Landscape Architecture,Central South University of Forestry and Technology,Changsha,Hunan 410004)
Abstract [Objective]To understand the community structure and soil environment status of the mountain top dwarf forest in Yangming Mountain,Hunan.[Method]A field investigation was conducted on the mountaintop dwarf forest in Yangming Mountain,Hunan Province,and the soil of the habitat was measured.[Result]The results showed that a total of 37 families,51 genera,and 68 species of plants were recorded in the low forest sample plot on the mountaintop of Yangming Mountain.Among them,Rhododendron are the dominant genera in the tree layer and shrub layer.Rhododendron fortunei occupies an absolute dominant position in the number,height and importance value,and showing single-dominant community structure.The important values of tree layer,shrub layer and herb layer were significantly correlated with OM and TN(P<0.05).[Conclusion]The mountaintop dwarf forest in Yangming Mountain,Hunan Province is a special community variant formed by the long-term development of subtropical evergreen broad-leaved forest in a special mountaintop habitat,with structural characteristics of single superior species communities;Among them,soil factors are of great significance for the development of subtropical mountaintop vegetation.
Key words The mountaintop dwarf forest;Community structure;Soil factors;Yangming Mountain in Hunan
基金項目 湖南省社会科学成果评审委员会课题(XSP18YBC018)。
