
光谱对许氏平鲉消化代谢节律及生理应激研究
2024-01-13 05:52房莹莹陈璐璐刘松涛吴燕玲
海洋科学 2023年10期
房莹莹, 陈璐璐, 刘松涛, 孙 飞, 吴燕玲, 李 鑫, 刘 鹰, 马 贺
光谱对许氏平鲉消化代谢节律及生理应激研究
房莹莹1, 2, 陈璐璐1, 2, 刘松涛1, 2, 孙 飞1, 2, 吴燕玲1, 2, 李 鑫1, 2, 刘 鹰1, 3, 马 贺1, 2
(1. 大连海洋大学海洋科技与环境学院, 辽宁 大连 116023; 2. 设施渔业教育部重点实验室, 辽宁 大连 116023; 3. 浙江大学生物系统工程与食品科学学院, 浙江 杭州 310058)
为探究不同光谱下许氏平鲉的消化代谢水平及外周激素节律, 在5种光谱(红光、绿光、黄光、蓝光、白光, 光周期12L∶12D)下, 通过对许氏平鲉的血清、肠道、肝脏进行24 h(8: 00, 12: 00, 16: 00, 20: 00, 24: 00)取样, 结果显示: 不同的光谱下许氏平鲉的消化代谢酶活性除胰蛋白酶(TRY)外均具有节律性, 红光、黄光下α-淀粉酶(α-AMS)、脂肪酶(LPS)活性的峰值相位与白光组相比左移; 胰蛋白酶活性无时间差异性; 在绿光、黄光及红光下乳酸脱氢酶(LDH)活性的峰值相位与白光组相比发生左移, 在五种光谱环境中, 红、黄光影响消化酶活性峰值提前出现; 丙酮酸激酶(PK)活性在绿、蓝及红光下峰值相位左移; 己糖激酶(HK)的活性在绿、蓝、黄光下, 其峰值相位与白光组相比发生左移, 在五种光谱环境中, 绿、蓝光影响代谢酶活性类峰值提前出现。许氏平鲉血清中褪黑素含量呈现昼低夜高分泌水平; 皮质醇的含量表现为在白天降低, 午夜显著升高至最高值后又持续降低。峰值相位的改变预示着生理节律的变动, 以上结果表明不同光谱可以影响鱼类的生理代谢节律, 在今后养殖过程中需要充分考虑光谱对养殖生物的生物学作用, 进而制定更为合理的光照条件。
许氏平鲉; 光谱; 昼夜节律; 消化代谢; 应激
在自然界中, 几乎所有生物体均具有昼夜节律, 称为昼夜节律钟(circadian time), 参与调节生物体的生理、生化功能和行为特征, 其标志是机体核心温度和血浆中褪黑素、皮质醇的含量[1]。在控制昼夜节律的众多因素中, 光(包括三个要素, 分别为光周期、光强和光谱)作为最重要的因素之一, 不仅影响生物体的生理, 也影响生殖和行为。科学家们早在20世纪90年代就已经开始了光照对水生生物影响的研究, 随着研究的不断深入, 越来越多的研究结果表明, 光照作为环境因子之一, 可以直接或间接地影响水生生物, 光照具有多方面的生态作用, 特别是对鱼类的生长发育、摄食、存活、繁殖、行为、生物节律等方面的均有重要影响[2]。
光可以诱导或抑制水生生物节律中的激素分泌, 影响生殖能力、生长和行为[3]。有关光照对鱼类生理节律的影响研究有较多报道。如, 学者发现真鲷()的生长及饲料利用率随着光照时间的延长而增加[4], 而在大菱鲆[5]及南极石首鱼()[6]上的研究上则发现长期光照对鱼类的生长及饲料利用具有负面影响。另外, 还有学者发现长光照或短光照会改变花鲈()昼夜节律的振荡强弱, 也会改变其峰值相位[7]; 光照对刺参()的摄食节律和消化酶(脂肪酶、淀粉酶、胰蛋白酶)活力昼夜变化都有显著影响, 刺参具有提前分泌消化酶为即将到来的摄食做好准备的调节机制[8]。目前有关光照对鱼类生理节律的研究多集中在光周期因子上,而光谱是否也会对鱼类的生理节律产生影响鲜有报道。
许氏平鲉()隶属于鲉形目(Scorpaeniformes)、鲉科(Scorpaenidae)、平鲉属(),又称黑鲪、黑寨等, 为卵胎生, 洄游范围较小, 主要分布于西北太平洋沿岸地区, 特别是在东海、黄海以及日本和韩国的沿海地区。许氏平鲉口感鲜嫩, 营养价值高, 富含多种营养物质, 深受消费者的喜爱, 为黄、渤海地区近岸底层的主要经济鱼类, 同时也是近海网箱饲养和池塘放养的主要鱼类之一[9]。目前在鱼类中, 生物钟的研究对象主要为一些模式物种, 如斑马鱼()、红鳍东方鲀()、青鳉()等[10-12]; 仅有少量的养殖物种, 如欧洲鲈()、虹鳟()、大西洋鲑()、大西洋鳕()等[13-16]进行过生物钟相关研究。目前国内外对于许氏平鲉大多数集中在生殖、消化道、摄食等方面, 而关于光谱对其昼夜节律的影响还未见报道。许氏平鲉养殖过程中相关的光照调控技术更是缺乏, 因此, 研究光谱对许氏平鲉生理节律的影响, 对探究鱼类行为生理生态学、丰富并完善养殖光照调控技术、提高生产效率及生物福利水平等具有重要的理论指导意义。
本研究针对光谱对鱼类生理节律影响尚不明晰的问题, 在实验室条件下, 设置五种不同的光谱(白光、蓝光、绿光、黄光、红光)对许氏平鲉进行养殖处理, 通过监测其组织中消化代谢酶活性及血清中激素水平的昼夜变化规律, 探究不同光谱对许氏平鲉生理节律的影响, 通过检测许氏平鲉的昼夜阶段各指标差异, 解析光谱对许氏平鲉激素水平及生理指标变化的昼夜节律模式, 揭示光谱对鱼类生物钟节律的影响规律, 为实际养殖中光环境的调控提供理论参考。
