
珍稀濒危植物海南粗榧群落树种组成与种间联结性
2024-01-22 12:14王如农寿千彭文成吴彪杨佳廖立国
生态环境学报 2023年10期
王如,农寿千,彭文成,吴彪,杨佳,廖立国
海南省林业科学研究院(海南省红树林研究院),海南 海口 571100
植物群落中的物种组成与区系分布是认识群落变化、发展的基础(Tilman et al.,2006)。种间联结反映了植物种群在空间分布上的相互关联性,是用于描述在某个时间范围内不同物种由于生境差异而形成的相互关系,对于研究植物的种间关系,维护群落的稳定性具有重要意义(Akatov et al.,2018)。濒危植物往往受限于自身的生物学与生态学特性,天然更新繁衍速度慢,因此抵御风险能力较弱,在遭到人类不合理的开发利用时容易面临灭绝的风险,导致生态系统生物多样性的丧失(De Sequeira et al.,2021;叶兴状等,2021)。研究濒危植物所在群落物种组成与种间联结特征,可揭示其种群生存状况和群落演替趋势,有助于认识物种的濒危机制,探究群落物种间的竞争或互利共生关系,为制定具有针对性的濒危植物保护策略提供科学理论基础(潘元琪等,2020)。国内众多专家学者近年来已对不同地区的珍稀濒危保护植物开展了关于种间联结性的研究,王博等(2017)对甘肃省安西保护区的裸果木(Gymnocarposprzewalskii)的调查结果表明,裸果木趋于独立生长,与大多数物种联结不显著,群落处于不稳定状态;蔡鑫等(2019)通过野外踏查浙江华顶杜鹃(Rhododendron huadingense)分布区,发现华顶杜鹃的群落种间关系松散,整体关联不显著;罗敏贤等(2022)以福建龟山的伞花木(Eurycorymbuscavaleriei)为研究对象,发现伞花木与群落优势种的种间联结性低,需要进行适当人工干预。这些研究揭示了不同濒危保护植物对生境的适应性及其群落物种间的动态联系,对于濒危物种的多样性保护至关重要。
海南粗榧(Cephalotaxushainanensis)是三尖杉科(Cephalotxaceae)三尖杉属(Cephalotaxus)的常绿高大乔木,主要分布在东亚和中南半岛北部,在国内主要分布在海南热带雨林国家公园的尖峰岭、吊罗山、五指山、霸王岭等海拔在700—1 100 m 的山地雨林沟谷两侧中,零星分布在广东、广西、西藏南部等地(符文英等,2003)。由于其种群数量稀少,种子萌发困难,且因能从其树皮及枝叶提取具有治疗白血病和急性淋巴病的三尖杉酯碱类药物而遭到大规模砍伐,现已被列入国家二级重点保护植物(祁珊珊等,2010)。目前已有许多学者对海南粗榧的遗传多样性(Du et al.,2002)、育种栽培(曾祥全等,2016)、表型结构(向志强等,2002)、药物合成(Qiao et al.,2023)等方面开展了较为深入的研究,但对其群落组成及种间联结性研究还未见报道。本团队以海南热带雨林国家公园内的野生海南粗榧群落为研究对象,通过对其分布点的样方调查,分析了海南粗榧群落的物种组成与植物区系特征,探讨了群落常见树种间的联结性,以期为海南粗榧濒危机制的诠释、野生种群的保护与恢复提供科学的理论支撑。
1 研究区概况和研究方法
1.1 研究区概况
海南热带雨林国家公园地处海南岛中南部山区(18°33′16″—19°14′16″N,108°44′32″—110°04′43″E),东起吊罗山国家森林公园,西至尖峰岭国家级自然保护区,南至保亭黎族苗族自治县毛感乡,北至黎母山省级自然保护区,范围涉及海南岛中部9 个市县,总面积达到4 269 km2。国家公园位于热带北缘,受印度洋季风、西北太平洋季风以及东亚季风三大季风影响,形成了热带海洋性季风气候,干湿季分明(姜超等,2017),光照时间充足,年均气温在22.5—26.0 ℃之间,多年平均降雨量为1 759 mm,是海南三大江(南渡江、昌化江、万泉河)的发源地,有海南“水塔”之称。