作者简介 陈艳华(1971— ),女,湖南永州人,副教授,博士,从事植物生态景观研究与设计。
收稿日期 2022-12-08
山顶矮林是亚热带山地常绿阔叶林在山顶或山脊、云雾线以上,生长在湿度大、山风强、气温低且昼夜温差大的生境下长期发育形成的特殊群落变型[1],具有敏感性、脆弱性、不可替代性等特点[2],如遭破坏轻则发展成以原有植物为主的萌生灌丛,重则成为莎草科、禾本科、灯芯草科、蕨类等为主的湿性草地,更严重的会难以成林甚至消失。
土壤是植物生长的基质,为植物根系生长提供场所及必需的矿质营养和水分[3],影响植物群落组成、结构及多样性,甚至影响植被的更新和演替[4]。目前,关于山顶矮林土壤等环境因子的研究集中于热带地区,如热带高山雨林地区山顶矮林土壤的粗质腐殖质层深厚[5-7]、半分解有机物多,N、P含量少[5-6]、C/N升高[5];降水、淋溶、内涝、灰化作用及树叶类型导致土壤pH小而呈酸性[8];我国云南南部6个国家级自然保护区的山顶矮林土壤TN、TP、TK含量较高,土壤肥力较海南岛热带山顶矮林高[8-9],且土壤磷元素可能是影响热带山顶矮林植被分布的重要因子[9]。而亚热带地区山顶矮林土壤等环境因子的研究则较为缺乏。笔者研究了亚热带山顶矮林群落结构及其与土壤因子的关系,对深入了解亚热带山顶矮林及其特定环境因子,维护亚热带山顶植被生态系统及其环境营建具有重要意义。
1 材料与方法
1.1 研究区概况
研究区位于湖南省永州市双牌县东北隅的阳明山国家级自然保护区(26°02′00″~26°06′15″ N,111°51′36 ″~111°57′36″ E)的阳明山山脊处,最高海拔1 523 m,最低海拔为1 499 m;地势西南高,东北低,总面积近0.1 km2。气候属中亚热带季风湿润气候,该山脊区年均温11.8 ℃,1月平均气温1.06 ℃,7月平均气温20.2 ℃,年降水量1 756.59 mm,年均相对湿度86%。20世纪90年代,该山脊西北方最高处建设了微波台及附属建筑和通往该处的盘山混凝土道路,该道路呈“几”字形将山顶矮林分为4个区域。该区植被为典型的亚热带常绿阔叶林-山顶矮林植被[10-12]。
1.2 研究方法
1.2.1 野外群落学调查。
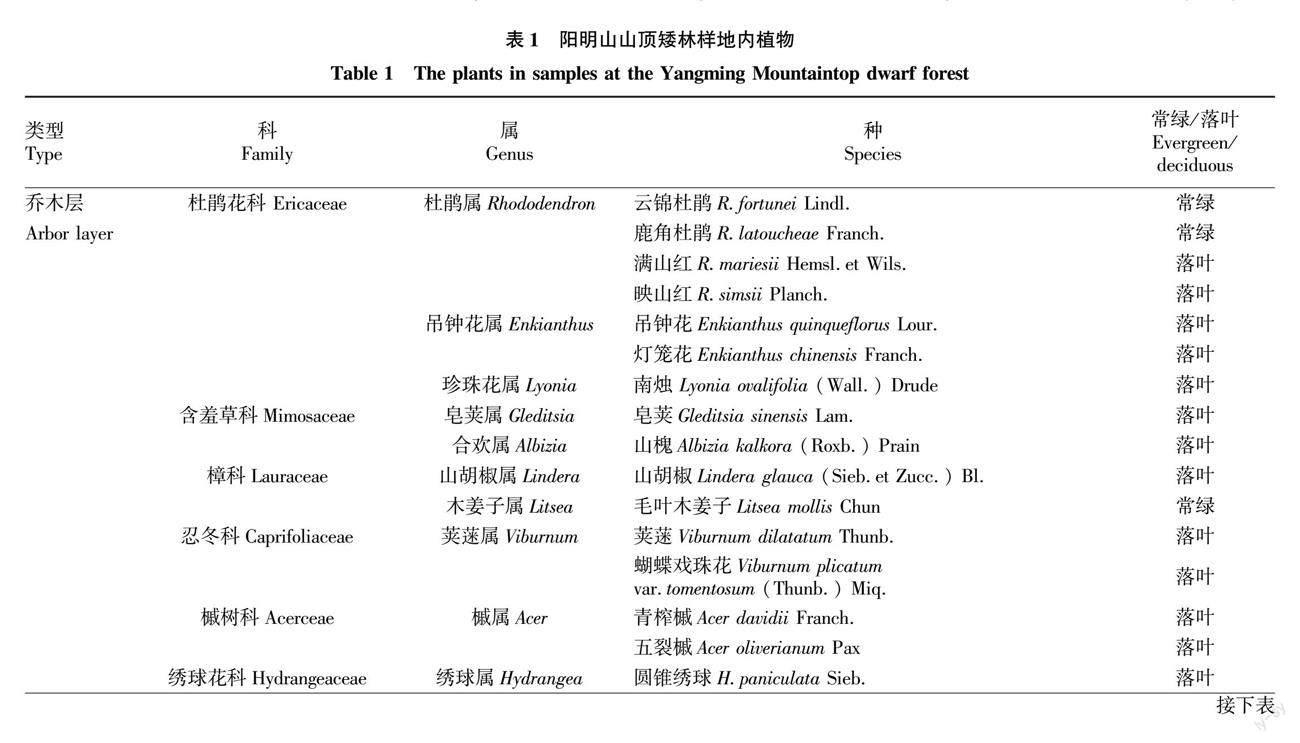
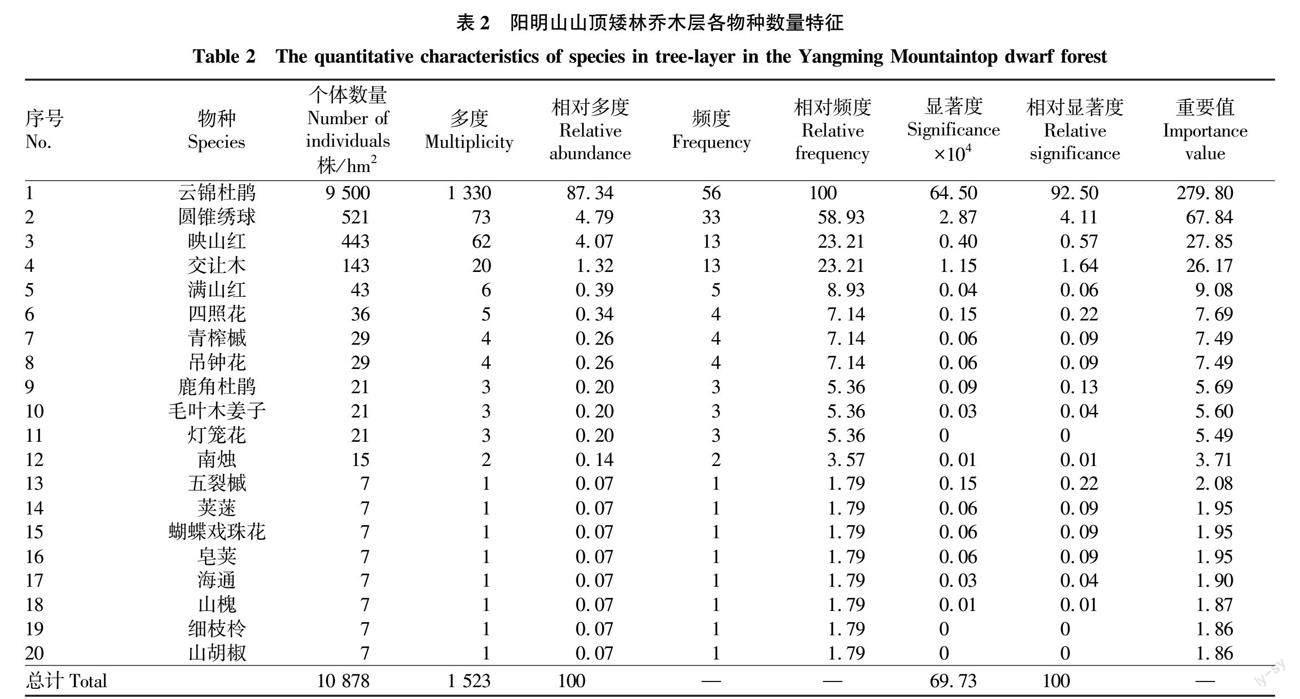
参照亚热带常绿阔叶林样地取样面积的范例与经验[13-15]及湖南阳明山山顶苔藓矮林种-面积曲线图[12],设置3个样地,分别为2个20 m×20 m样地(标记为S、E),1个30 m×20 m样地(标记为N) ,两两样地间距为15 m左右;采用相邻网格法划分成10 m×10 m样方和5 m×5 m小样方,对样地内植物进行每木调查,分别将高度在2.5 m以上,胸径2.0 cm以上的植物视为小乔木(因为地处山顶,生境条件导致该处植物高度不超过7.0 m),对小乔木分别登记植物名称、株数,测量树高、基径等;并在3个样地内各布设1个10 m×10 m的样方登记灌木层植物名称、高度、盖度和多度等指标;各布设5个1 m×1 m的小样方登记草本植物的名称、高度、多度和盖度等,同时记录出现在样地内的其他草本及藤本植物名称[14,16]。根据山顶矮林的特性,选用基径(DGH)作为径级分析指标,划分为4个级别[17-18]:Ⅰ 为幼苗,DGH≤2.5 cm;Ⅱ 为小树,2.5 cm 1.2.2 主要土壤因子的測定。 