
气候变化情景下大花杓兰在中国的适生区预测
2024-01-25 06:49董树斌亓秀金杨明琪任志河刘青昊
生态学报 2024年1期
吴 齐,董树斌,杨 蕾,亓秀金,张 毓,杨明琪,任志河,刘青昊,程 瑾,*
1 北京林业大学生物科学与技术学院,林木育种与生态修复国家工程研究中心,花卉种质创新与分子育种北京市重点实验室,北京 100083 2 国家植物园,北京 100093 3 北京林大林业科技股份有限公司,北京 100083 4 河北省大海陀国家级自然保护区管理处,张家口 075500
气候变化能够影响植物物候期、形态特征、生理指标,或通过改变其生长环境引起分布范围的适应性迁移,甚至是物种消亡[1-2]。有研究表明,第四纪以来气候冷暖和干湿的频繁交替以及未来时期的气候变化,不仅会改变植物本身的物候生长特征,并且能通过影响周围区域内其他物种的生存状态从而对各植物类群的生长产生间接影响[3-4]。物种分布模型(Species Distribution Model,SDM)能够将已知的物种分布信息与其分布地的环境变量相关联,以此来模拟物种适宜栖息地的空间分布[5-6]。结合大量数字化标本记录和高分辨率环境变量数据,物种分布模型在气候变化背景下预测生物多样性丧失[7]、警告生物入侵风险[8]和设计规划濒危物种保护区域等领域都有广泛的应用[9-10]。近年来,多种基于算法的物种分布模型相继被开发出来,根据分布数据预测动植物物种的空间分布也随之被应用于保护研究[11-12]。最大熵模型(MaxEnt)被普遍认为具有操作便捷、运行速度快和预测精度高等优点,是分析影响珍稀濒危植物等植物的关键环境变量和分布情况等研究的首选模型[13-15],相关成果为研究物种的保护提供了指导性建议,并进一步引发人们对气候变化的持续关注。
兰科是被子植物中最大的科之一,是一个拥有众多珍稀濒危物种的草本类群。所有野生兰科物种均被列入《濒危野生动植物种国际贸易公约》(CITES)应保护植物物种[16]。2021年9月颁布的《国家重点保护野生植物名录》中,兰科植物约占整个名录保护物种数量的30%[17]。兰科植物在种子萌发、生殖等生活史关键环节中对传粉者、土壤微生物等存在特殊要求[18-19],其物种形成与分化受到生物因素和非生物因素的共同影响,因此,开展兰科植物全面系统研究对深刻理解生物多样性具有重要意义[20]。近年来,全球范围内气候变化和人类活动是许多受威胁兰科植物栖息地缩小的重要影响因素[21-24]。此外,气候变化还会导致一些原先分布在低海拔的物种向较高海拔地区迁移[25],这可能导致分布于高海拔地区的兰科植物由于较差的生境适应性和种间竞争而处于生存劣势[26-27]。因此,有关兰科植物应对气候变化的响应以及何种环境变量是影响其分布的关键性因素成为热点研究领域[28],相关的研究成果也为兰科植物保育策略的制定提供了理论参考和科学依据。
大花杓兰(Cypripediummacranthos)隶属兰科(Orchidaceae)杓兰属,是我国北方温带地生兰的典型代表物种,主要分布于高山草甸、林下、林间草甸和林缘等区域[29]。与大多数杓兰属植物分布在我国西南山区不同,大花杓兰在我国主要分布于华北、东北和台湾等地区[29-31],独特的分布格局预示其在研究兰科植物适应和进化方面具有重要的科学价值。大花杓兰花色艳丽、花形独特(图1),具有极高的观赏价值,但多年来气候变化加上盗采盗挖等人为干扰导致其栖息地破碎化程度日益加剧[32]。此外,大花杓兰自身繁殖过程中欺骗性传粉机制带来的低自然结实率,以及自然状态下种子发育和萌发中的一些限制因素,使得大花杓兰野生种群数量减少,大多数现存的野生种群也是一些植株数量较少的小种群,且呈零星状态分布[33],已被收录进《中国生物多样性红色名录-高等植物卷》(级别:濒危EN)、《濒危野生动植物种国际贸易公约》(CITES)和《国家重点保护野生植物名录》(级别:二级),迫切需要提高保护力度和制定针对性的保护策略,全方位推进大花杓兰种群的保护与复壮。