1 材料和方法
1.1 实验材料
实验用鱼来自大连海洋大学水产设施养殖与装备工程技术研究中心实验室的循环水实验系统, 许氏平鲉90日龄, 驯养一周后从中挑选450尾体质优良、大小均匀平均体质量为[(36±2.93) g, 平均体长(10.07±0.56) cm)]的许氏平鲉进行实验。
1.2 实验方法
本实验设置5个LED光谱处理组: 绿光(波长525~ 530 nm)、蓝光(波长450~455 nm)、黄光(波长590~ 595 nm)、红光(波长625~630 nm)、白光(波长400~ 780 nm)。实验采用LED光源, 光谱的选择及具体布置方式均参考魏平平等人已发表的研究进行[17]。每个处理组下设置3个重复即放置3个养殖桶(内径: 80 cm, 水深: 60 cm, 有效水体体积: 250 L), 每个重复养殖30尾许氏平鲉幼鱼, 采用流水养殖, 光照强度统一设定为250 mW/m2, 光周期设定为12L∶12D(开灯时间7: 30、关灯时间19: 30), 实验周期为75 d。为了避免各处理组之间光源的交叉污染, 每个处理组之间均用遮光布隔开。每日上午8: 30和下午16: 00各饱食投喂一次, 实验期间投喂商业饲料, 每日投喂饲料的重量按每养殖桶内鱼体总质量的2%进行计算。每天吸底2次, 用于清理残饵粪便。
1.3 样品采集与测定
作为实验初始样本, 实验结束后, 实验用鱼均需停食24 h, 每个养殖桶随机抽取3尾作为实验后期样本。样品采集在一昼夜进行, 每隔4 h进行取样(每个时间点分别为8: 00, 12: 00, 16: 00, 20: 00, 24: 00, 第二天04: 00, 8: 00, 分别对应时区(zone time, ZT) ZT4、ZT8、ZT12、ZT16、ZT20、ZT24)。取样使用MS—222(200 mg/L)将鱼麻醉, 使用肝素钠粉末, 配制成1%肝素生理盐溶液抗凝剂, 将抗凝剂润湿注射器管壁, 从尾静脉抽取血液, 分开放入1.5 mL离心管中, 4 ℃静置一夜离心, –80 ℃冰箱保存备用, 每个时间点抽血后的实验用鱼立即活体解剖取其肠、肝脏。迅速置于液氮中, –80 ℃冰箱保存备用。
1.3.1 消化代谢酶活性的测定
样品分析前于4 ℃冰箱内解冻, 在冰盘剪碎并迅速准确称取组织重量(称取0.1 g组织), 加入9倍体积的匀浆介质, 在冰水浴条件下机械匀浆, 制成10%匀浆, 4 ℃下2 500 r/min离心10 min, 取上清液测定消化代谢酶活力。肠道组织用于测定消化酶, 包括: α-淀粉酶(α-AMS)、胰蛋白酶(TRY)和脂肪酶(LPS); 肝脏组织用于测定代谢酶, 包括: 丙酮酸激酶(PK)、乳酸脱氢酶(LDH)和己糖激酶(HK)。所有酶活力测定均严格按照由南京建成生物工程研究所生产的试剂盒说明书进行。
1.3.2 血清皮质醇、褪黑素含量的测定
血清褪黑素、皮质醇含量测定按照酶联免疫分析(ELISA)试剂盒(酶联生物)的操作说明书使用双抗体夹心法进行测定, 使用酶标仪450 nm波长下测定吸光度(OD值), 通过标准曲线计算样品中褪黑素、皮质醇含量。
1.4 数据统计分析
实验数据经Excel 2016初步处理后, 均以平均值±标准差表示。用 SPSS 22.0 软件进行单因素方差分析(One-way ANOVA), 时间差异显著性(ZT4、ZT8、ZT12、ZT16、ZT20、ZT24), 结果表示为<0.05。用Matlab 软件进行余弦分析, 拟合余弦方程为()=+cos(/12−); 其中()是指时间对应的酶活性水平;为波动变化的中线称为中值;为振幅;为峰值相位, 是振荡达到峰值的时刻[13]。
2 结果与分析
2.1 不同光谱下许氏平鲉消化酶活性的昼夜变化
5种光谱条件下, 许氏平鲉肠内各消化酶: α-淀粉酶、脂肪酶活性有显著性时间差异 (<0.05), 胰蛋白酶活性的时间差异不显著。各消化酶在肠中活性的昼夜节律性参数见表1。
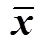
表1 许氏平鲉消化酶活性的昼夜节律性参数
注: 振幅为拟合波形峰值之间距离的一半; 中值为周期平均值; 峰值为相位周期最高幅度的时间点 (弧度);为不同时间点间比较的差异; “—” 表示时间差异不显著。
2.1.1 不同光谱下许氏平鲉肠中α-淀粉酶活性的昼夜变化