2016年8月上旬至2017年12月,在调查样地内,按“S”形路线先将枯枝落叶层扒开,然后采用土钻(土钻内径10 cm、筒长25 cm、全长120 cm)进行土壤取样;在各样地将采集到的同一层土壤样品混匀后放入准备好的自封袋中编号,带回室内后手工小心粉碎自然风干,剔除杂物,研磨过100目筛进行测定。主要选取土壤含水量(Moisture Content,MC)、pH、有机质(Organic matter,OM)、全氮(Total nitrogen,TN)、全磷(Total phosphorus,TP)、全钾(Total potassium,TK)、全钙(Total calcium,TCa)、全镁(Total magnesium,TMg)、全铁(Total ferro,TFe)和全锰(Total manganese,TMn)等全量元素指标,每个样品重复测定3次,取平均值。土壤MC测定采用环刀法(LY/T 1215—1999),MC采用%为单位;土壤OM测定采用重铬酸钾氧化还原法(LY/T 1237—1999),TN、TP、TK测定分别采用凯氏定氮法(LY/T 1228—1999)、氢氧化钠熔融法(LY/T 1232—1999)、氢氧化钠熔融法(LY/T 1234—1999),TCa、TMg、TFe、TMn的测定均参照《森林土壤分析方法》[19]。OM、TN、TP、TK、TCa、TMg、TFe、TMn以g/kg为单位。 1.2.3 数据处理与分析方法。 根据获取的资料,对湖南阳明山山顶矮林的群落特征进行全面总结分析,山顶矮林数量特征指标仅计算乔、灌、草3层,藤本和蕨类植物不计算。相关计算公式[13,20]如下: 相对多度=(某种植物个体数/全部植物个体数)×100(1) 相对高度=(某种植物高度总和/全部植物高度总和)×100(2) 相对显著度=(某种植物显著度总和/全部植物显著度总和)×100(3) 相对盖度=(某种植物盖度总和/全部植物盖度总和)×100(4) 式中,各物种在计算相对多度时,用独立个体的数量;计算相对显著度时,乔木层用基径处的显著度、灌木层和草本层植物运用相对盖度。计算相对频度时,乔木层总样方数为56个,灌木层总样方数为4个,草本层总样方数为15个。 科的重要值(FIV)计算公式[21]为 FIV = RΔ+ RA + RD(5) 式中,RΔ为相对多样性,是指一个科的树种数占总树种数的百分比;RA为相对多度,仅计算独立个体的数量;RD为相对显著度,乔木层计算RD用基径处的显著度包括分枝的显著度;对于灌木层和草本层植物运用相对盖度来表示相对显著度。 所有数据的统计分析和作图利用Excel 2010和SPSS 23.0软件完成。 2 结果与分析 2.1 山顶矮林物种组成、数量特征及群落结构 2.1.1 物种组成及数量特征。 山顶矮林样地内共记录到37科51属68种植物(不包含蕨类植物),其中乔木层植物有10科14属20种,灌木层有8科8属9种,草本层有13科20属28种,藤本植物8科9属11种,蕨类植物2科2属3种(表1)。 由表2可知,在阳明山山顶矮林样地内,云锦杜鹃种群无论是个体数量、占比、重要值、显著度及频度均占绝对优势地位,反映杜鹃花科杜鹃花属植物在亚热带山顶矮林中的优势地位;其次优种为圆锥绣球,再次是映山红。 在已知的29种木本植物中,10个(占总数的34.5%)常绿阔叶树种、19个(占总数的65.5%)落叶阔叶树种,无针叶树种(表1)。