但目前大花杓兰的大多数研究仅涉及授粉、种子萌发和根际真菌等方面[34],为全面了解大花杓兰种群分布现状,并为其资源的系统调查和保护提供思路和技术支持,本研究选用MaxEnt物种分布模型对大花杓兰分布数据建模分析,预测当前情景下大花杓兰在我国适生区分布情况,确定影响大花杓兰分布的关键环境变量,并预测过去和未来适生区分布及其面积的变化情况,分析其种群迁移路径。上述研究结果将指导大花杓兰资源调查范围的确定,优先保护生境区域的划定,为大花杓兰的就地保护、迁地保护和回归区域的规划提供一定的理论指导和科学依据,同时也为植物保护中的“旗舰类群”兰科植物保育工作的开展提供研究案例。

图1 盛花期的大花杓兰 Fig.1 Cypripedium macranthos at the full flowering stage
1 材料与方法
1.1 物种分布模型的选用
选用最大熵模型(MaxEnt)为预测物种分布的模型。为检测该模型对大花杓兰适生区分布预测的适用性,对过去3个情景、当前情景和未来4个情景环境数据分别进行MaxEnt模型运行,使用受试者操作特征曲线(receiver operating characteristic curve,ROC)下面积值(areas under curves,AUC)作为模型评估指标,AUC值范围0-1,当AUC值大于随机预测分布模拟值0.5时表示模型可信,越接近1表示预测结果越准确[35-37]。
1.2 物种分布点数据采集和处理
从中国数字植物标本馆(CVH,https://www.cvh.ac.cn/)以及全球生物多样性信息设施网络(GBIF,https://www.gbif.org/)中获取大花杓兰的地理分布数据。另外,对北京、河北和黑龙江等省市开展过多次野外调查,获取了大花杓兰的部分地理分布数据。分布数据收集完毕后,使用Google Earth (http://www.Google.com/Earth)为其中缺失准确地理坐标的分布点补齐经纬度数据,并进行分布点的筛选和修正(确保每10 km×10 km网格单元内只出现1个数据点),最后输出为MaxEnt软件建模所需的“*.csv”格式的数据用于后续分析。经过对采集到的167个大花杓兰地理分布点筛选和修正后,共有80个分布点数据用于后续模型分析(图2)。
1.3 环境变量的筛选
大花杓兰一般分布于林下、林缘、草坡等富含腐殖质的土壤和排水良好处,光照、温度、降水及土壤等环境因素对其生长有重要影响[38-39]。因此,本研究选取了紫外线辐射、气候、土壤等38个环境变量进行生态位建模分析。从全球气候数据网站(WorldClim-Global Climate Data,http://www.worldclim.org/)分别下载获得8个气候情景下的19个生物环境变量(BIO1-BIO19)数据,8个情景包括当前情景,末次间冰期(Last Inter Glacial,LIG,约12-14万年前)、末次盛冰期(Last Glacial Maximum,LGM,约2.2万年前)和全新世中期(Mid Holocene,MH,约6000年前)3个过去情景,以及IPCC第五次评估报告中定义的2种温室气体排放典型浓度路径(RCPs)下4个未来情景,分别是RCP2.6-2050、RCP2.6-2070、RCP8.5-2050和RCP8.5-2070,其中2050和2070分别代表2041-2060年和2061-2080年,模式为通用气候系统模式CCSM4,空间分辨率为2.5 arcmin。除此之外,还从全球紫外线辐射宏观生态数据网站(gIVU,https://www.ufz.de/gluv/index.php?en=32435)获得6个紫外线辐射环境变量(Ultraviolet-B radiation,UV-B1-UV-B6)数据。