5种光谱下许氏平鲉肠道淀粉酶活性的昼夜节律如图3所示。结果显示, 绿光下淀粉酶的活性在ZT24达到最大值。在8 h(ZT8)后降到最低点(图1a), 活性较低。蓝光组中, 淀粉酶的活性于ZT12至ZT24之间显著增高<0.05), 并在ZT24达到峰值0.90后缓慢降低(图1b)。黄光组中, 淀粉酶的活性在ZT8-ZT16内显著增高, 并在ZT16达到最大值1.32, 在黑暗条件下ZT4降到最低点(图1c)。红光组中淀粉酶活性在ZT8到ZT16之间呈上升趋势, 在ZT16达到最大值后出现下降趋势, 在ZT20继续缓慢下降在ZT24达到最低值, 随后呈显著上升趋势(<0.05, 图1d)。白光组中出现两个峰值分别为ZT16、ZT24, 淀粉酶活性在ZT16达到最大后在4 h内显著下降<0.05), 在ZT24达到峰值后显著下降, 在ZT8达到最低值(图1e)。绿、蓝光在夜间淀粉酶活性水平相对较多高, 呈现夜高昼低现象, 而黄光与红光组则呈现昼高夜低现象, 白光组峰值则出现在昼夜交替时, 无明显的昼高夜低现象。绿光、蓝光下许氏平鲉消化酶的活性水平在ZT24达到峰值, 蓝光与绿光环境相比峰值酶活性水平较高, 黄光和红光的峰值出现在ZT16, 黄光与红光环境相比峰值活性水平较高, 白光组出现两个消化酶的活性峰值分别为ZT16和ZT24。与白光相比, 黄光和红光下酶活性的峰值相位均有不同程度的左移, 而绿、蓝光下酶活性峰值相位出现的时间均出现右移。就平均活性来看(图1f), 白光下淀粉酶平均活性量显著高于其他光色, 蓝光、黄光、红光显著高于绿光下的平均活性<0.05)。
2.1.2 不同光谱下许氏平鲉肠中脂肪酶活性的昼夜变化
5种光谱下许氏平鲉肠道脂肪酶活性的昼夜节律及平均活性水平变化如图2所示。绿光下脂肪酶的活性在ZT16出现峰值后缓慢下降, 在ZT4达到最低活性, 随后在4 h后显著升高(<0.05, 图2a)。蓝光组分别在ZT20、ZT4出现峰值, 最大值为ZT20, ZT12到ZT20缓慢上升达到峰值后在ZT24显著下降(< 0.05), 随后呈上升趋势(图2b)。黄光组各组间无显著性差异, 在ZT12脂肪酶的活性最低(图2c)。红光组中, ZT8-ZT12显著下降(<0.05), 4 h后第三个时间点呈显著上升趋势, 在ZT20-ZT4呈缓慢下降趋势, 在最后时间点脂肪酶的活性升高(图2d)。白光组中峰值出现在ZT16, 在ZT8-ZT12呈现显著下降趋势, 随后逐渐升高并在ZT16达到峰值, 在ZT4出现最低活性, 随后上升(图2e)。在绿、蓝光下出现昼高夜低现象, 绿光环境下活性较高, 绿、蓝光组的酶活性的峰值相位相较黄、红光组发生右移。与白光相比, 绿、蓝光下酶活性的峰值相位出现的时间均发生左移。在脂肪酶平均活性水平中(图2f), 各光谱处理组活性无明显差异。
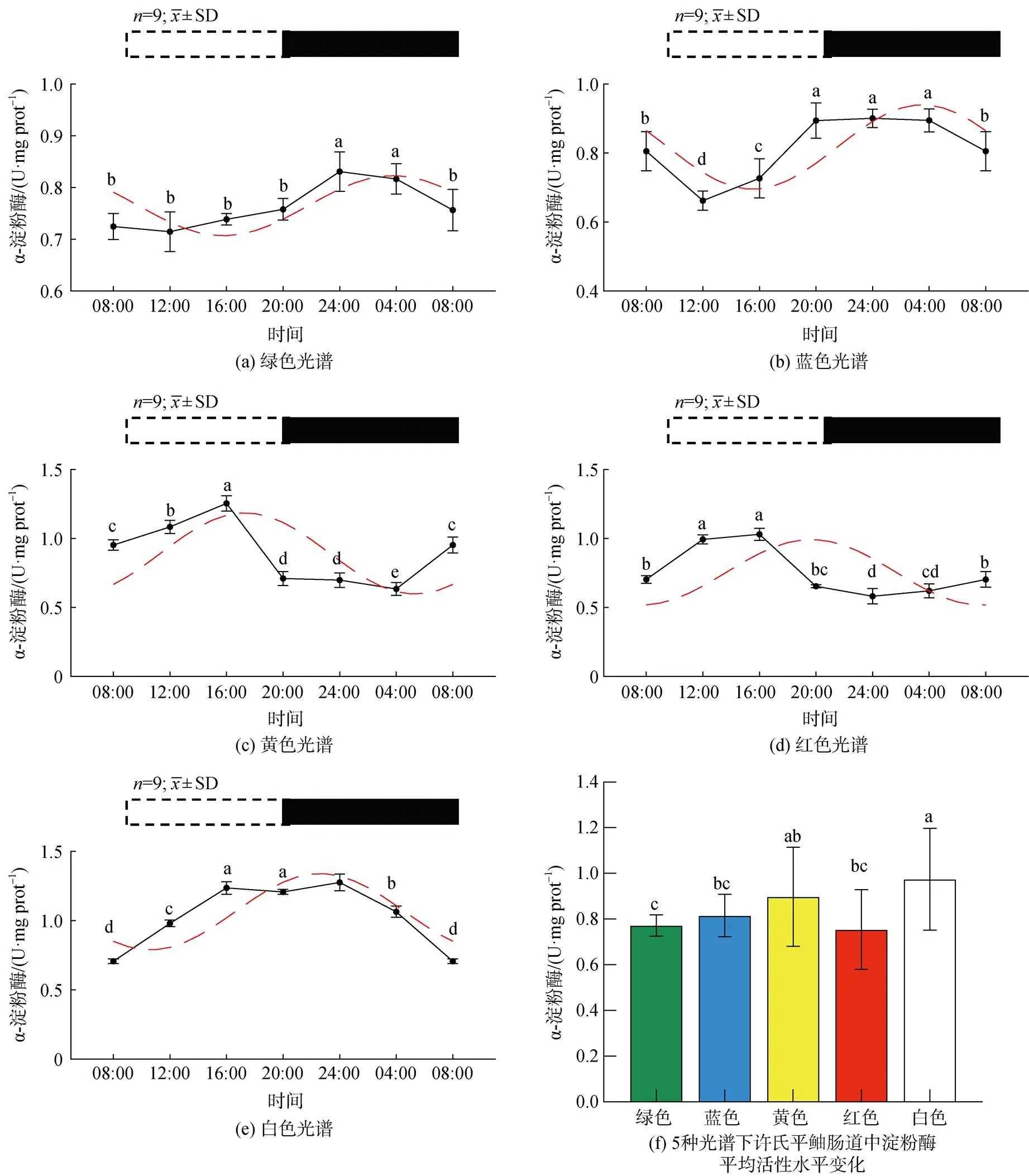
图1 5种光谱下许氏平鲉肠道淀粉酶活性水平的昼夜节律及平均活性水平变化
注: 统计时间点之间的差异(<0.05)由不同的字母符号(a, b, c)表示。图片上方的条状图为光周期, 白色代表光照阶段, 黑色代表黑暗阶段
2.1.3 不同光谱下许氏平鲉肠中胰蛋白酶活性的昼夜变化
5种光谱下许氏平鲉肠道胰蛋白酶活性的昼夜节律及平均活性水平变化如图3所示。绿光组中, 胰蛋白酶的活性在ZT8最高, 在ZT8-ZT20显著下降(<0.05), ZT20-ZT24呈上升趋势, 随后下降(图3a)。蓝光组中, 在光照期阶段显著下降(<0.05), 在ZT16- ZT20持续升高达到峰值后继续显著下降(<0.05), 在ZT4到达最低点, 随后升高(图3b)。黄、红、白光组的趋势大致相同, 峰值出现在ZT12-ZT16(图3 c、d、e), 绿光下胰蛋白酶活性的峰值在五种光谱环境中最低。黄、红、白光环境下胰蛋白酶活性呈现昼高夜低的现象, 胰蛋白酶的平均活性水平各组间无明显差异(图3f)。
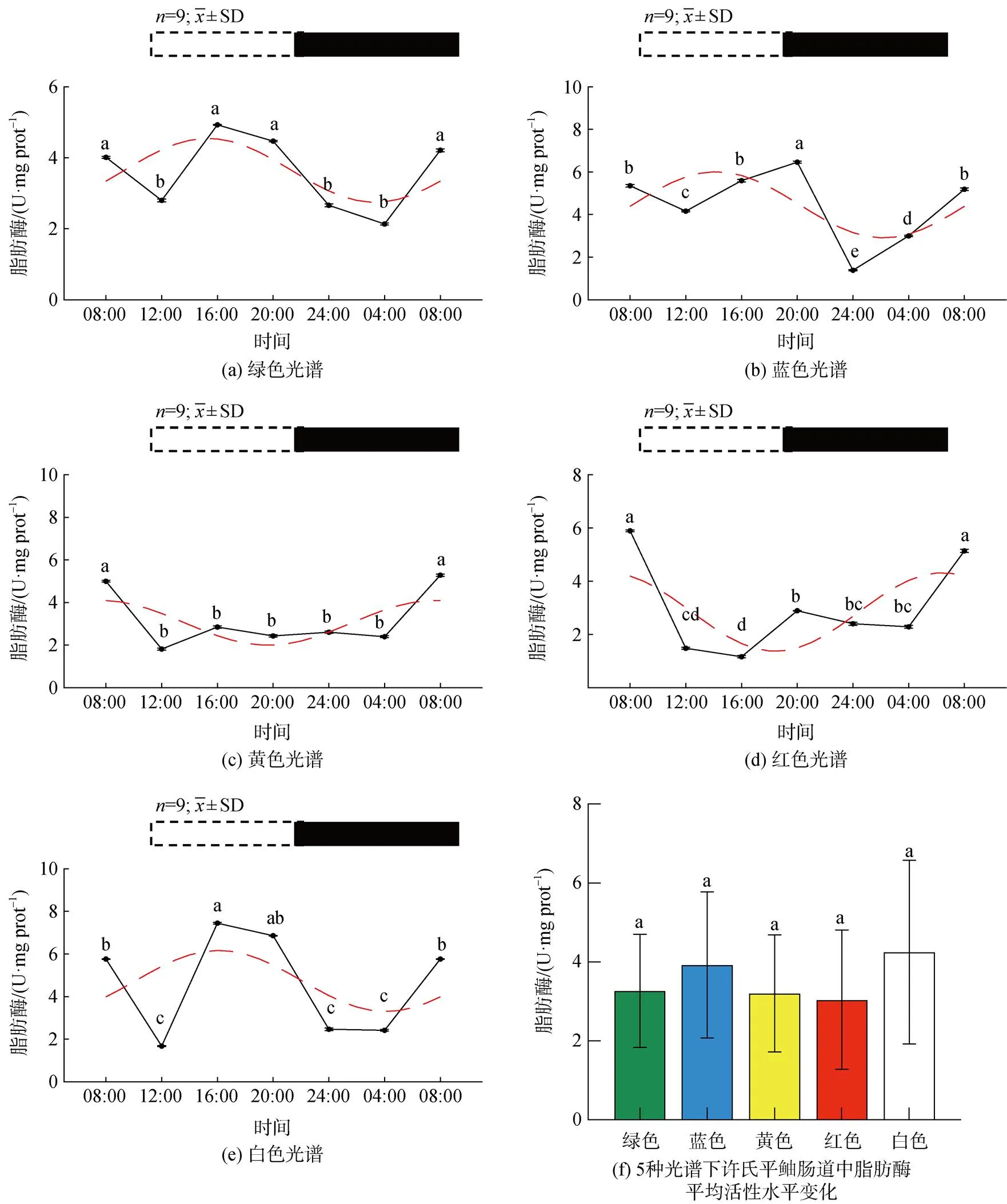
图 2 5种光谱下许氏平鲉肠道脂肪酶活性水平的昼夜节律及平均活性水平变化
注: 统计时间点之间的差异(<0.05)由不同的字母符号(a, b, c)表示。图片上方的条状图为光周期, 白色代表光照阶段, 黑色代表黑暗阶段
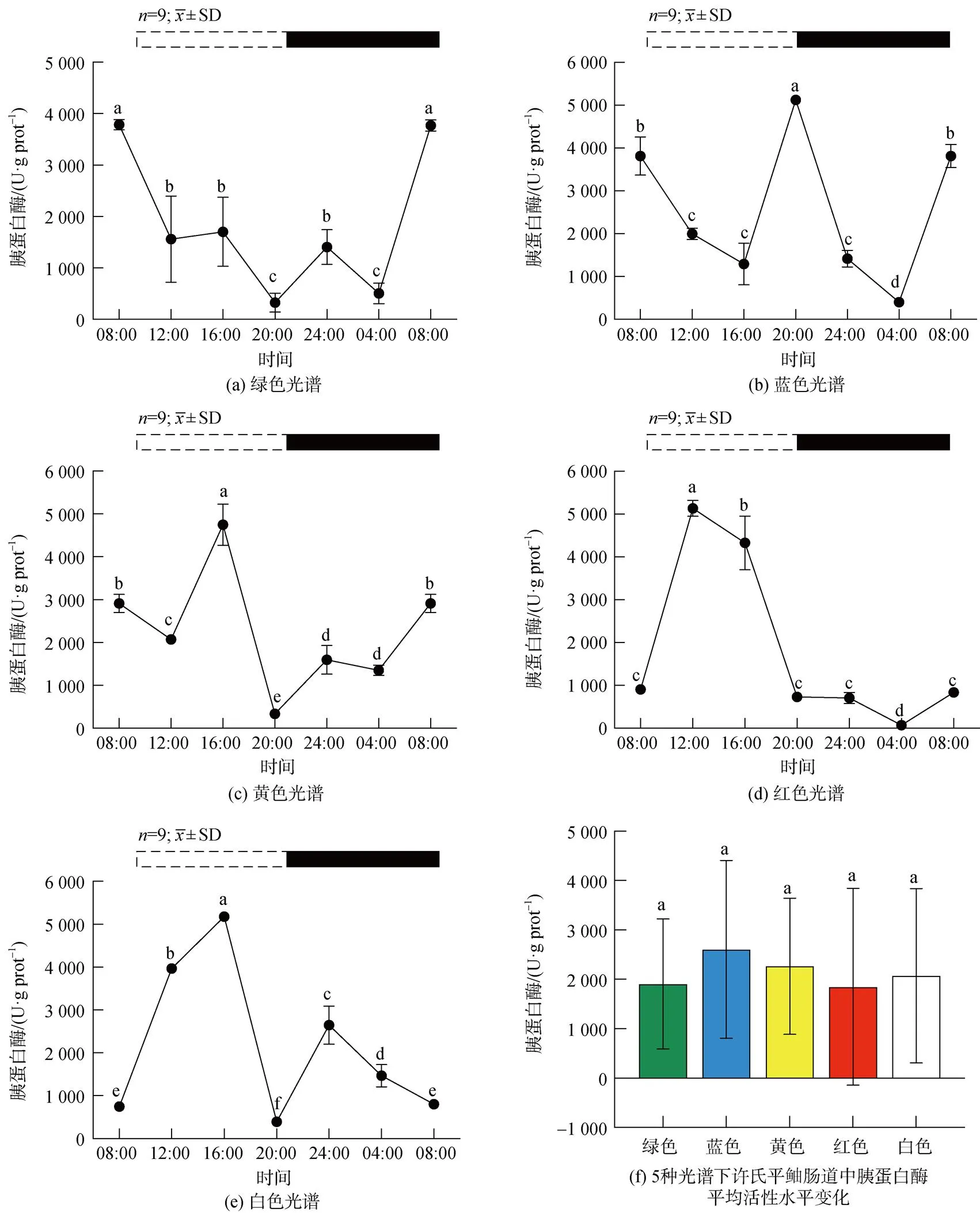
图 3 5种光谱下许氏平鲉肠道胰蛋白酶活性水平的昼夜节律及平均活性水平变化
注: 统计时间点之间的差异(<0.05)由不同的字母符号(a, b, c)表示。图片上方的条状图为光周期, 白色代表光照阶段, 黑色代表黑暗阶段
2.2 不同光谱下许氏平鲉肝脏中代谢活性的昼夜变化
5种光谱条件下, 许氏平鲉肝脏内各代谢酶: 己糖激酶、乳酸脱氢酶、丙酮酸激酶的活性有显著性时间差异 (<0.05), 各代谢酶在肝脏中活性的昼夜节律性参数见表2。
表2 许氏平鲉代谢酶活性的昼夜节律性参数
注: 振幅为拟合波形峰值之间距离的一半; 中值为周期平均值; 峰值为相位周期最高幅度的时间点 (弧度);为时间点间的差异
2.2.1 不同光谱下许氏平鲉肝脏中己糖激酶活性昼夜变化