其中,10个常绿阔叶树种分别占总多度、总胸高断面积(总盖度)和总重要值的83.8%、99.7%和94.7%;19个落叶阔叶树种分别占总多度、总胸高断面积(总盖度)和总重要值的16.2%、0.3%和5.3%。体现了亚热带自然群落的以常绿阔叶树种为主、常绿与落叶树种混交的群落基本外貌规律性。 2.1.2 群落结构。 由表3可知,从群落高度与结构上看,山顶矮林所设样地内种群高度均在7.0 m以下,划分为3个亚层,映山红在第1亚层储备较多,云锦杜鹃储备相对较少,表明映山红种群更新较好,云锦杜鹃在今后的群落更新上可能会受到一定阻力;云锦杜鹃在第2亚层和第3亚层储备均丰富,种群相对稳定,在将来的群落发展中占据绝对优势地位,决定着未来山顶矮林群落的发展趋势;而映山红种群在第2亚层储备少,第3亚层更少,在将来的群落发展中处于劣势地位。从径级结构上看,云锦杜鹃和映山红种群的大小径级结构完整,而圆锥绣球和交让木种群径级结构有缺失,尤其是圆锥绣球种群幼苗和大树级均缺失,可以判定该种群在山顶矮林中可能处于衰退状态,而映山红和云锦杜鹃则呈现出增长型物种的发展态势。 2.2 山顶矮林物种组成与土壤因子的相关性 由图1可知,乔木层、灌木层和草本层各科的重要值与土壤TN、OM呈显著相关(P<0.05),表明土壤有机质为阳明山山顶矮林优势种云锦杜鹃的生长提供了养分,而TN促进了阳明山山顶矮林优势种云锦杜鹃高度与盖度的增加与生长。山顶矮朴各层重要值与TMg、TP、TK、pH及MC无显著相关性(P>0.05)。 3 讨论 3.1 山顶矮林物种组成、数量特征、群落结构特征 3.1.1 物种组成特征。 阳明山山顶矮林的统计资料显示,物种组成上反映出杜鹃花科杜鹃花属植物云锦杜鹃最丰富,在乔木层和灌木层中的优势地位明显,且落叶树种数超过常绿阔叶树种数,但常绿阔叶树种的多度、胸高断面积(盖度)和重要值均居首位且高达90%以上,常绿树种优势地位明显,呈现出以常绿阔叶树种为主,常绿与落叶阔叶树种混交的亚热带常绿阔叶林植物群落物种构成特征。 在自然界,当自然植物群落同特定环境发生相互作用后,形成特定群落,该特定自然群落就会形成自己独特的种类组成、外貌、层次和结构[3]。山顶矮林是我国亚热带地区典型植被类型的常绿阔叶林的一种变型[1],是由亚热带典型常绿阔叶林向高海拔山顶地带延伸并由山顶特有地形與气候所致[22],物种组成上以杜鹃花科为优势科,山茶科、壳斗科、樟科等居于次要地位[1,22-29],与同为亚热带地区江西井冈山江西垇地区散布成片居于凹地的云锦杜鹃矮林[29]相比,阳明山山顶云锦杜鹃矮林成片聚集于山顶山脊部位,群落结构与外貌景观独特。与云南山顶苔藓矮林和海南岛热带山顶矮林有所不同,云南山顶苔藓矮林以壳斗科、杜鹃花科为优势科,具热带高山起源特征,而海南岛热带山顶矮林则以樟科、茜草科、壳斗科和木樨科为优势科,具热带云雾林特征[8-9]。 3.1.2 种群数量特征。 湖南阳明山山顶矮林内云锦杜鹃种群数量波动较小,优势地位明显,表明山顶是其最适宜的生境区域。一般来说,如果群落内种群数量波动较小或相对恒定,则说明该种群生长在最适环境[15]。天然林中,密度限制是调节种内结构的重要机制,这种机制主要通过提高同种相邻个体的死亡率,加大同种个体之间的间距,以获得更多空间与资源,使自身径级增大[30]。该研究中,矮林内优势种群的各龄级及多年个体数量波动较小[11],这由于同种个体的自疏作用[11,31]及林内各物种种内与种间关系是影响山顶矮林物种数量、结构与多样性的主要原因。