从威斯康辛大学网站(http://nelson.wisc.edu/sage/data-&-models/atlas/maps.php)收集了土壤pH值(Soil pH,SpH)、生长度日(Growing degree days,GDD)、土壤有机碳含量(Soil organic carbon,SC)环境变量。海拔(Global elevation,GloElev)、土壤质量(Soil quality,SQ1-SQ7)、森林覆盖率(Forest land,FOR)、草/灌木/林地(Grass/Scrub/Woodland,GRS)环境变量则是从联合国粮食及农业组织(Food and Agriculture Organization of the United Nations,http://www.fao.org/soils-portal/data-hub/soil-maps-and-databases/harmonized-world-soil-database-v12/en/)下载获得。
为消除模型的过拟合以提高模型预测的准确度,首先通过MaxEnt模型中的刀切法(Jackknife)检验上述当前情景下38个环境变量的贡献率[40],预跑20次后,剔除贡献率为0的变量。之后对剩余变量进行多重共线性分析,去除高相关性变量,计算变量之间的pearson相关系数(pearson correlation coefficients,r)[41],若两个变量间|r|≥0.8则认为相关性较强,去除贡献率低的变量,再运行20次,最终筛选出影响大花杓兰在我国适生区分布的关键环境变量。
在统一分辨率和坐标系后,使用ArcGIS 10.7的Extract工具从上述世界范围内的气候情景环境数据图层提取出中国范围的相应环境变量图层,用于后续MaxEnt模型的各项分析。
1.4 大花杓兰适生区预测
为了更直观地展示适生区面积和分布特征,采用自然间断点法(Jenks)将大花杓兰的适生区划分为4类:高适生区(0.6-1)、中适生区(0.4-0.6)、低适生区(0.2-0.4)和非适生区(0-0.2)[10,42-43]。通过ArcGIS对生成的环境变量图层二值化分析后进行差值计算8个情景下我国大花杓兰种群的潜在分布区域。
1.5 大花杓兰适生区面积变化与分布中心迁移
以当前情景适生区面积为参照,使用MaxEnt模型对过去和未来情景的适生区进行交叉检验,以此预测和计算过去和未来情景大花杓兰的适生区面积变化情况。随后,通过追踪图层中心的变化确定从末次间冰期至未来8个情景的适生区分布中心及迁移路线。
2 结果
2.1 模型精确度评估
当前情景下ROC曲线结果显示所得的AUC值为0.923(图3),远大于随机预测分布模拟值0.5,其他7个情景的AUC值也均在0.9以上,表明MaxEnt模型稳定可信,可用于大花杓兰在我国适生区分布的准确预测。
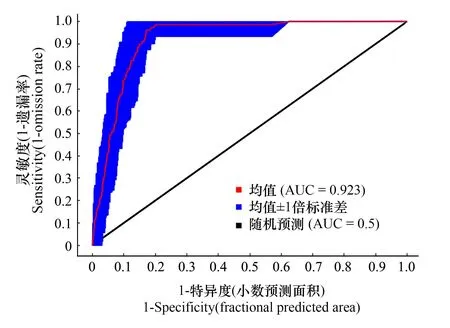
图3 模型验证AUC值Fig.3 AUC value of the model test 图中AUC表示受试者工作特征曲线下面积 Area under the receiver operating characteristic curve
2.2 当前情景下影响大花杓兰适生区分布的关键环境变量