5种光谱下许氏平鲉肝脏中己糖激酶活性的昼夜节律及平均活性水平变化如图5所示。绿光组中, 在ZT8-ZT24中基本保持缓慢下降趋势, 在ZT24达到最小值, 随后呈升高趋势(图4a)。蓝光组中, 光照期阶段呈下降趋势, 在ZT24达到最低值, ZT24- ZT8呈上升趋势(图4b)。黄光下在ZT8-ZT24保持缓慢下降趋势, 各组差异不大, 随后呈逐渐上升趋势(图4c)。红光和白光组(图4e)均在ZT16达到峰值, 趋势大致相同(图4d、e)。白光组峰值活性水平在五种光谱环境中最高, 绿、蓝、黄光组峰值相位左移, 各光谱组均出现昼高夜低现象, 峰值相位均出现在白天。就平均活性水平来看(图4f), 红光平均活性显著高于其他各光色组, 绿、蓝光活性显著低于其他光色组。
2.2.2 不同光谱下许氏平鲉肝脏中乳酸脱氢酶活性的昼夜变化
5种光谱下许氏平鲉肝脏中乳酸脱氢酶活性的昼夜节律及平均活性水平变化如图5所示。绿光组中,酶活性在ZT8-ZT12期间呈下降趋势, 在ZT12时出现最小值, 4 h后出现峰值, 在光暗时期呈逐渐下降趋势(图5a)。蓝光组中乳酸脱氢酶活性水平呈缓慢上升趋势, 在ZT16达到峰值, 在4 h后显著下降(<0.05), 随后呈缓慢上升趋势(图5b)。黄光组中, 在分别在ZT20、ZT4出现峰值。红光组(图5d)中, 在ZT12达到最低值随后呈缓慢上升趋势, ZT24达到峰值后保持缓慢下降趋势(图5c)。白光组中, ZT8-ZT16呈缓慢下降趋势, 4 h后呈显著上升趋势(<0.05), 在ZT20达到最大值, 随后呈下降趋势, 在ZT24达到最低值(图5e)。白光组峰值活性水平在五种光谱环境中最高, 红光组出现夜高昼低现象, 与白光组相比, 绿、黄、红组峰值相位左移, 蓝光组发生右移。各光色平均活性水平无明显差异(图5f)。
2.2.3 不同光谱下许氏平鲉肝脏中丙酮酸激酶活性的昼夜变化
5种光谱下许氏平鲉肝脏中丙酮酸激酶活性的昼夜节律及平均活性水平变化如图6所示。绿光组呈现两极化变化, 分别在ZT16、ZT24达到峰值, 在光周期第一时间段呈现下降趋势, 4 h后逐渐升高, 在黑暗期第一时间段显著降低(<0.05), ZT20-ZT24缓慢上升, 4 h后呈缓慢下降趋势, ZT4-ZT8显著上升(<0.05, 图6a)。蓝光组中, 光周期第一时间段缓慢上升, 在ZT16达到峰值, 随后缓慢保持下降趋势, 在ZT4达到最低值, 随后在ZT4-ZT8显著升高(<0.05, 图6b)。
黄光组在ZT12的活性达到峰值, 4 h后显著降低(<0.05), 在ZT16-ZT4变化缓慢后呈上升趋势(图6c)。红光组中, 酶的活性水平一直保持下降趋势, 在ZT20达到最低值, 随后呈现上升趋势(图6d)。白光组中, 酶的活性水平呈现缓慢下降趋势, 在ZT20达到最低值。随后升高, 在ZT24达到峰值, 4 h后呈现缓慢上升趋势(图6e)。黄光组峰值活性在五种光谱环境中最高, 各光谱组的丙酮酸激酶的活性均出现昼高夜低现象。白光组与绿、蓝光、红光组相比峰值相位右移。黄光与白光组峰值相位基本一致。平均活性水平无明显差异(图6f)。
2.3 不同光谱下许氏平鲉血清皮质醇含量变化
皮质醇在许氏平鲉血清中表现出的含量变化如图7。绿光组中, 含量呈现出两极性的变化, 光周期阶段(ZT8-ZT12)显著上升, 直至第3个时间点(ZT16)持上升趋势, 并在ZT16达到峰值, 随后缓慢降低, 继续保持缓慢上升的趋势, 直到ZT24呈现下降趋势(图7a)。蓝光组中, 在光周期阶段(ZT8- ZT20)直至黑暗阶段(ZT4)保持上升趋势, ZT4激素的含量为最高峰, 随后呈现下降趋势(图7b)。黄光、红光组中, ZT8至ZT24保持上升趋势, 在第一个黑暗阶段(ZT24)出现缓慢下降趋势, ZT24达到峰值 (图7c、d)。白光组中, 峰值出现在ZT12, 在ZT12-ZT20显著下降(<0.05), 在ZT20-ZT4保持缓慢上升趋势(图7e)。与白光组相比, 蓝、黄、红光组峰值均出现在黑暗时期, 峰值为ZT24-ZT4。各光谱组皮质醇无明显昼夜节律, 白光组平均分泌量显著低于其他光谱组(图7f)。
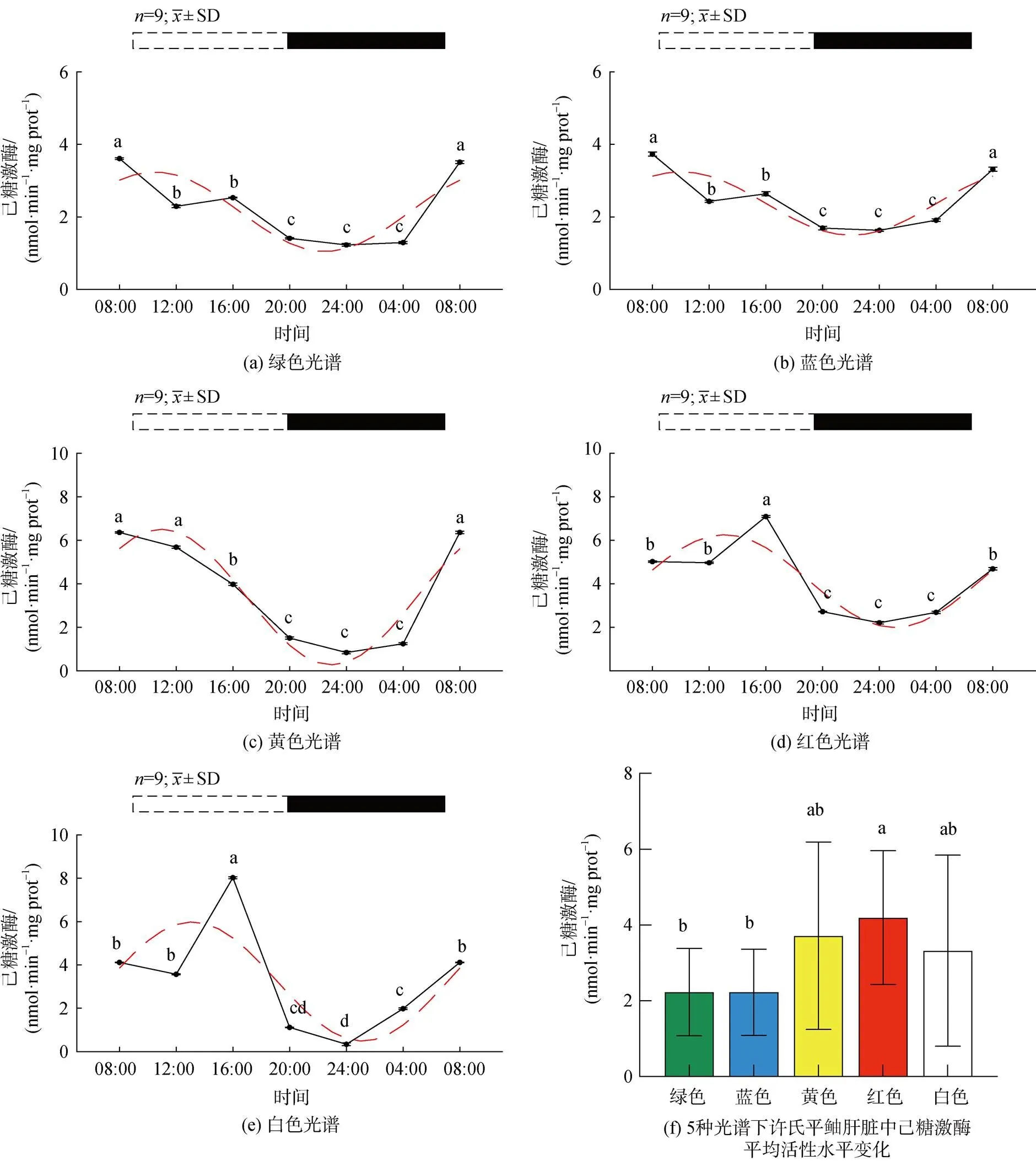
图4 5种光谱下许氏平鲉肝脏中己糖激酶活性水平的昼夜节律及平均活性水平变化
注: 统计时间点之间的差异(<0.05)由不同的字母符号(a, b, c)表示。图片上方的条状图为光周期, 白色代表光照阶段, 黑色代表黑暗阶段
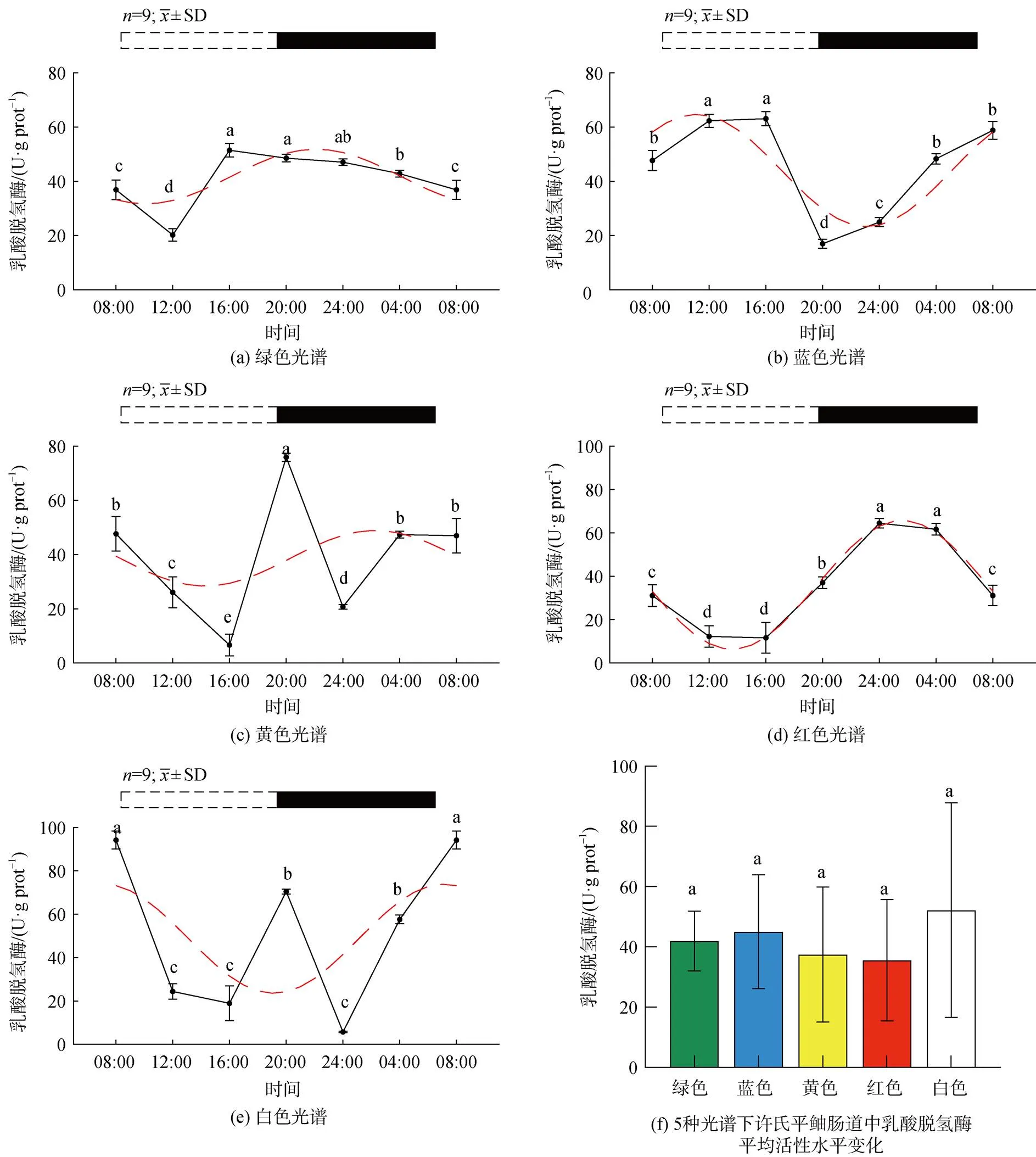
图5 5种光谱下许氏平鲉肝脏中乳酸脱氢酶活性水平的昼夜节律及平均活性水平变化
注: 统计时间点之间的差异(<0.05)由不同的字母符号(a, b, c)表示。图片上方的条状图为光周期, 白色代表光照阶段, 黑色代表黑暗阶段
2.4 不同光谱下许氏平鲉血清褪黑素含量变化