其二,山顶矮林中优势树种在数量、胸径显著性占绝对优势及上层郁闭度高,导致其他伴生物种及矮林内个体数量减少[16];同时随着物种间竞争加剧,优势物种的重要值上升,非优势物种的重要值下降,这是在群落演替过程中必然的发展趋势[32]。山顶矮林内云锦杜鹃占据绝对优势且历经多年地位稳定,也会在今后的群落演替中进一步推动整个山顶矮林向顶极状态发展。与此同时,种间竞争和云锦杜鹃的常绿性及优势地位所致的郁闭度加大,使矮林内其他物种地位进一步下降,不耐竞争的稀有种受优势种排挤加剧,导致矮林群落物种多样性进一步下降[33]。由多年生木本植物所构成的群落种群数量会在一定范围内波动,以保持群落相对的动态平衡状态[34]。笔者2005和2016年分别对该山顶矮林进行调查可知,云锦杜鹃个体数量和重要值均在一定范围内波动,且一直占据优势地位[11],维持山顶矮林在一个相对稳定的动态平衡中,由此也说明山顶的特定环境是云锦杜鹃种群生长的最适生境。 3.1.3 群落结构特征。 阳明山山顶矮林是已发育成熟、性质稳定,个体数量、年龄结构处于动态平衡的特殊森林群落。一般来说,由于森林群落内的各个种群所处的地位和功能不同,从而表现出3种不同的径级结构类型:一是年幼个体极多,年老的个体极少的增长型。二是各径级个体分布较均匀的稳定型。三是年幼个体比其他各级个体都少的衰退型[35]。该研究中,云锦杜鹃和映山红的幼树数量较少、小树丰富,中树最多,大树个体极少,大小径级结构完整,种群相对稳定;圆锥绣球和交让木2个种群幼树和大树个体缺失,且在调查中,圆锥绣球大多数立木枯萎,因而圆锥绣球种群可能处于衰退期。 3.2 物种组成与土壤因子的相关性 土壤是植被生存发展的物质基础,土壤有机质是反映土壤质量的重要指标之一,表征营养元素N、P等的可利用状态,较高的有机质含量即表征较高的土壤肥力,相反,较低的OM含量与土壤贫瘠紧密相关,且较高的土壤氮素利于植物生长,促进优势种的高度、盖度增加,过量的养分加快优势种的生长速率[36]。该研究中,阳明山山顶矮林集中分布区域土壤OM含量远低于云南和海南地区的山顶矮林[8-9],可见土壤贫瘠,但TN含量较高,高于海南山顶矮林土壤,低于云南地区,可能与云锦杜鹃根系具有能吸收利用各种氮素的杜鹃花类菌根有关[37];OM和TN与山顶云锦杜鹃矮林各层科重要值为显著正相关,表明土壤TN能显著影响山顶云锦杜鹃矮林植物的生长发育[38],土壤贫瘠是限制低海拔物种向山顶矮扩散、定居和生长的主要物质因素,因而在物种组成上优势种个体数超多,而非优势种个体数仅为个位数;由于山顶云锦杜鹃矮林林下光照异质性较低,难以为其他常绿物种生存提供相应的生态位,因而容纳的常绿树种物种数较少,落叶树种较多。该研究只分析了山顶矮林乔木层物种与土壤因子的部分特征,灌木层和草本层设置的样地数量稍显不足,有可能放大或缩小了某些环境因子的解释能力,然而如何量化每个环境因子影响物种组成及其多样性的作用力是今后的研究重点。 参考文献 [1] 吴征镒.中国植被[M].北京:科学出版社,1995. [2] 彭华,杨世雄,孔冬瑞.无量山山顶苔藓矮林植物区系特征研究[J].云南大学学报(自然科学版),2001,23(S1):5-10. [3] 苏雪痕.植物造景[M].北京:中国建筑工业出版社,1992. [4] 崔宁洁,张丹桔,刘洋,等.不同林龄马尾松人工林林下植物多样性与土壤理化性质[J].