刀切法共筛选出森林覆盖率(FOR)、UV-B最强月份均值(UV-B3)、最干季降水量(BIO17)、草/灌木/林地(GRS)、海拔(GloElev)、养分供应(SQ1)、降水量季节性变化(BIO15)、最热季降水量(BIO18)、土壤pH值(SpH)、最冷季平均气温(BIO11)、温度季节性变动系数(BIO4)等12个环境变量用于预测影响适生区分布的环境变量。其中,UV-B最强月份均值(UV-B3)、森林覆盖率(FOR)、降水量季节性变化(BIO15)、温度季节性变动系数(BIO4)和草/灌木/林地(GRS)环境变量对大花杓兰分布影响的贡献率位于前5位,分别是54.0%、14.3%、7.4%、6.8%、4.6%,累计贡献率达87.1%(表1),为关键环境变量。环境变量响应曲线显示UV-B最强月份均值大于约3482 J m-2d-1时高度适宜(存在概率>0.6)大花杓兰的生长,约3702 J m-2d-1时达峰值;大花杓兰分布概率与森林覆盖率、降水量季节性变化、温度季节性变动系数值正相关,森林覆盖率在46.33%以上高度适宜大花杓兰的生长;降水量季节性变化值大于约86.55%时高度适宜大花杓兰生长;温度季节性变动系数值在11611时高度适宜大花杓兰生长;草/灌木/林地在2.60%-34.27%时高度适宜大花杓兰生长(图4)。

表1 影响大花杓兰分布的5个关键环境变量Table 1 5 key environmental variables affecting C. macranthos distribution
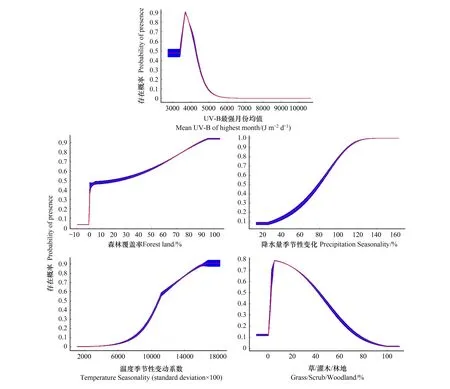
图4 5个关键环境变量的响应曲线 Fig.4 Response curves of 5 key environmental variablesUV-B3:UV-B最强月份均值Mean UV-B of highest month;FOR:森林覆盖率Forest land;BIO15:降水量季节性变化Precipitation Seasonality (Coefficient of Variation);BIO4:温度季节性变动系数Temperature Seasonality (standard deviation×100);GRS:草/灌木/林地Grass/Scrub/Woodland
2.3 8个情景下大花杓兰的适生区预测
适生区预测结果显示当前情景大花杓兰中高适生区主要集中在我国黑龙江省东部小兴安岭、吉林省东南部和辽宁省东部长白山脉、内蒙古东北部大兴安岭、河北省、北京市等地(图5)。末次间冰期大花杓兰中高适生区主要集中在我国东北地区;而末次盛冰期大花杓兰中高适生区主要集中在辽宁省、京津冀地区、山东省中北部等地,且是8个情景中适生区面积最少的时期;全新世中期大花杓兰中高适生区的分布范围与当前情景下非常相似(图6)。未来4种气候情景下,大花杓兰适生区与当前情景相比有增加趋势,RCP2.6路径下的2050年和2070年中高适生区与当前情景相差不大,均集中在我国东北地区小兴安岭、长白山、内蒙古北部大兴安岭、华北地区燕山、百花山、山东省南部等地,在东北地区的适生区进一步扩大;RCP8.5路径下的2050年和2070年中高适生区依旧分布在东北、华北等地(图6)。整体来看,过去情景较为寒冷的情况下,大花杓兰种群向西南方迁移,而在未来气候持续变暖的情况下,当前适生区的西部和南部有小幅度扩大。
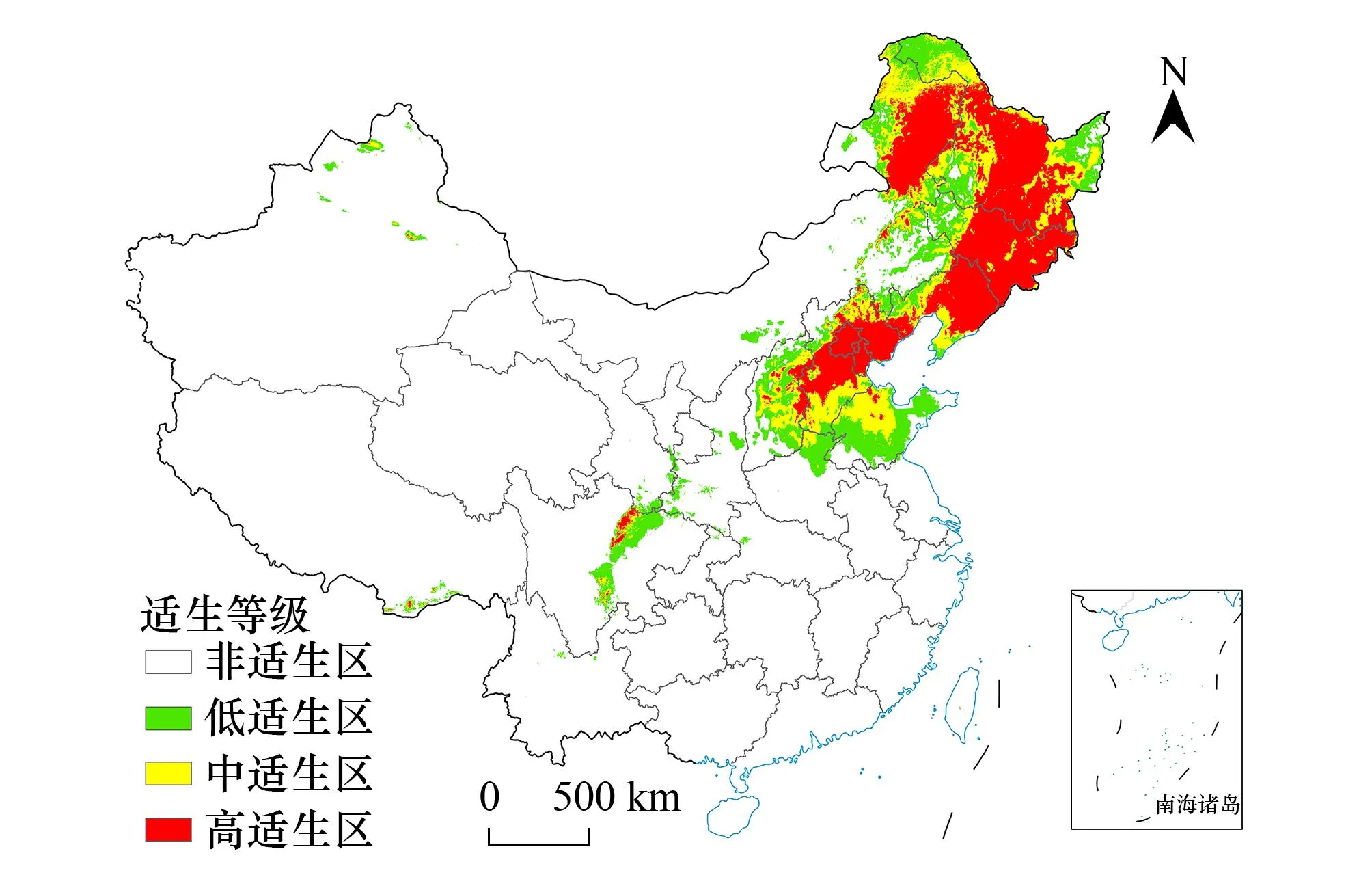
图5 当前情景下大花杓兰在中国的潜在适生区 Fig.5 The potential suitable area of C. macranthos at current scenario in China
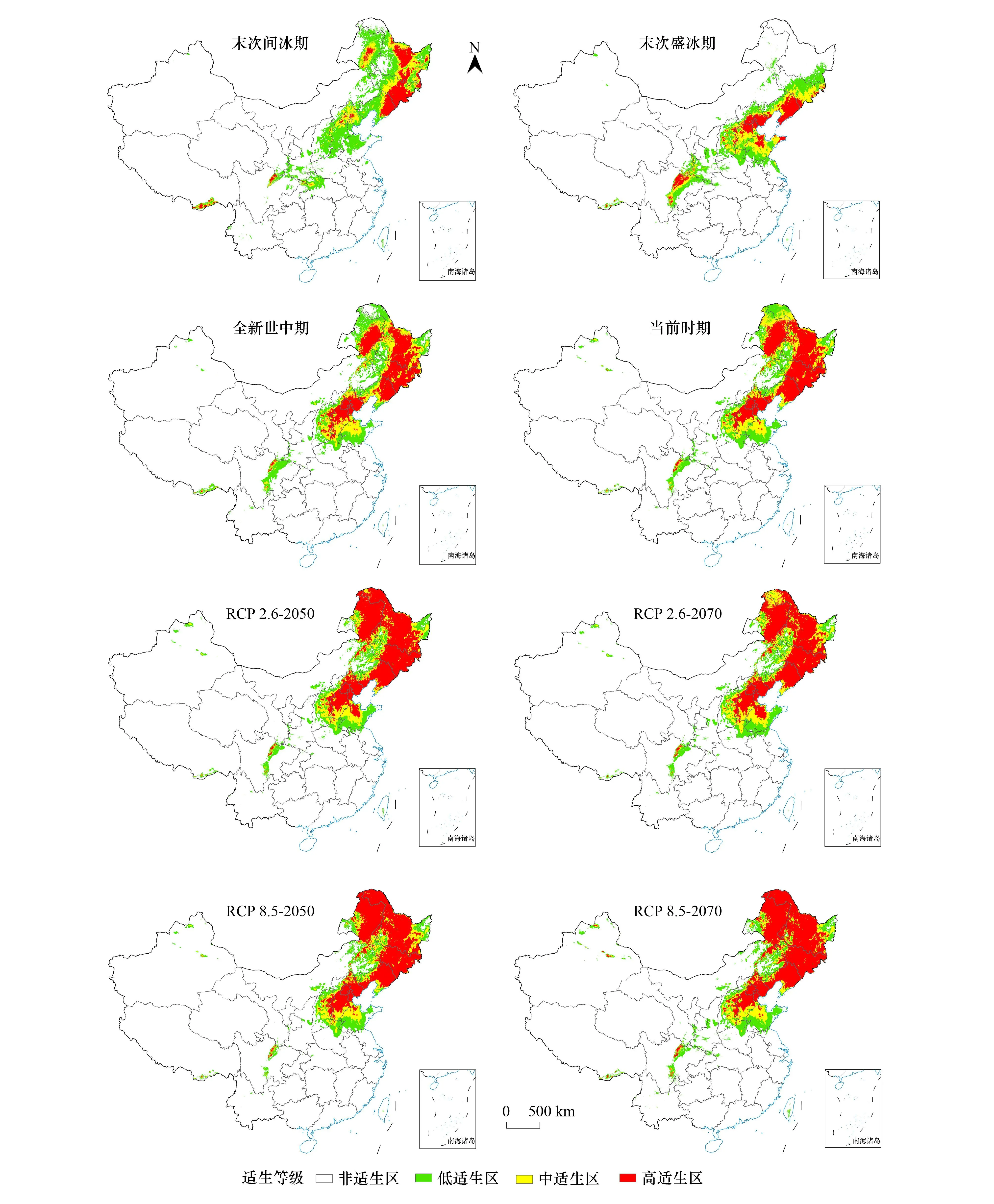
图6 8个气候情景下大花杓兰的潜在适生区 Fig.6 Potential suitable area for C. macranthos under 8 climate scenarios
2.4 气候变化下大花杓兰适生区的变化情况