褪黑素在许氏平鲉血清中表现出的水平变化为: 绿光组中, 各组间无显著性差异, 在ZT8-ZT16降低, ZT16-ZT24保持缓慢升高趋势, ZT24-ZT4降低。在光周期阶段保持缓慢升高趋势(图8a)。蓝光组在ZT8-ZT4缓慢升高, ZT4达到高峰后激素含量降低(图8b)。黄光组在ZT8-ZT12激素含量缓慢降低(图8c), 并随后在ZT12-ZT24显著升高(<0.05)随后降低。红光组中, 在光周期阶段ZT8-ZT12激素含量下降, 随后缓慢升高, 在ZT24达到峰值, 黑暗阶段显著升高(<0.05), ZT24-ZT4激素含量缓慢降低(图8d)。白光组中, 褪黑素含量在光照阶段ZT8-ZT16缓慢降低, ZT16-ZT24时激素含量显著升高(<0.05), ZT24-ZT8缓慢降低(图8e)。与其他光谱组相比, 蓝光组峰值相位右移至ZT4, 各光谱在夜间褪黑素分泌量较多, 但差异不显著。绿、蓝、红光组平均分泌量显著高于白、黄光组(图8f)。
注: 统计时间点之间的差异(<0.05)由不同的字母符号(a, b, c)表示。图片上方的条状图为光周期, 白色代表光照阶段, 黑色代表黑暗阶段
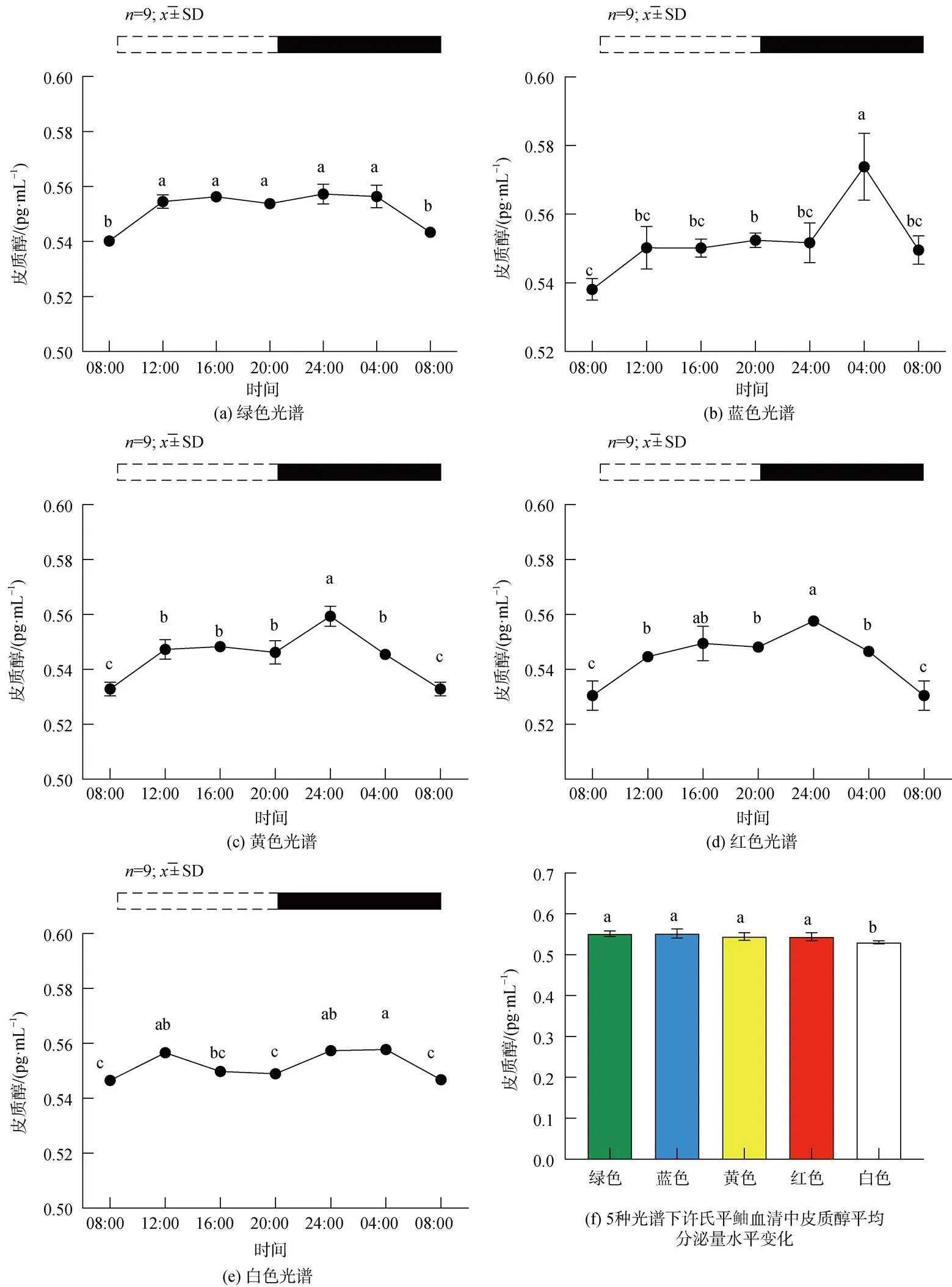
图7 5种光谱下许氏平鲉血清中皮质醇含量的日变化及平均分泌量水平变化
注: 统计时间点之间的差异(<0.05)由不同的字母符号(a, b, c)表示。图片上方的条状图为光周期, 白色代表光照阶段, 黑色代表黑暗阶段
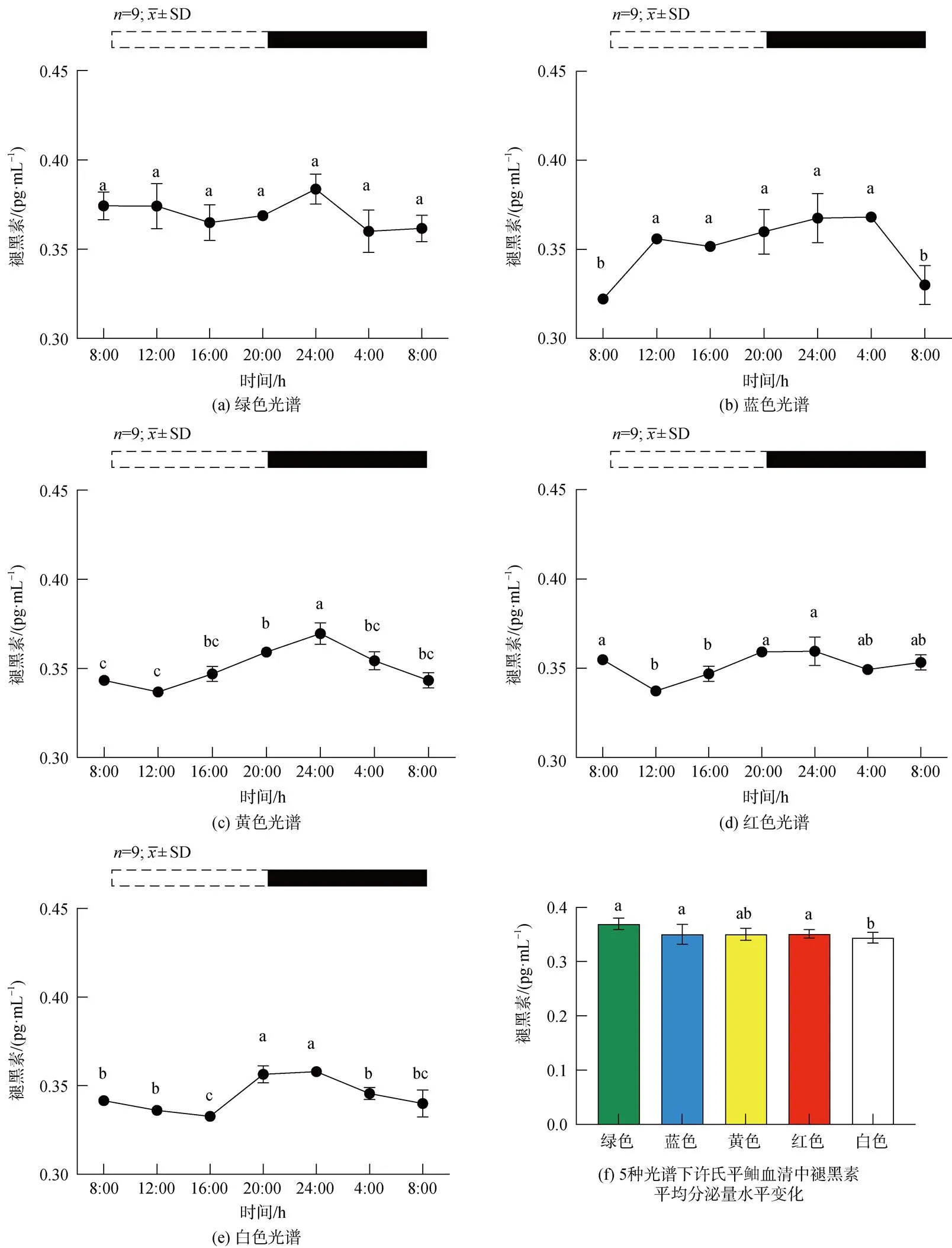
图8 5种光谱下许氏平鲉血清中褪黑素含量的日变化及平均分泌量水平变化
注: 统计时间点之间的差异(<0.05)由不同的字母符号(a, b, c)表示。图片上方的条状图为光周期, 白色代表光照阶段, 黑色代表黑暗阶段
3 讨论
3.1 不同LED光谱对许氏平鲉消化节律的影响
消化酶是一类具有催化活性和高度专一性的特殊蛋白质, 主要由消化腺和消化系统分泌, 其活性高低决定对营养物质消化吸收的能力。胰蛋白酶、脂肪酶与α-淀粉酶作为生物体消化吸收的重要酶类, 标志着对所进食食物的转化能力, 转化能力越高, 越有利于消化吸收, 进而影响生物体的生长发育。影响消化酶活性的因素很多, 如食性[18]、温度[19]、pH值[20-22]、盐度[23]、饲料成分[24-26]、驯饲方式[27]、环境因子[28]等。光作为最重要的环境因子之一, 会对水生生物产生一定的影响。有关光环境对水生生物消化酶活性的影响研究也有部分展开, 有研究发现, 鮸鱼) 幼鱼的脂肪酶活性随着光周期的延长而显著增加, 在18L: 6D和24L: 0D时明显高于12L: 12D[29]。红光环境下可以显著降低斑点鲈()的胰蛋白酶活性[30]。但是消化酶在不同光环境下昼夜节律的变化尚不清楚。本实验通过研究不同光谱下许氏平鲉消化酶活性的昼夜节律, 结果表明, 绿光下的许氏平鲉在ZT8出现胰蛋白酶活性峰值, 在ZT16出现脂肪酶活性峰值, 在ZT24出现淀粉酶活性峰值, 而蓝光条件下于ZT24、ZT20依次出现淀粉酶、胰蛋白酶、脂肪酸酶活力峰。黄、白、红光于ZT16出现消化酶活力峰。绿、蓝光下的许氏平鲉在夜间淀粉酶活性较高, 出现夜高昼低现象。而黄、红光则呈现昼高夜低现象, 这说明蓝、绿光环境下消化高峰期可能在ZT20-ZT24, 出现在夜间, 而黄、红光下的摄食高峰期可能与白光一样, 出现在ZT16左右。曹香林[31]等人研究表明, 草鱼的摄食高峰出现在ZT17左右, 在这个时段蛋白酶、淀粉酶和脂肪酶活性较高, 与本研究在黄、红光下研究结果类似。从实验结果可以看出, 在五种光谱环境中, 红、黄光会影响消化酶活性峰值提前出现, 消化节律发生改变。本实验还发现光、黄光下淀粉酶、脂肪酶的峰值相位左移, 蓝光、绿光下消化酶的峰值相位均右移; 胰蛋白酶无表达的时间差异性。有可能的原因是许氏平鲉的投喂存在时间差, 不同光谱下消化时间会略有不同。同时, 营养素存在消化时序, 消化酶活性与之相适应, 出现节律性。另外, 许氏平鲉消化酶活性的昼夜变化可能受激素的调节, 不同种类酶的调节机制不一定相同, 至于激素的种类、分布以及调节作用, 有待于从细胞水平和分子生物学水平进行更深入的研究。
3.2 不同LED光谱对许氏平鲉代谢节律的影响