生态学杂志,2014,33(10):2610-2617. [5] SCHRIMPFF E,KLEMM O,EIDEN R,et al.Anwendung eines grunow-nebelfngers zur bestimmung von schadstoffgehalten in nebelniederschlgen[J].Staub-Reinhaltung Luft,1984,44(2):72-75. [6] TANNER E V J.Four montane rain forests of Jamaica:A quantitative characterization of the floristics,the soils and the foliar mineral levels,and a discussion of the interrelations[J].Journal of ecology,1977,65(3):883-918. [7] GRUBB P J.Control of forest growth and distribution on wet tropical mountains:With special reference to mineral nutrition[J].Annual review of ecology and systematics,1977,8:83-107. [8] 施济普.云南山顶苔藓矮林群落生态学与生物地理学研究[D].西双版纳:中国科学院研究生院(西双版纳植物园),2007. [9] 龙文兴.海南岛热带云雾林群落结构及组配机制研究[D].北京:中国林业科学研究院,2011. [10] 陈艳华,彭重华,肖育檀.湖南阳明山野生杜鹃花属植物[J].亚热带农业研究,2005,1(4):8-11. [11] 陈艳华,龙岳林,彭重华.湖南省阳明山山顶苔藓矮林物种组成与多样性的十年动态[J].应用生态学报,2018,29(7):2111-2120. [12] 陈艳华,龙岳林,彭重华.湖南省阳明山山顶苔藓矮林优势种空间分布格局[J].生态学报,2019,39(18):6690-6700. [13] 张金屯.数量生态学[M].北京:科学出版社,2004. [14] 方精云,王襄平,沈泽昊,等.植物群落清查的主要内容、方法和技术规范[J].生物多样性,2009,17(6):533-548. [15] 王伯荪,李鸣光,彭少麟.植物种群学[M].广州:广东高等教育出版社,1995. [16] 杨秀清,史婵,王旭刚,等.关帝山寒温性云杉次生林样地树种组成、结构和空间分布[J].应用生态学报,2017,28(5):1421-1430. [17] 马克平,刘灿然,于顺利,等.北京东灵山地区植物群落多样性的研究Ⅲ.几种类型森林群落的种-多度关系研究[J].生态学报,1997,17(6):573-583. [18] 曲仲湘,文振旺,朱克贵.南京灵谷寺森林现况的分析[J].植物学报,1952(1):18-49. [19] 国家标准局.森林土壤分析方法[S].北京:中国标准出版社,1999. [20] 张维康,李贺,王国宏.北京西北部山地两个垂直样带内主要植被类型的群落特征[J].植物生态学报,2013,37(6):566-570. [21] LINARES-PALOMINO R,PONCE ALVAREZ S I.Tree community patterns in seasonally dry tropical forests in the Cerros de Amotape Cordillera,Tumbes,Peru[J].Forest ecology & management,2005,209(3):261-272. [22] 解丹丹,蘇志尧.