上述适生区预测结果显示,整体上看,过去3个气候情景下,大花杓兰适生区面积呈现先减少后增加的状态(表2和图5)。与末次间冰期相比,当前情景下大花杓兰适生区面积有所增加,主要增加区域位于山东省南部、内蒙古东北部及黑龙江北部等地,而且面积增加主要集中在高适生区,增加的面积约438091 km2,位于我国中部地区以及西藏南部的适生区减少,面积减少127247 km2。与末次盛冰期相比,大花杓兰当前适生区面积增加的地区主要位于内蒙古东北部及黑龙江省等高适生区地区,在我国中部地区有部分减少,增加面积752941 km2,减少面积199422 km2。全新世中期与当前情景下大花杓兰适生区面积相似,增加的区域位于黑龙江北部,面积增加110155 km2,减少67047 km2。与当前情景相比,4个未来气候模式下大花杓兰适生区与当前情景相比有增加的趋势(表2和图7)。RCP2.6气候情景下2050年和2070年大花杓兰适生区增加的区域主要集中在山东省以南,中高适生区增加较多,多位于东北地区,2050年面积增加113003 km2,减少22155 km2;2070年面积增加108074 km2,减少26196 km2。在RCP8.5气候情景下,2050年和2070年大花杓兰适生区增加区域主要位于内蒙古北部与黑龙江北部,低中适生区增加较小,高适生区增加较多,2050年面积增加119627 km2,减少64586 km2;2070年面积增加165987 km2,减少15777 km2。

表2 不同时期大花杓兰各适生区的面积变化/km2Table 2 Suitable area for C. macranthos under different climate change scenarios

图7 8个气候情景下大花杓兰的适生区变化情况 Fig.7 Change of suitable area of C. macranthos under 8 climate scenarios
2.5 气候变化下大花杓兰适生区分布中心及迁移变化趋势
末次间冰期时大花杓兰适生区分布中心(42°37′4.8″N,121°26′49.2″E)位于内蒙古东南部,与辽宁省相近。末次盛冰期时,适生区分布中心迁移幅度较大,向西南方向迁移至河北省(38°27′14.4″N,117°22′44.4″E)。而到了全新世中期,适生区分布中心则又向北迁移回内蒙古东南部(43°5′27.6″N,121°3′50.4″E)。当前情景下大花杓兰适生区分布中心位于内蒙古东南部(43°43′19.2″N,121°28′1.0″E)。未来情景下,大花杓兰适生区分布中心依旧位于内蒙古自治区,RCP2.6-2050时大花杓兰适生区分布中心略向西迁移(43°41′49.1″N,121°18′54.0″E),RCP2.6-2070时适生区分布中心向南略有迁移(43°32′52.8″N,121°17′52.8″E);RCP8.5-2050时适生区分布中心向西北方向小幅度迁移(43°57′14.4″N,121°31′15.6″E),RCP8.5-2070时适生区分布中心向西南方稍有迁移(43°41′2.4″N,120°57′21.6″E)(图8)。
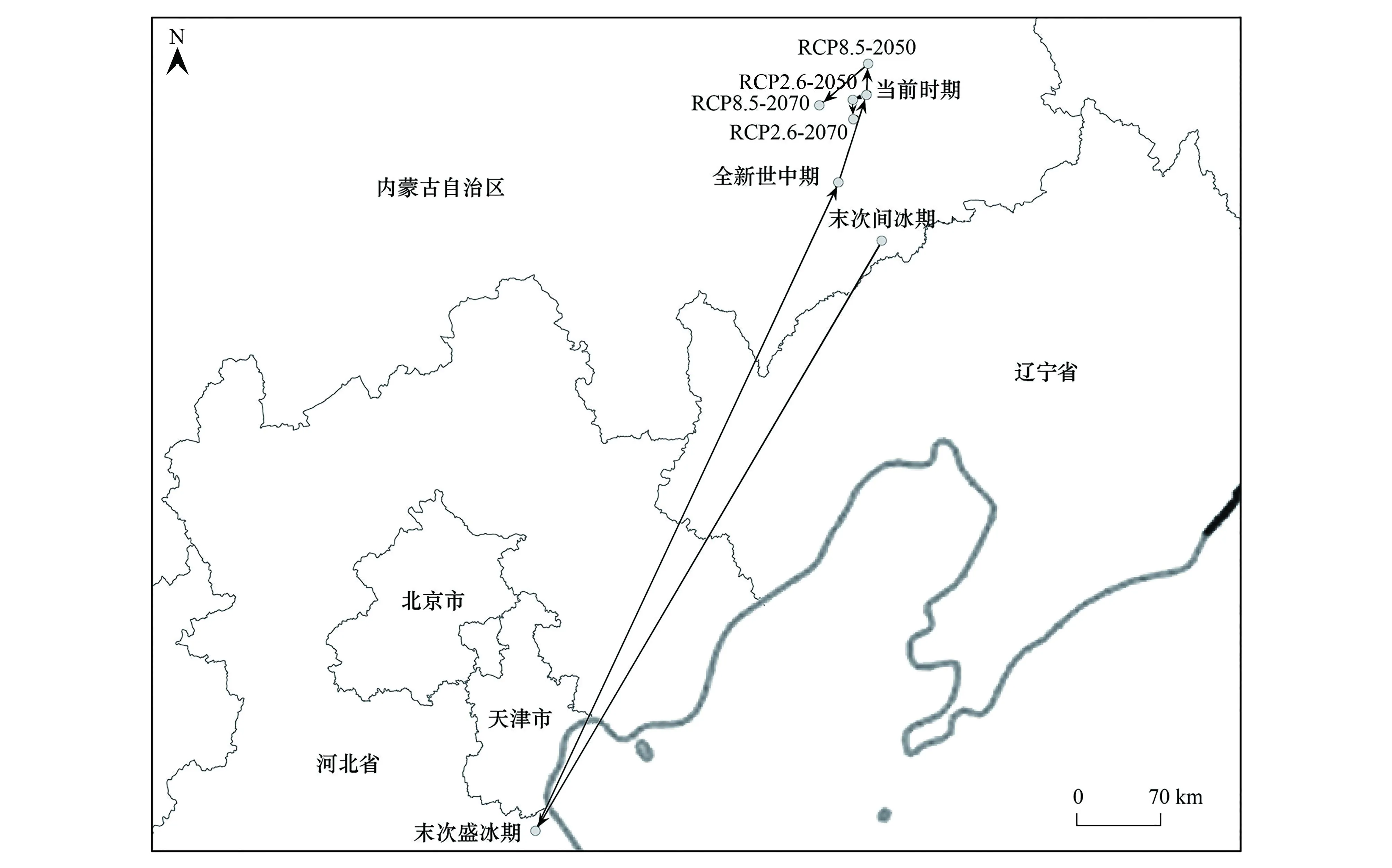
图8 8个情景下的大花杓兰分布中心迁移路径 Fig.8 Centroid migration of C. macranthos under 8 scenarios
3 讨论
3.1 影响大花杓兰适生区分布的关键环境变量
本研究结果显示UV-B最强月份均值是影响大花杓兰在我国分布的最关键环境变量,贡献率高达54.0%,其余四个关键环境变量是森林覆盖率、降水量季节性变化、温度季节性变动系数和草/灌木/林地,其中,UV-B最强月份均值、森林覆盖率、草/灌木/林地这三个环境变量的累计贡献率高达72.9%。大花杓兰适宜生长在偏中等森林覆盖度的区域[44],对百花山、玉渡山、海陀山、雾灵山等地大花杓兰种群实地调查时也发现,大花杓兰多分布于林下、林缘或草甸上,花期为6-7月,果期为8-9月[30]。北半球6-8月份为夏季紫外线峰值期,此时正是大花杓兰开花、传粉和受精以及种子初步发育的重要生长阶段,是其生活史中最旺盛的生长阶段。