园区的森林资源丰富,覆盖率达到95.6%,其中天然林面积为3 294.36 km2,占国家公园面积的 74.9%,常见植物有楝科(Meliaceae)、樟科(Lauraceae)、茜草科(Rubiaceae)、锦葵科(Malvaceae)等;人工林面积824 km2,主要以橡胶(Heveabrasiliensis)、杉木(Cunninghamia lanceolata)、马占相思(Acaciamangium)、槟榔(Arecacatechu)等经济、用材林种为主的纯林。植被类型涵盖分布在中部山区海拔800 m 以下的热带低地雨林、面积最大的热带山地雨林、较稀少的热带针叶林及高山云雾林。截止至2019 年,共统计有包括各类保护植物432 种在内的野生维管植物3 653 种(陈宗铸等,2021)。
1.2 研究方法
1.2.1 样地设置及调查
调查工作于2022 年2—4 月进行,根据海南粗榧常散生于海拔700—1 200 m 山地雨林或季雨林的沟谷或溪涧密林山坡的分布特征,在海南热带雨林国家公园林区保护人员的指导下,对生长有野生海南粗榧且人为干扰极小的霸王岭、尖峰岭、黎母山、五指山原始森林地段进行调查,以调查发现的8 株海南粗榧母树为中心,设置8 个20 m×20 m 标准样地(表1),记录各样方的海拔、经纬度、坡度等数据,同时对样地内的胸径大于1 cm 且树高大于1.5 m 的乔木树种进行每木检尺,记录树高、冠幅、胸径、枝下高、树种名等指标。

表1 海南粗榧样方地理位置及母树信息Table 1 Geographic location and maternal plants information of Cephalotaxus hainanensis sample plot
1.2.2 树种组成与区系分析
通过查阅中国植物志在线资源库(http://www.iplant.cn/frps)及《海南植物图志》(杨小波,2015),对样地内乔木的科、属、种进行分类鉴定及统计分析。根据吴征镒等(2003)提出的世界种子植物科的分布区类型系统,分析群落中树种的区系成分,统计样地内的乔木科的分布区类型。
1.2.3 总体关联性检验
总体关联性采用Schlute(1984)提出的方差比率法(Rv)检验群落乔木物种间的总体联结性,并用统计量W来检验总体联结是否显著,具体公式如下:
式中:
S——总物种数;
ST——所有物种数的方差;
δT——所有物种数出现频度的方差;
N——总样方数;
ni——物种i出现的样方次数;
Pi——物种出现的频度;
Ti——样方i的物种总数;
t——样方的物种平均数。
Rv——独立假设条件下群落物种间的总体联结性指数,若Rv>1,则物种间总体表现为正关联,若Rv<1,则物种间总体呈现负关联,若Rv=1,则物种间总体无关联。
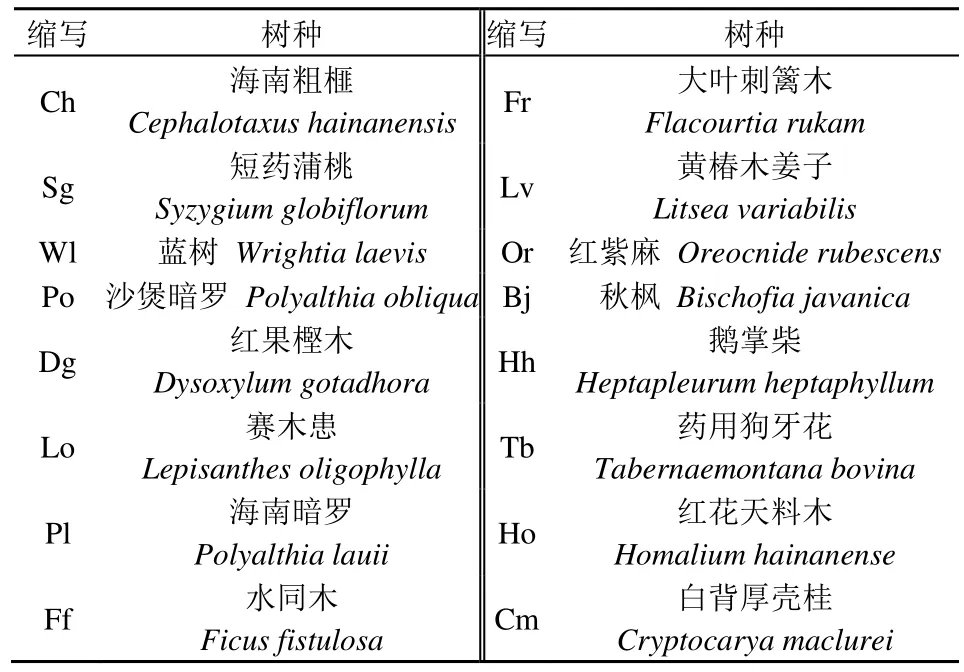
通过统计量W检验Rv值偏离1 的显著程度,W=Rv×N,若W<χ2(0.95,N)或W>χ2(0.05,N),则物种间为显著关联;若χ2(0.95,N) 1.2.4 种间关联性检验 根据样方调查数据中物种的存在与否,建立2×2 列联表,利用V值来判断物种间的正负关联性,依据χ2统计量来表示物种间的关联性,χ2值通过Yates 的连续校正公式(张金屯,2018)计算,公式如下: 式中: ɑ——两个物种都出现的样方数; b和c——两个物种单独出现的样方数; d——两个物种都没出现的样方数。若V>0,则表示种间正关联,当V<0,则表示种间负关联。若χ2<3.841,即P>0.05,代表种间关联不显著;若3.841≤χ2≤6.635,即0.01≤P≤0.05,表明种间联结性显著,当χ2>6.635,即P<0.01,种间联结性为极显著。 联结系数(AC)可以反映种间联结性的关联强度,而共同出现百分率(PC)能较准确判断物种间正联结强度,避免ɑ值和d值偏高或偏低造成的偏差,计算公式如下: AC的值域为[-1, 1],正关联越强越靠近1,负关联越强越靠近-1;PC的值域为[0, 1],反映正关联的紧密程度,值越接近1 正关联越强。 