丙酮酸激酶和己糖激酶作为糖酵解中的关键限速酶, 其活性的变化在糖代谢的调节中具有重要作用, 反映了糖酵解的水平[32]。乳酸脱氢酶在生物体中作为能量代谢参与糖酵解的一种重要的酶。可催化乳酸和氧化性辅酶I转变为丙酮酸和还原性辅酶I, 进而参与机体的能量代谢。其活性的改变直接影响机体的能量代谢[33]。代谢酶活性作为反映鱼类代谢机能的重要指标, 在不同光谱下有着不同的影响。有研究发现在蓝光和绿光下, 皱纹盘鲍()肝胰脏中己糖激酶和丙酮酸激酶的活性较高, 表明在这些光照条件下有较高的糖酵解速率[34]; 刘松涛等人研究发现绿光组下的红鳍东方鲀()己糖激酶、丙酮酸激酶活性较高, 与蓝、黄、红、白光相比生长发育较好[35]。在本实验中, 一天24 h内, 不同光谱下许氏平鲉代谢酶活性均具有明显的昼夜节律, 绿光于08: 00同时出现己糖激酶、丙酮酸激酶活性峰值, 蓝光条件下于ZT8、ZT16依次出现己糖激酶、丙酮酸激酶、乳酸脱氢酶活力峰。黄光下各类代谢酶分别出现在ZT8、ZT12和ZT20。红光于ZT8、 ZT16、 ZT24出现代谢酶活性峰, 白光在ZT8、ZT12和ZT16依次出现代谢酶活性峰值。许氏平鲉各光谱组的己糖激酶活性均出现昼高夜低现象, 从本实验结果可以看出, 在五种光谱环境中, 绿、蓝光影响代谢酶类活性峰值提前出现, 这说明绿、蓝光环境改变了代谢节律。乳酸脱氢酶活性在绿、黄、红光组下峰值相位左移, 蓝光下峰值相位右移, 丙酮酸激酶活性在绿、蓝、红光组下峰值相位左移。在绿光下各代谢酶的活性峰值相位均会提前出现, 有研究表明丙酮酸激酶、己糖激酶活性的改变会影响ATP水平, 影响肝脏的能量供给, 进而影响糖酵解能力[36-37], 不同光谱影响了许氏平鲉的糖酵解能力, 峰值时间的变化意味在不同光谱下代谢酶产生不同的影响, 原因有可能是不同光谱扰乱了许氏平鲉的代谢节律。
3.3 不同LED光谱对许氏平鲉外周激素的影响
生理节律是受一系列与节律相关激素和基因构成的调控网络来进行调控的, 通过研究相应激素(褪黑素、皮质醇)的时空表达模式对于了解生物钟系统的调控机制与作用规律至关重要。褪黑素作为脊椎动物生物钟系统的最重要输出信号, 能够把产生的节律信息传递到各个组织。它主要由松果体分泌产生[38]。同时, 褪黑素还具有多种功能, 主要功能是调节昼夜节律和季节节律[39], 此外还可以调节神经内分泌因子和抗氧化系统[40-41], 影响鱼类行为、增加或减少运动和摄食活动[42]。相关研究证明, 褪黑素的合成及释放受光调节, 黑暗可刺激褪黑素合成及释放, 光线可抑制褪黑素合成及释放[43]。相反, 皮质醇浓度在睡眠阶段显著升高, 日间活动期间持续降低[44-45]。水生生物体内的皮质醇参与许多过程, 例如应激反应、代谢、渗透调节、行为、生长和繁殖[46]。此前已经报道了许多鱼类皮质醇的每日变化[47-50]。此外, 褪黑素水平受应激和皮质醇的影响, 其影响似乎与应激类型有关。例如, 高饲养密度增加了鲷鱼体内的皮质醇和褪黑素水平[50]; 暴露于急性干扰应激的虹鳟的血浆褪黑素下降[1]。从本实验的结果中可以观察到, 许氏平鲉血清的褪黑素水平均在光照阶段显著降低, 黑暗阶段显著上升, 在尼罗河罗非鱼 () 和非洲鲶鱼()中有相似的研究结果[51], 就各光谱组平均分泌量来看, 白光、黄光组与绿光、蓝光、红光组平均分泌量差异显著; 皮质醇在早上含量高, 白天降低, 并在午夜显著升高至最高值后又开始持续降低。正如在其他昼间动物如人类和灌丛八齿鼠()[52-53]中观察到的一样, 皮质醇的节律在暗期到光照的过渡期显示出峰值。两种激素所测得的昼夜节律性与正常自然节律大致相同[54-55], 且大部分最低值都出现在ZT12-ZT16。因此, 推测许氏平鲉的生理性暗周期可能在ZT16(16: 00)开始, 而生理性光周期则可能在ZT24(24: 00)开始, 对于这一观点还需进一步地研究论证。
4 结论
在长期的进化过程中, 每种生物都形成了自身的昼夜节律, 现在关于这方面的研究还处于种属水平的资料累积阶段。在本实验中, 通过对许氏平鲉褪黑素、皮质醇和消化酶、代谢酶的昼夜变化规律进行初步的分析发现: 1)消化酶活性的昼夜节律均受光谱的影响。红光、黄光下许氏平鲉淀粉酶、脂肪酶活性的峰值相位左移; 胰蛋白酶的活性无时间差异性; 2)代谢酶活性的昼夜节律受光谱影响, 绿、黄及红光下乳酸脱氢酶活性水平的峰值相位发生左移; 丙酮酸激酶的活性在绿、蓝及红光下峰值相位发生左移; 绿、蓝、黄光下己糖激酶活性的峰值相位左移; 3)皮质醇、褪黑素的昼夜节律性与正常自然节律大致相同。综上所述, 在五种光谱环境中, 红、黄光会影响消化酶活性峰值提前出现, 绿、蓝光影响代谢酶类活性峰值提前出现, 光谱可以影响动物生理节律的峰值相位, 不合理的光环境也会扰乱其生理节律。以上结果为鱼类生物钟调节机制的进一步研究和鱼类生理学的研究提供一定价值的基础资料。
[1] MANCERA J M, VARGAS-CHACOFF L, A GARCIA- LOPEZ, et al. High density and food deprivation affect arginine vasotocin, isotocin and melatonin in gilthead sea bream ()[J]. Comparative Biochemistry and Physiology Part A: Molecular & Integrative Physiology, 2008, 149(1): 92-97.