数据显示UV-B最强月份均值大于约3482 J m-2d-1时高度适宜(存在概率>0.6)大花杓兰的生长,约3702 J m-2d-1时达到峰值(图4),超过峰值后大花杓兰的存在概率开始降低。UV-B辐射是与光照密切相关的环境变量,自然环境中适量的UV-B辐射在一定程度上对植物的生长发育有利,如增加次生代谢产物、提高生物胁迫或非生物胁迫抗性[45-49]、提高光合特性以及促进成花[50-53],但过量的紫外线会对植物的生长发育造成不良影响,植物也将产生一定的生态适应性[54-55],如黄花杓兰在自然光照变化时,可以通过有效利用氮和较高的气孔和叶肉导度对抗光抑制来抵消其对强光的敏感性,从而保持叶片光合作用强度,提高光合性能[56]。但光照过强的环境中生长的兰花大多表现出“光合午休”现象[57-59],这一现象在杓兰属其他植物的研究中得到证实,如遮荫条件好的生境更适宜扇脉杓兰的生长,午休现象的时长也相对更短[60-61]。因此,我们推测适量的UV-B辐射对大花杓兰叶片形态结构有一定的影响,增强光合性能,合成更丰富的次生代谢产物,进而提升了对气候变化的适应能力。本研究森林覆盖率(贡献率:14.3%)和草/灌木/林地(贡献率:4.6%)两个关键环境变量与UV-B辐射变化密切相关,上述变量也会对大花杓兰生境的温度、湿度、土壤透气性等非生物因素和林下植物分布、传粉者分布、土壤中真菌分布等生物因素造成影响,进而影响到大花杓兰的营养生长和生殖生长[62-63]。
温度和水分等环境变量的综合效应,与山地植物分布规律有着密切的关系[64]。本研究显示,温度季节性变动系数越大,大花杓兰适生程度越高。大花杓兰是我国北方温带地生兰的典型代表物种,这些区域四季分明,夏季日照时间长,气温高,冬季日照时间短,气温低,分布于此的植物对于温度的承受范围较大。降水作为调节植物生长发育的关键变量对物种分布有重要作用。降水量较高的地区植被类型丰富,尤其是在生长季,降水充足是维持大部分草本植物生存的重要因素[65-66]。研究结果显示降水量季节性变化在86.55%以上时高度适合大花杓兰的生长,大花杓兰主要分布的华北和东北地区夏秋季节降雨多,春冬季降雨少,而夏秋季是大花杓兰生长和繁殖最为旺盛的阶段。此外,夏季降水的增加能够促进土壤微生物数量的增加[67],生长季内丰富的降水还可能在一定程度上通过促进地下真菌种类及数量的增加,进而促进大花杓兰的生长。对于杓兰亚科中杓兰属和兜兰属植物叶片形态结构和光合生理特性的研究结果表明,杓兰属植物有更高的光合诱导速度和光合速率[68],二者的叶片形态结构分别适应了自身独特的自然环境,杓兰属植物的叶片特征反映了对肥沃土壤、充沛土壤水分、明显温度和降水季节性波动环境的适应[69]。李中跃[70]研究表明,东北地区大花杓兰根茎叶的功能性状会随分布地纬度升高、年均温和年降水降低等变化而改变,如叶片增厚、气孔密度增加、比叶面积降低,根系直径增粗等,进而适应上述环境变化,这与本预测中温度和降水季节性变化对大花杓兰的分布具有高的贡献率这一结果是互相佐证的。
在基于MaxEnt模型预测气候变化情景下物种的适生区预测研究中,UV-B辐射、森林覆盖率和草/灌木/林地等被加入到环境变量中参与分析的研究案例并不多见,相关研究多以世界气候数据库中气候变量为主[15,71-73]。在有关杓兰属植物适生区预测的研究中,温度季节性变动系数、最热季度降水量、最干月份降水量、6月的降水量、2月的最高气温、最暖月降水、等温性、最热月最高温、最干季度降水量、降水季节性等相继被报道是影响其分布的关键环境变量[74-76]。在研究中,我们也运用仅19个气候变量进行了预测分析,结果显示贡献率前三位的分别为降水量季节性变化、温度季节性变动系数和最热季降水量,这与前人杓兰属的相关研究结果类似。这在一定程度上证明了本研究结果具有一定参考价值的同时,也意味着UV-B辐射对杓兰属植物分布的影响值得更多关注。已有的将紫外辐射相关环境变量纳入到物种分布预测的研究显示,这些变量对生长在温带气候范围内的植物分布贡献率较高,如影响灌木铁线莲、灰叶铁线莲、矮牡丹和木梨分布的关键环境变量中均至少有一个紫外辐射相关变量的贡献率在10%以上[77-79]。因此,后续对温带分布植物的适生区预测时,在环境变量筛选时建议增加与紫外辐射相关的一系列因素,使得研究更加全面,研究结果更为准确。