Pearson 相关系数(rp)和Spearman 秩相关系数(rs)通过物种数量反映物种间的线性关系,是分析种间关联程度大小的有效补充和完善(徐满厚等,2016)。 式中: rp和rs的值域为[-1, 1],大于0 为正相关,小于0 为负相关; N——样方数; xik和xjk——物种i和物种j在样方k中的秩; t检验可以检验相关系数的显著性,公式如下: 式中: t——检验量; r——相关系数; n——样本数量即样方数N,计算得到t值后查询对应概率P值。 调查的8 个海南粗榧样地中共发现乔木105种,隶属44 科78 属(表2),其中双子叶植物占绝对优势,为41 科75 属102 种,分别占科、属、种总数的93.2%、96.2%、97.1%,单子叶植物仅为1科1 属1 种,裸子植物为2 科2 属2 种。群落乔木树种类型丰富,科、属的种类较为复杂,5 种乔木以上的科有6 科,共26 属43 种,分别占科、属、种总数的 13.6%、33.3%、41.0%,其中以桑科(Moraceae)4 属10 种及樟科5 属10 种为最,楝科为4 属7 种,茜草科为5 属6 种,夹竹桃科(Apocynaceae)及锦葵科都为4 属5 种,群落乔木在科的层面较为集中,优势科较为明显。群落的78属中,有59 个单种属,占总属数的75.6%,较多种类的为榕属(Ficus)6 种,根据统计结果可看出在属的层面,群落分布较为分散,无明显优势属,体现了热带森林群落组成的复杂性。 表2 海南粗榧伴生群落树种科属统计Table 2 Family and genus statistics of tree species in Cephalotaxus hainanensis concomitant community 海南粗榧样地群落的伴生树种共有9 种分布区类型(表3),在调查发现的44 科植物中,世界分布类型(1)8 科,包括群落种数最多的桑科,还有豆科(Fabaceae)、榆科(Ulmaceae)等。热带分布类型(2—6)最多,达29 科,占总科数的65.9%,其中泛热带分布 21 科, 包括夹竹桃科(Apocynaceae)、棕榈科(Arecaceae)、橄榄科(Burseraceae)等,多为群落常见种和重要标志种;东亚(热带、亚热带)及热带南美间断分布5 科,有冬青科(Aquifoliaceae)、杜英科(Elaeocarpaceae)等,其他热带类型皆为1 科。温带分布类型(7—9)7 科,其中北温带分布4 科,仅占总科数的9.09%,有杨柳科(Salicaceae)、山茱萸科(Cornaceae)等。分析海南粗榧样地群落伴生树种区系可知,热带成分占主要组成部分,温带成分最低,群落区系具有明显的热带性质(杜家贤等,2020)。 表3 海南粗榧伴生群落树种科的分布区类型Table 3 Distribution area types of tree species family in Cephalotaxus hainanensis concomitant community 根据所调查样方物种出现的频度数据,选择频度不小于0.5 的树种(表4)分析种间的联结性(魏亚情等,2022),如表4 所示,共有16 种树种进行分析,经过计算分析,海南粗榧群落常见树种的总体关联性方差比率Rv为 0.534<1,且统计量W=4.27,介于χ20.95(2.733)与χ20.05(15.507)之间,表明总体关联性表现为不显著的负关联,群落常见树种总体上还没形成紧密的耦合关系。 表4 海南粗榧群落常见树种种名及缩写Table 4 Name and abbrevition of common species in Cephalotaxus hainanensis community 2.4.1χ2检验分析 海南粗榧群落常见树种的χ2检验结果显示如下(图1),由于本次调查的特殊性,所有样方均有海南粗榧的存在,因此种间关联分析2×2 列联表中的c、d 值均为0,与海南粗榧相关的AC值与χ2值均无。