广东石门台亚热带典型常绿阔叶林和亚热带山顶矮林群落特征[J].植物科学学报,2014,32(4):355-361. [23] 《云南植被》编写组.云南植被[M].北京:科学出版社,1987. [24] 徐捷,王希华.中国山顶苔藓矮曲林的分布及其特征[J].华东师范大学学报(自然科学版),2010(4):44-57. [25] 姚永慧,张百平,赵超.中国矮曲林的分布特征及生态意义[J].地理科学进展,2017,36(4):491-499. [26] 叶居新.中国的猴头杜鹃矮林[J].武汉植物学研究,1994,12(2):170-174. [27] 朱守谦,杨业勤.贵州梵净山山顶苔藓矮林初步研究[J].植物生态学与地植物学学报,1987,11(2):92-105. [28] 邓贤兰,吴杨,刘玉成,等.井冈山猴头杜鹃群落特征的研究[J].生态环境学报,2011,20(10):1430-1435. [29] 邓贤兰,刘鹏,吴杨,等.井冈山云锦杜鹃群落特征研究[J].亚热带植物科学,2011,40(4):20-25. [30] 祝燕,白帆,刘海丰,等.北京暖温带次生林种群分布格局与种间空间关联性[J].生物多样性,2011,19(2):252-259. [31] 刘贵峰,丁易,臧润国,等.天山云杉种群分布格局[J].应用生态学报,2011,22(1):9-13. [32] 黄云霞,徐萱,张莉芗,等.百山祖常绿阔叶林灌草层物种组成和分布的10年动态[J].生物多样性,2016,24(12):1353-1363. [33] 张梦弢,张青,亢新刚,等.长白山云冷杉林不同演替阶段群落稳定性[J].应用生态学报,2015,26(6):1609-1616. [34] 毕润成.山西霍山山核桃群落生态特征及其区系分析[J].应用生态学报,1999,10(6):650-656. [35] 杨允菲,祝廷成.植物生态学[M].北京:高等教育出版社,2016. [36] 崔倩,潘存德,李贵华,等.喀纳斯泰加林群落物种多样性环境解释与自然火干扰[J].生态学杂志,2018,37(6):1824-1832. [37] 张春英.云锦杜鹃菌根及其菌根真菌多样性研究[D].北京:北京林业大学,2008. [38] VONLANTHEN C M,BHLER A,VEIT H,et al.Alpine Plant Communities:A statistical assessment of their relation to microclimatological,pedological,geomorphological,and other factors[J].Physical geography,2006,27(2):137-154.
猜你喜欢
热带农业科学(2016年11期)2017-01-21
热带农业科学(2016年11期)2017-01-21
河北渔业(2015年12期)2015-12-21
河北渔业(2015年11期)2015-11-18
河北渔业(2015年11期)2015-11-18
河北渔业(2015年10期)2015-10-15
河北渔业(2015年7期)2015-07-21
河北渔业(2015年6期)2015-06-24
安徽农学通报(2015年5期)2015-03-23
安徽农学通报(2015年5期)2015-03-23