考虑到大花杓兰是典型的温带兰科植物,对气候变化背景下全世界分布的地生兰潜在适生区的主要研究进行了汇总,但由于地理范围大,相关物种的分布面积存在差异,影响亚热带、热带和温带兰科植物分布的环境变量间没有显著的共同点。当将目标物种范围缩小到我国地生兰科植物,关键环境变量与不同气候类型间也未见显著的相关性。这可能是缘于目前已开展的兰科植物分布的研究案例有限及研究结果相对复杂,但可以明确的是,对影响不同气候类型兰科植物关键环境变量的横向比较研究具有重要意义,将为兰科植物的适应性进化分析和保护工作的开展提供科学依据[28,71,74-76,80-81]。
3.2 我国大花杓兰分布的历史分布格局
大约在白垩纪晚期兰科植物开始出现,杓兰亚科是兰科中较原始的类群之一,为适应气候和地理环境的变化,逐渐形成了现今的物种多样性水平和广泛分布格局;杓兰属则是杓兰亚科中地理分布式样最为复杂的类群,与其他属植物的分布区域位于热带和亚热带不同,杓兰属主要分布于温带和亚热带地区[75,82-85]。刘焕楚[76]对杓兰属植物的生物地理学及生态位进化分析认为,随着中新世时期地球气候持续降温,杓兰属植物祖先对年均温的适应性进化可能是其分布格局变化的重要驱动因素。同关键环境变量的讨论中指出的,有专家学者认为杓兰属的分布受到了水分和温度因素的共同影响[75]。进入第四纪后,植物对冰期和间冰期交替引起的气候变化响应表现为地理分布迁移和适应性进化[86-87]。末次间冰期气温相对较暖,出现了较大范围的大花杓兰适生区。然而随着盛冰期的到来,气温逐渐下降,全球温度总体上约比现今低5-12℃,中国冰川面积是当前的8.4倍[87]。在遭遇恶劣气候条件时,大多数植物的分布范围都缩小到合适的栖息地而维持生存[88-90]。虽然大花杓兰具有较强的耐寒性,但气温的过度降低也使其遭受了极大的破坏,导致适生区急剧收缩,中高适生区主要保留在长白山、太行山脉等地。海拔2000 m以上的长白山地区存在中国罕见的高山冻原带,拥有较多古老孑遗植物[91],这些地区的独特地理条件可能使得其成为大花杓兰在冰期气候极端条件下的生境“避难所”。进入全新世时期,气温回升[92],中期温度与现今温度较为相似[93],总体温暖湿润的气候环境使得大花杓兰的适生区又有一定程度的扩张,这一时期大花杓兰的分布范围与当前时期的适生区相差不大。
随着全球变暖的趋势加强,未来气候的波动可能导致北半球许多物种的分布中心向北(纬度增加)或更高的海拔转移[94-96]。在RCP2.6和RCP8.5两种未来浓度路径下,大花杓兰的适生区均有不同程度的扩张,两种浓度路径下大花杓兰往高海拔地区迁移较多,但低浓度代表性路径(RCP2.6)大花杓兰的分布范围表现为向当前适生区的周边扩张,高浓度代表性路径(RCP8.5)下分布范围则在我国北部和南部地区的潜在适宜面积均有增加,且增加的面积较多。虽然有一些学者研究发现未来全球气候气温上升会使得一些种类的兰花的野外种群趋向灭绝[97],但也有研究认为温度上升对中高海拔适应性强的植物影响较小,同时,大花杓兰适生区进一步扩大也可能是对未来情景下降水量增多气候[98-99]的响应变化。同时,气候变化是一个渐变过程,对植物生存影响也是一个累积过程,短期内并不一定产生剧烈的影响[97],大花杓兰对全球变暖的应对趋势还需要长期的监测和进一步研究。
3.3 影响大花杓兰分布的其他因素