其余15 种常见树种的105 个种对中,正关联的种对有33 对,负关联41,无关联31,分别占总对数的31.4%、39.1%、29.5%,均不存在显著关联种对,说明海南粗榧群落常见树种的种间关联结构较为松散,联结性弱。 图1 海南粗榧群落常见树种种间关联χ2半矩阵图Figure 1 Semi-matrix of interspecific correction χ2-test of association of common species in Cephalotaxus hainanensis community 2.4.2 关联度分析 通过χ2检验可以判断树种间是否存在显著联结,但仍然需要联结系数(AC)及共同出现百分率(PC)对种对间的极性大小及不显著但仍存在关联的种对进行测定。由图2a 知,海南粗榧群落主要树种种对中,AC>0.67 的种对有9 对,占总对数的8.57%,包括短药蒲桃(Syzygiumglobiflorum)和沙煲暗罗(Polyalthiaobliqua)、黄椿木姜子(Litseavariabilis)、秋枫(Bischofiajavanica)、白背厚壳桂(Cryptocaryamaclurei),蓝树(Wrightialaevis)和药用狗牙花(Tabernaemontanabovina)等种对,正联结系数越大说明这两个树种对环境差异具有相似的响应,所需的生境条件接近;0.33≤AC<0.67 的种对有23 对,占总对数的21.9%;0≤AC<0.33 的种对有28 对,占总对数26.7%;-0.33≤AC<0 的种对有22 对,占总对数的21.0%;-0.67≤AC<0.33 的种对有11 对,占总对数的10.5%;AC<-0.67 的对数有12 对,占总对数的11.4%,系数较小表示树种间独立性较强,联结性弱,而负值的则表示这些树种间存在排斥作用,或者生境需求不同。 图2 海南粗榧群落常见树种种间关联AC值(a)和PC值(b)半矩阵图Figure 2 Semi-matrix of interspecific correction AC value (a), PC value (b) of association of common species in Cephalotaxus hainanensis community 共同出现百分率可以表示不同树种间正联结程度的大小,并在一定程度上克服AC值的偏差,由图2b 可知,PC≥0.75 的种对仅有7 对,占总对数的5.83%,即海南粗榧和短药蒲桃,短药蒲桃和沙煲暗罗,蓝树和海南暗罗(Polyalthialauii)等种对,说明这些种对能较好的和谐相处,利用共同生境资源;0.5≤PC<0.75 的种对有41 对,占总对数的34.2%;0.25≤PC<0.5 的种对有55 对,占总对数的45.8%;PC<0.25 的对数有17 对,占总对数的14.2%,百分率较低说明这些树种间对生境的要求差异较大,共同出现在同一环境的可能性较小。从AC值和PC值的总体大小可看出海南粗榧的群落主要树种的联结呈现较为松散或无联结状态,树种间的独立性较强,与χ2检验的结果基本一致。 2.4.3 相关性分析 种对间的相关分析可进一步对种间联结程度大小进行补充完善,从树种的Pearson 检验结果(图3a)可知,在所有的120 个种对中,正关联种对有57 对,占总对数的47.5%,其中显著正关联种对仅有赛木患和红紫麻;无关联种对3 对,占总对数的2.50%;负关联种对60 对,占总对数的50.0%,其中显著负关联有3 对,包括海南粗榧和药用狗牙花、蓝树和鹅掌柴(Heptapleurumheptaphyllum)、红果樫木(Dysoxylumgotadhora)和大叶刺篱木(Flacourtia rukam),极显著负关联仅有秋枫和白背厚壳桂。 