[2] 穆迎春, 王芳, 董双林, 等. 去眼柄对凡纳滨对虾稚虾耗氧率昼夜节律的影响[J]. 海洋湖沼通报, 2004(3): 42-46. MU Yingchun, WANG Fang, DONG Shuanglin, et al. Effects of eyestalk ablation on the diurnal rhythm of oxygen consumption of juvenile[J]. Transactions of Oceanology and Limnology, 2004(3): 42-46.
[3] PIERCE L X, NOCHE R R, PONOMAREVA O, et al. Novel functions for Period 3 and Exorhodopsin in rhythmic transcription and melatonin biosynthesis within the zebrafish pineal organ[J]. Brain Research, 2008, 1223: 11-24.
[4] BISWAS A K, SEOKA M, TANAKA Y, et al. Effect of photoperiod manipulation on the growth performance and stress response of juvenile red sea bream ()[J]. Aquaculture, 2006, 258(1): 350-356.
[5] IMSLAND A K, GUNNARSSON S, ROTH B, et al. Long-term effect of photoperiod manipulation on growth, maturation and flesh quality in turbot[J]. Aquaculture, 2013, 416/417: 152-160.
[6] BALLAGH D A, PANKHURST P M, FIELDER D S. Photoperiod and feeding interval requirements of juvenile mulloway,[J]. Aquaculture, 2008, 277(1/2): 52-57.
[7] 袁满, 王鹏飞, 闫路路, 等. 花鲈垂体和下丘脑中生物钟基因在3种光周期下的表达节律分析[J]. 南方水产科学, 2020, 16(6): 39-46. YUAN Man, WANG Pengfei, YAN Lulu, et al. Circadian rhythmicity of clock genes in pituitary and hypothalamus of spotted sea perch () under three photoperiod conditions[J]. South China Fisheries Science, 2020, 16(6): 39-46.
[8] 孙佳敏, 王芳, 张立斌, 等. 不同光照周期对刺参摄食节律和消化生理的影响[J]. 海洋科学, 2015, 39(5): 1-8. SUN Jiamin, WANG Fang, ZHANG Libin, et al. Effects of photoperiods on feeding rhythm and digestive physiology of sea cucumber,[J]. Marine Sciences, 2015, 39(5): 1-8.
[9] 王晓杰, 张秀梅, 李文涛. 盐度胁迫对许氏平鲉血液免疫酶活力的影响[J]. 海洋水产研究, 2005, 26(6): 17-21. WANG Xiaojie, ZHANG Xiumei, LI Wentao. Effects of salinity on the non-specific immuno-enzymetic activity of[J]. Progress in Fishery Sciences, 2005, 26(6): 17-21.
[10] 魏平平, 李鑫, 刘鹰, 等. 光周期对红鳍东方鲀脑组织中GH和SS基因表达水平和昼夜表达模式的影响[J]. 大连海洋大学报, 2020, 35(1): 108-113. WEI Pingping, LI Xin, LIU Ying, et al. Effects of photoperiod on expression level and daily expression pattern of GH and SS genes in brain of tiger puffer[J]. Journal of Dalian Ocean University, 2020, 35(1): 108-113.
[11] WHITMORE D, FOULKES N S, STRAHLE U, et al. Zebrafish Clock rhythmic expression reveals independent peripheral circadian oscillators[J]. Nature Neuroscience, 1998, 1(8): 701-707.
[12] CUESTA I H, LAHIRI K, LOPEZ-OLMEDA J F, et al. Differential maturation of rhythmic clock gene expression during early development in medaka ()[J]. Chronobiol International, 2014, 31(4): 468-478.
[13] SANCHEZ J A, MADRID J A, SANCHEZ-VAZQUEZ F J. Molecular cloning, tissue distribution, and daily rhythms of expression of per1 gene in European sea bass ()[J]. Chronobiology International, 2010, 27(1): 19-33.
[14] PATINO M A, RODRIGUEZ-ILLAMOLA A, CONDE- SIEIRA M, et al. Daily rhythmic expression patterns of clock1a, bmal1, and per1 genes in retina and hypothalamus of the rainbow trout,[J]. Chronobiology International, 2011, 28(5): 381-389.
[15] HUANG T S, RUOFF P, FJELLDAL P G. Effect of continuous light on daily levels of plasma melatonin and cortisol and expression of clock genes in pineal gland, brain, and liver in Atlantic salmon postsmolts[J]. Chronobiology International, 2010, 27(9/10): 1715- 1734.
[16] LAZADO C C, NAGASAWA K, BABIAK I, et al. Circadian rhythmicity and photic plasticity of myosin gene transcription in fast skeletal muscle of Atlantic cod ()[J]. Marine Genomics, 2014, 18: 21-29.
[17] 魏平平, 李鑫, 费凡等. 光谱对红鳍东方鲀仔稚鱼生长及相关基因表达量的影响[J]. 大连海洋大学学报, 2019, 34(5): 668-673. WEI Pingping, LI Xin, FEI Fan, et al. Effects of light spectrum on growth and related gene expression of larval and juvenile tiger puffer[J]. Journal of Dalian Ocean University, 2019, 34(5): 668-673.
[18] JONAS E, RAGYANSZKI M, OLAH J, et al. Proteolytic digestive enzymes of carnivorous (L.), herbivorous (Val.) and omnivorous (L.) fishes[J]. Aquaculture, 1983, 30(1/4): 145-154.
[19] 方之平, 潘黔生, 何瑞国, 等. 温度对彭泽鲫主要消化酶活力的影响[J]. 水利渔业, 1998 (2): 15-17, 47.
[20] 沈文英, 寿建昕, 祝尧荣, 等. 银鲫消化酶活性与pH值的关系[J]. 水产科学, 2002, 21(6): 10-12.SHEN Wenying, SHOU Jianxin, ZHU Yaorong, et al. The effect of pH on activity of digestive enzymes of[J]. Fisheries Science, 2002, 21(6): 10-12.
[21] 伍莉, 陈鹏飞. pH值对黄鳝肠道和肝胰脏主要消化酶活力的影响[J]. 饲料工业, 2002, 23(8): 40-41. WU Li, CHEN Pengfei. The Impact of temperature on the activity of digestion enzymes in[J]. Feed Industry, 2002, 23(8): 40-41.
[22] 叶元土, 林仕梅, 罗莉, 等. 温度、pH值对南方大口鲶、长吻鮠蛋白酶和淀粉酶活力的影响[J]. 大连水产学院学报, 1998, 13(2): 19-25. YE Yuantu, LIN Shimei, LUO Li, et al. The effect of temperature and pH value on activiies of proteases and amylases fromand[J]. Journal of Dalian Fisheries University, 1998, 13(2): 19-25.
[23] 李培伦, 刘伟, 王继隆, 等. 盐度对养殖大麻哈鱼血液生化指标及消化酶活力的影响[J]. 西北农林科技大学学报(自然科学版), 2020, 48(9): 10-16. LI Peilun, LIU Wei, WANG Jilong, et al. Effects of salinity on blood biochemical parameters and digestive enzyme activities of farmed chum salmon ()[J]. Journal of Northwest A & F University(Natural Science Edition), 2020, 48(9): 10-16.
[24] 张家国, 王义强, 邹师哲. 不同蛋白质能量比饲料与夏花草鱼消化酶的关系[J]. 上海海洋大学学报, 1997, 6(1): 54-58. ZHANG Jiaguo, WANG Yiqiang, ZOU Shizhe. The relationship of dietary protein/energy ratios and activities of digestive enzyme of juvenile grass carp[J]. Journal of Shanghai Ocean University, 1997, 6(1): 54-58.
[25] 王红权, 孙桂芳, 赵玉蓉. 异育银鲫摄食5种不同动物蛋白源饲料后消化酶活性变动比较[J]. 内陆水产, 2000, 25(2): 9-11. WANG Hongquan, SUN Guifang, ZHAO Yurong. Comparison of changes in digestive enzyme activities after feeding 5 different animal protein sources in allotrophic silver crucian carp[J]. Current Fisheries, 2000, 25(2): 9-11.
[26] 田丽霞, 林鼎. 草鱼摄食两种蛋白质饲料后消化酶活性变动比较[J]. 水生生物学报, 1993, 17(1): 58-65. TIAN Lixia, LIN Ding. Changes in digestive enzyme activities in grass crapafter feeding of two test diets[J]. Acta Hydrobiologica Sinica, 1993, 17(1): 58-65.
[27] 钱国英. 不同驯食方式对鳜鱼胃肠道消化酶活性的影响[J]. 浙江农业大学学报, 1998, 24(2): 99-102. QIAN Guoying. Change of digestive enzyme activities in intestinal canal of domesticated mandarin fish[J]. Journal of Zhejiang University (Agriculture and Life Sciences), 1998, 24(2): 99-102.
[28] SINGH A, ZUTSHI B. Photoperiodic effects on somatic growth and gonadal maturation in Mickey Mouse platy,(Gunther, 1866)[J]. Fish Physiology and Biochemistry, 2020, 46(4): 1483-1495.
[29] SHAN X, XIAO Z, HUANG W, et al. Effects of photoperiod on growth, mortality and digestive enzymes in miiuy croaker larvae and juveniles[J]. Aquaculture, 2008, 281(1/4): 70-76.
[30] HOU Z S, WEN H S, LI J F, et al. Effects of photoperiod and light Spectrum on growth performance, digestive enzymes, hepatic biochemistry and peripheral hormones in spotted sea bass ()[J]. Aquaculture, 2019, 507: 419-427.
[31] 曹香林, 郭蓓, 彭墨, 等. 不同发育阶段草鱼消化酶活力的变化及其昼夜节律[J]. 河南农业科学, 2009, 38(7): 120-123. CAO Xianglin, GUO Bei, PENG Mo, et al. Circadian rhythms of digestive enzyme activities ofof different developmental stages[J]. Journal of Henan Agricultural Sciences, 2009, 38(7): 120-123.