兰科植物常具有独特的传粉机制和高度特化的传粉系统[100],在统计的456种兰花里约67%仅有一种传粉者[18],因此传粉者的未来分布对兰科植物的适生区域有着重要影响。生态位模型预测Pseudorchisalbida、Leporellafimbriata、大黄花虾脊兰三种兰科植物未来传粉者的栖息地范围将明显收缩,三种兰科植物与其传粉者的重叠分布面积也将减小,在气候变化和传粉者分布减少的情况下,上述植物适生区范围逐渐减少的情况极有可能发生[71-73]。我国分布的大花杓兰传粉昆虫目前尚不十分明确,有学者报道了日本境内Cypripediummacranthosvar.rebunense具有欺骗性传粉系统,Bombuspseudobaicalensis是其唯一传粉者,其繁殖成功与否受到传粉者的影响[101-102],较低的自然结实率也在我国大花杓兰的研究中被报道[33]。此外,大花杓兰种子萌发以及植株生长依赖于特定的共生真菌[103],因此,传粉者和土壤微生物等生物因素也将对未来大花杓兰的适生区分布造成影响。刘焕楚[76]调查发现,在长白山中部许多地区有中等强度人类活动,大花杓兰的栖息地和生长受到了一定程度的负面影响,当将人类活动强度作为关键变量加入到大花杓兰未来分布的预测研究中,上述适生区将会消失。李中跃[70]通过固定样地监测及利用Leslie矩阵和密度限制增长模型预测了东北地区大花杓兰种群植株数量呈几何增长,并在短期内达到环境最大容量,引入密度限制因子后,经历较长时间种群达到饱和状态,认为大花杓兰自身的生物学特性并不会导致其濒危,人为干扰特别是非法采集则是主要致濒因素,破坏了大花杓兰及其共生生物赖以生存的环境,限制了大花杓兰种群的进一步扩大。
3.4 保护建议
研究发现预测的当前情景下大花杓兰适生区比目前标本采集区广泛得多,且在未来情景下大花杓兰的适生区略有扩增趋势。这一方面归因于来自于数据库的大花杓兰分布点大多收集于较早年代,近年来的可用分布点数据占比较少;另一方面,大花杓兰自然条件下野生种群的高适生区分布在长白山、太行山、大兴安岭、小兴安岭等地,这些地区气候条件和植被类型复杂多样,存在有许多尚未发现的新分布点的可能。2022年6月,内蒙古兴安盟索伦牧场和阿尔山市飞仙岭两地被报道发现多株野生大花杓兰(https://baijiahao.baidu.com/s?id=1737064482749903814&wfr=spider&for=pc),这两个新发现种群皆处于预测的当前大花杓兰种群高适生区内,这充分说明了通过物种分布模型预测来指导资源调查的思路可行。基于中国大花杓兰的适生区预测结果,针对其种群保护工作,提出以下建议:(1)在物种分布模型预测的中高适生区,开展更为详尽的资源调查,旨在系统全面地了解大花杓兰的分布和生长情况。(2)尽管根据预测结果华北是大花杓兰的高适生区,但是华北是目前我国大陆大花杓兰分布区域的南部边缘,容易受到相对更严重的人为干扰,值得关注和保护。(3)开展大花杓兰种群的长期监测,记录气候条件和环境变量,并进行种子萌发及生长、传粉和繁殖等生活史的生长监测,明确和保护其传粉昆虫及共生真菌,为构建大花杓兰种群健康评价和存活风险预警技术体系提供基础数据。(4)加强科普宣教工作,提高民众对大花杓兰等国家重点保护野生植物和珍稀濒危物种的保护意识,减少人为干扰。
猜你喜欢
中国野生植物资源(2023年9期)2023-10-20
中国中医药信息杂志(2023年6期)2023-06-13
电脑爱好者(2020年10期)2020-07-28
植物资源与环境学报(2020年2期)2020-04-20
电脑爱好者(2019年16期)2019-10-30
竹子学报(2018年2期)2018-10-25
青岛科技大学学报(社会科学版)(2015年4期)2016-01-25
河池学院学报(2015年2期)2015-02-27
河北林业科技(2011年5期)2011-07-10
商业经济研究(2009年26期)2009-10-27