图3 海南粗榧群落常见树种Pearson 相关性和Spearman 秩相关性检验Figure 3 Pearson correlation coefficien and Spearman rank correlation coefficient of common species in Cephalotaxus hainanensis community 种对间的Spearman 秩相关性检验结果显示(图3b),正关联种对有61 对,占总对数的50.8%,其中呈显著正关联的种对 2 对,分别为赛木患(Lepisanthesoligophylla)和大叶刺篱木、赛木患和红紫麻(Oreocniderubescens),极显著正关联的2对,为沙煲暗罗和鹅掌柴、大叶刺篱木和白背厚壳桂;无关联仅1 对,为红花天料木(Homalium hainanense)和白背厚壳桂;负关联种对有58 对,占总对数的48.3%,其中显著负关联种对4 对,包括沙煲暗罗和赛木患、秋枫和白背厚壳桂等,极显著负关联仅有1 对,为海南暗罗和药用狗牙花。整体而言,海南粗榧群落中显著正关联种对相对较少,物种间较为相互独立,而显著负关联的种对明显多于显著正关联。 海南粗榧样地群落的伴生树种调查结果有44 科78 属105 种,其中热带区系植物占总科数的65.9%,具有明显的区域性质(杜家贤等,2020),与同属濒危植物的海南假韶子(Paranepheliumhainanense)群落具有相似的区系特征(罗金环等,2018),但从群落物种组成来看,海南粗榧群落的乔木树种有105种,要显著高于海南假韶子群落的36 种,这可能是因为物种间生长环境受人为干扰的强度差异所导致(王丽等,2019)。分析群落的主要优势科可知,海南粗榧群落主要由桑科、樟科、楝科等树种组成,而海南假韶子群落优势科为大戟科(Euphorbiaceae)、梧桐科(Sterculiaceae)、无患子科(Sapindaceae),这说明不同的热带森林濒危树种在群落树种组成与优势植物都存在明显差异,因此针对不同的濒危植物开展野外抚育和保护工作时需要根据树种间的区别制定不同方案(杨阳等,2019)。 种间联结性与群落的稳定性密切相关,是群落各物种在生境影响下相互作用、影响形成的有机联系,可在一定程度上反映该群落组成物种间的相互关系(黄祥童等,2015)。本研究中的总体关联性结果显示海南粗榧群落主要树种间为不显著的负关联,群落的组成结构还未成熟,稳定性较弱,处于不断发展的动态演替阶段,与同属常绿针叶树种的贵州雷公山自然保护区台湾杉(Taiwania cryptomerioides)群落(王加国等,2015)总体负关联结果相似,而在海南省吊罗山濒危植物石碌含笑集中分布区开展的总体关联性研究结果表现为显著正相关(魏亚情等,2022),说明濒危植物受限自身的生态学和生物学特性以及群落的发展阶段,种群数量较少,在群落中趋于独立生长,而其种群数量的增加可以加强与其他物种的联系,则有助于植物群落向稳定共存的顶级群落演替。种间联结理论认为负关联往往表明群落间的物种存在排斥性,这主要是由于不同物种的生物学特征与水分、光照等环境因子的需求存在较大差异,或者是有些物种为了占据相同的生态位而产生了激烈竞争所导致(徐满厚等,2016)。正关联则是反映了群落物种面对相同环境具有相似的生活策略,植物种对间互利共生,资源互补,相伴生长(陈玉凯等,2011)。本研究χ2检验结果表明海南粗榧群落主要树种105个种对均无显著关联种对,而正关联33 对,仅占总对数的31.43%,表明群落中相互依赖、联系紧密的种对较少,说明海南粗榧群落结构较为松散,稳定性弱,各树种间独立性强,这与濒危植物黄梅秤锤树(Sinojackiahuangmeiensis)的种间联结性研究结果相似(王世彤等,2018)。联结系数AC值与共同出现百分率PC值均为系数较小种对占群落总对数绝大多数的结果也验证了χ2检验结果,两种关联强度指数值均较大的种对有短药蒲桃和沙煲暗罗、白背厚壳桂和沙煲暗罗、赛木患和白背厚壳桂等种对,这可能是由于这些物种在生境需求和资源获取方面具有较高的一致性。短药蒲桃是与海南粗榧共同出现的次数最多的植物,两者均为耐阴湿植物,生长环境相似,且短药蒲桃为小乔木,与高大乔木海南粗榧占据森林中的不同的生态位,避免了种间的强烈竞争,是海南粗榧合适的伴生树种。