[32] 陈婉情. 5种光色对豹纹鳃棘鲈幼鱼生长特征及生理生化功能的影响[D]. 上海: 上海海洋大学, 2016. CHEN Wanqing. The influences of light color on growth, physiological and biochemical indices of juvenile[D]. Shanghai: Shanghai Ocean University, 2016.
[33] ZAKHARTSEV M, JOHANSEN T, PORTNER H O, et al. Effects of temperature acclimation on lactate dehydrogenase of cod (): genetic, kinetic and thermodynamic aspects[J]. The Journal of Experimental Biology, 2004, 207(Pt1): 95-112.
[34] GAO X L, ZHANG M, LI X, et al. Effects of LED light quality on the growth, metabolism, and energy budgets of[J]. Aquaculture, 2016, 453: 31-39.
[35] 刘松涛, 李伊晗, 李鑫, 等.不同LED光谱对红鳍东方鲀幼鱼生长、摄食及消化酶活性的影响[J]. 中国水产科学, 2021, 28(8): 1011-1019. LIU Songtao, LI Yihan, LI Xin, et al. Effects of LED spectra on growth, feeding, and digestive enzyme activities of juvenile[J]. Journal of Fishery Sciences of China, 2021, 28(8): 1011-1019.
[36] MARQUEZE A, KUCHARSKI L C, SILVA R S M D. Effects of anoxia and post-anoxia recovery on carbohydrate metabolism in the jaw muscle of the crabmaintained on carbohydrate-rich or high-protein diets[J]. Journal of Experimental Marine Biology and Ecology, 2006, 332(2): 198-205.
[37] LAIZ-CARRION R, SANGIAO-ALVARELLOS S, GUZMAN J M, et al. Energy metabolism in fish tissues related to osmoregulation and cortisol action[J]. Fish Physiology and Biochemistry, 2002, 27(3/4): 179-188.
[38] BORNESTAF C, MAYER I, BORG B. Melatonin and maturation pace in female three-spined stickleback,[J]. General and Comparative Endocrinology, 2001, 122(3): 341-348.
[39] FALCON J. Cellular circadian clocks in the pineal[J]. Progress in Neurobiology, 1999, 58(2): 121-162.
[40] FALCON J, BESSEAU L, SAUZET S, et al. Melatonin effects on the hypothalamo-pituitary axis in fish[J]. Trends in Endocrinology & Metabolism, 2007, 18(2): 81-88.
[41] HARDELAND R, COTO-MONTES A, POEGGELER B. Circadian rhythms, oxidative stress, and antioxidative defense mechanisms[J]. Chronobiology international 2003, 20(6): 921-962.
[42] LOPEZ-OLMEDA J F, MADRID J A, SANCHEZ- VAZQUEZ F J. Melatonin effects on food intake and activity rhythms in two fish species with different activity patterns: Diurnal (goldfish) and nocturnal(tench)[J]. Comparative Biochemistry and Physiology Part A: Molecular & Integrative Physiology, 2006, 144(2): 180-187.
[43] AMANO M, IIGO M, IKUTA K, et al. Roles of melatonin in gonadal maturation of underyearling precocious male masu salmon[J]. General and Comparative Endocrinology, 2000, 120(2): 190-197.
[44] 费广鹤. 褪黑素和皮质醇水平的昼夜节律性变化及其mt1受体在中枢神经系统中的表达与支气管哮喘[D]. 合肥: 中国科学技术大学, 2003. FEI Guanghe. Alterations in circadian rhythms of melatonin and cortisol, and expression of mt1 receptor in the central nervous with asthma[D]. Hefei: University of Science and Technology of China, 2003.
[45] VELDHUIS J D, IRANMANESH A, JOHNSON M L, et al. Amplitude, but not frequency, modulation of adrenocorticotropin secretory bursts given rise to the nyctohemeral rhythm of the corticotrophic axis in man[J]. The Journal of Clinical Endocrinology and Metabolism, 1990, 71(2): 452-463.
[46] MOMMSEN T P, VIJAYAN M M, MOON T W. Cortisol in teleosts: dynamics, mechanisms of action, and metabolic regulation[J]. Reviews in Fish Biology and Fisheries, 1999, 9(3): 211-268.
[47] JM CERDA-REVERTER, ZANUY S, CARRILLO M, et al. Time-course studies on plasma glucose, insulin, and cortisol in sea bass () held under different photoperiodic regimes[J]. Physiology & Behavior, 1998, 64(3): 245-250.
[48] EBBESSON L, BJORNSSON B T, EKSTROM P, et al. Daily endocrine profiles in parr and smolt Atlantic salmon[J]. Comparative Biochemistry and Physiology Part A: Molecular & Integrative Physiology, 2008, 151(4): 698-704.
[49] LI Y Y, TAKEI Y. Ambient salinity-dependent effects of homologous natriuretic peptides (ANP, VNP, and CNP) on plasma cortisol level in the eel[J]. General and Comparative Endocrinology, 2003, 130(3): 317-323.
[50] POLAKOF S, CEINOS R M, FERNANDEZ-DURAN B, et al. Daily changes in parameters of energy metabolism in brain of rainbow trout: Dependence on feeding[J]. Comparative Biochemistry and Physiology Part A: Molecular & Integrative Physiology, 2007, 146(2): 265-273.
[51] MARTINEZ-CHAVEZ C C, AL-KHAMEES S, CAMPOS- MENDOZA A, et al. Clock-controlled endogenous melatonin rhythms in Nile tilapia () and African catfish ()[J]. Chronobiology International, 2008, 25(1): 31-49.
[52] BAILEY S L, HEITKEMPER M M. et al. Circadian rhythmicity of cortisol and body temperature: Morningness-eveningness effects[J]. Chronobiology international, 2001, 18(2): 249-261.
[53] MOHAWK J A. Restraint stress delays reentrainment in male and female diurnal and nocturnal rodents[J]. Journal of Biological Rhythms, 2005, 20(3): 245-256.
[54] WHITSON P A, PUTCHA L, CHEN Y M, et al. Melatonin and cortisoI assessment of circadian shifts in astronauts before flight[J]. Journal of Pineal Research, 1995, 18(3): 141-147.
[55] KANEMATSU N, HONMA S, KATSUNO Y, et al. lmmediate response to light of rat pineal melatonin rhythm: analysis by in vivo microdialysis[J]. The American Journal of Physiology, 1994, 266(6): 1849-1855.
Effects of spectra on digestion, metabolic rhythm, and physiological stress of
FANG Ying-ying1, 2, CHEN Lu-lu1, 2, LIU Song-tao1, 2, SUN Fei1, 2, WU Yan-ling1, 2, LI Xin1, 2, LIU Ying1, 3, MA He1, 2
(1. College of Marine Technology and Environment, Dalian Ocean University, Dalian 116023, China; 2. Key Laboratory of Environment Controlled Aquaculture (KLECA), Dalian 116023, China; 3. College of Biosystems Engineering and Food Science, Zhejiang University, Hangzhou 310058, China)
The serum, intestinal tract, and liver ofwere sampled over a period of 24 h (8: 00, 12: 00, 16: 00, 20: 00, and 24: 00) under five spectra (red, green, yellow, blue, and white lights, respectively; photoperiod 12L: 12D). to investigate the digestion and metabolism levels and peripheral hormone rhythms. Results revealed that the digestion and metabolism enzymes ofunder different spectra exhibited rhythmic patterns, except for trypsin. The peak phases of α-amylase (α-AMS) and lipase (LPS) under yellow light shifted to the left compared to those under white light, and trypsin (TRY) showed no temporal difference. The peak phase of Lactate dehydrogenase (LDH) under green, yellow, and red lights moved to the left compared to that under white light. In the five spectral environments, red and yellow light affected the peak of digestive enzyme activity earlier. The peak phase of pyruvate kinase (PK) shifted to the left under green, blue, and red lights. The secretion of hexokinase (HK) was observed under green, blue, and yellow lights, and its peak phase shifted to the left compared to that under white light. In five spectral environments, green and blue lights affected the peak of metabolic enzymes substantially. The melatonin level in the serum ofwas low during the day and high at night. Moreover, cortisol decreased during the day and began to diminish continuously after reaching its highest value at midnight. Changes in the peak phase indicate variations in circadian rhythms. The above results show that different spectra can influence the physiological metabolic rhythm of fish. Thus, future research should fully consider the biological effect of spectra on cultured organisms and then formulate appropriate lighting conditions.
; spectra; circadian rhythm; digestion and metabolism; stress
Aug. 19, 2022
[National Natural Science Foundation of China, No.32202961; Science & Technology Program of Liaoning Province, No. 2021JH2/10200011; Open Project of Key Laboratory of Environment Controlled Aquaculture (Dalian Ocean University) Ministry of Education, No. 202202; Modern Agro-industry Technology Research System, No. CARS-49]
S9645.3
A
1000-3096(2023)10-0094-18
10.11759/hykx20220819003
2022-08-19;
2022-11-09
国家自然基金(32202961); 辽宁省科学技术计划项目(2021JH2/10200011); 设施渔业教育重点实验开放课题(202202); 现代农业产业技术体系专项资金(CARS-49)
房莹莹(1997—), 女, 山东济南人, 硕士研究生, 研究方向为海洋科学, E-mail: taylorfang1997@163.com;马贺(1986—),通信作者, 女, 辽宁大连人, 讲师, 研究方向为光环境与水生生物互作机理, E-mail: mahe@dlou.edu.cn
(本文编辑: 杨 悦)
猜你喜欢
当代水产(2021年9期)2021-12-02
当代水产(2021年8期)2021-11-04
Plasma Science and Technology(2021年8期)2021-08-05
中国美容医学(2021年5期)2021-06-22
水产养殖(2021年5期)2021-05-21
医学前沿(2021年18期)2021-04-14
保健与生活(2018年17期)2018-01-27
作文与考试·小学低年级版(2016年10期)2016-09-10
中国民族民间医药·下半月(2014年4期)2014-09-26
中国海洋大学学报(自然科学版)(2014年12期)2014-02-28