Pearson相关性与Spearman 秩相关性检验结果发现海南粗榧仅和药用狗牙花具有显著负相关,这与海南粗榧喜欢生长在阴凉潮湿的沟谷环境,而药用狗牙花喜欢生长在阳光充足的温暖环境中有关,且由于海南粗榧幼苗早期生长对光照存在一定需求,与林下生长的药用狗牙花可能存在光照资源上的竞争。共同出现百分率结果表明海南粗榧与蓝树、沙煲暗罗、红果樫木等物种共同出现的频率较高却不存在显著相关,说明由于各物种可以通过占据不同生态位,利用不同空间的资源,从而在群落达到动态共存,同时也说明其他物种进入该群落的概率较大(薛卫星等,2023)。 林下的调查结果发现海南粗榧母树更新幼苗数量较少,且仅在1 个样地发现1 株幼树,可知其的天然更新繁衍能力较弱,戴志聪等(2010)连续10 年在海南岛对野生海南粗榧幼苗的存活率进行观测,发现复杂的雨林环境,尤其是林下光照资源不足对其生长产生了较大影响,侧面说明了植被对光照的竞争可能导致群落呈现总体负关联。根据前人的研究(王献溥等,1994)可知,海南粗榧为雌雄异株,母树的结实率低,并且种子有休眠特性,通常落地之后需要到翌年甚至第3 年的春天才能发芽,而大部分由于不能接触到土壤,往往在休眠过程中逐渐失去活性然后腐烂,此外其果实深受松鼠等啮齿动物的喜爱,成熟的果实常被动物吃掉,而在种子即将成熟发育的时期还容易遭到台风的侵扰,造成其结实率降低,调查中还发现海南粗榧样地的郁闭度均在0.6 以上,部分甚至超过了0.8,过高的森林郁闭度使得幼苗难以获得满足生长所需的阳光(叶兴状等,2021),这些自然因素是导致海南粗榧陷入濒危境地的重要原因,但最为关键的影响因素是人类对其的过度采伐利用,导致其难以形成自然更新的群落(祁珊珊等,2010)。 种间联结分析的本质是根据物种出现与否来分析种对间的关系,不考虑到物种的多度,频度等信息的影响,本研究中的海南粗榧数量并不影响联结分析结果,但对于以物种在样方中多度为基础,对物种间的关联强度进行分析的种间相关分析,海南粗榧在调查样方中的成株仅为8 株,数量特征并不明显,可能会导致海南粗榧与其他树种间的相关系数偏低。后续研究可通过增加样方数量,结合林下调查数据验证本文结果,同时通过分析相关种在环境中所占据的生态位、不同树种的发育周期及功能性状的差异及灌木层与草本层的物种生长状况对海南粗榧与其他物种的种间生态关系开展进一步探究。 综合以上研究结果,海南粗榧群落常见树种间的种间联系较为分散,总体呈现不显著的负关联,种对间的关联性较弱,显著相关的物种较少,群落处在动态共存的演替阶段中。海南粗榧在群落中的独立性较强,成株生长状况较好,但调查结果也发现林下海南粗榧幼苗、幼树数目极为短缺,说明林下植被对资源的竞争可能导致海南粗榧幼苗生长受限,加之受到自身繁衍困难的影响,海南粗榧种群存在逐渐衰退的风险,因此需要在保护海南粗榧个体的同时,结合群落的结构特征采取适当的人工辅助措施,对高大乔木进行修枝和开林窗,改善林下植被环境,保证海南粗榧幼苗获得充足空间资源,增加其幼苗的光照时长;收集海南粗榧种子培育成健康幼苗后,根据其生态习性,选择合适生境进行人工补植,扩大其分布面积,增加其种群数量,满足其对阴暗潮湿生境的需求,促进海南粗榧种群的生长与更新。2 结果与分析
2.1 伴生树种组成

2.2 伴生树种区系分析

2.3 总体关联性分析
2.4 种间关联性分析



3 讨论
4 结论
猜你喜欢
植物研究(2023年5期)2023-09-09
河南师范大学学报(自然科学版)(2022年5期)2022-08-08
数学物理学报(2022年2期)2022-04-26
新世纪智能(数学备考)(2021年9期)2021-11-24
新世纪智能(数学备考)(2020年9期)2021-01-04
中学生数理化·高一版(2018年10期)2018-11-08
现代园艺(2018年2期)2018-03-15
中国林业产业(2016年5期)2016-04-03
中国林业产业(2016年5期)2016-04-03
杂草学报(2015年2期)2016-01-04
