
籼型水稻两用核不育系遗传多样性与抗性品质基因分析
2024-03-03 12:18管俊娇张建华奎丽梅徐雨然谷安宇张锦文李小林
中国种业 2024年2期
管俊娇 张建华 奎丽梅 涂 建 邓 伟 徐雨然 谷安宇 张锦文 蓝 舵 李小林
(云南省农业科学院粮食作物研究所,昆明 650205)
水稻产量关系着粮食安全,其产量的提高离不开杂交水稻的成功培育[1]。目前水稻杂种优势的利用主要采用两系法和三系法,随着两系法的广泛使用,突破了传统水稻杂种优势利用的局面[2]。恢保关系制约了三系杂交稻的自由配组,限制了杂种优势的利用范围,而两系法不需要保持系,可自由配组,能广泛地利用杂种优势。因此,两系法育种发展迅速,越来越受到育种家们的重视,育成的品种已大面积推广种植,为保障我国粮食安全起到了关键作用。近年来,两系杂交籼稻成为了长江流域的主要种植类型[3]。随着市场需求和种植环境的变化,两系杂交稻的发展也面临着更新换代的挑战。
病虫害会对水稻产量和品质造成无法估量的损害,化学农药仍然是目前主要使用的预防手段,大量化学农药的使用除污染环境外还使病虫害的抗药性增强,形成了恶性循环。发掘抗病虫害基因,培育抗性品种成为了实现绿色生产的前提。褐飞虱是中国水稻主要虫害之一,研究人员在水稻上定位了46个褐飞虱抗性基因,7 个褐飞虱抗性基因已经被克隆[4]。水稻白叶枯病的暴发对水稻具有巨大的破坏性,现已有38 个抗白叶枯病基因(Xa)被定位,其中15 个抗性基因被克隆[5]。稻瘟病是水稻三大病害之一,已鉴定抗稻瘟病基因有100 多个,其中36 个被克隆[6-7]。
高产优质是育种家所追求的育种目标,杂交稻稻米品质受不育系品质的制约[8]。加工品质和外观品质是除食味品质和营养品质之外的主要评价指标[9],虽然已有一些加工品质的QTL 位点被定位,但已克隆的仅有影响整精米率的Chalk5基因[10]。对外观品质影响较大的是垩白,现已定位的QTL 也较多,Chalk5基因也是控制稻米垩白的主效QTL[10]。有研究表明,BADH2/fgr基因控制了稻米香味[11]。根据这些已知基因开发的功能标记已广泛应用在水稻育种实践中,大大提高了水稻的育种速度。
华智生物技术有限公司开发的水稻1K 液相芯片包括1048 个目标区间内的5400 个SNP 标记位点的基因型。1048 个目标区间在参考基因上均匀分布,平均1cM/区间,在83 个水稻重要的性状/基因(抗稻瘟病37 个,抗白叶枯病4 个、抗褐飞虱10个,稻米品质基因8 个、育性基因6 个、产量性状基因6 个、生理发育基因8 个和耐逆基因4 个)200kb范围内平均有10 个紧密连锁的标记。其中有供体材料验证的功能基因信息标记87 个,包括抗褐飞虱基因(Bph3、Bph14、Bph15和Bph27)、稻瘟病抗性基因(Pi1、Pi2、Pi9、Piz、Pigm、Pita和Pikh)、抗白叶枯基因(Xa7、Xa13、Xa21和Xa23)、垩白度基因(Chalk5)、香味基因(Badh2)、低镉基因(OsHMA3)等。该芯片已用于水稻育种材料的鉴定、分子标记辅助育种及基因聚合育种等研究领域,为优异种质资源的挖掘和高效育种提供了有利工具[12]。
云南省农业科学院粮食作物研究所通过与地方软米品种等具有优良农艺性状和抗逆特性的优质稻种资源间的多目标性状聚合,育成了一批性状优良的两用核不育系。本研究以15 个自主选育的水稻两用核不育系为试验材料,利用水稻1K mGPS芯片(华智生物技术有限公司)构建不育系分子指纹图谱,分析水稻抗逆、品质相关基因的关联位点,参照芯片供体材料的功能基因信息,分析15 份不育系材料抗性基因及优良品质基因分布,对不育系材料的可利用性进行评估,为长江上游稻作区籼稻新品种遗传改良提供科学依据。
1 材料与方法
1.1 试验材料本研究中以14 份籼稻两用核不育系和1 份粳稻两用核不育系YUN1025s(CK)作为试验材料,2022 年种植于云南省昭通市水富育种基地(表1)。所使用的两用核不育系材料均由云南省农业科学院粮食作物研究所选育,其亲本包括了不同类型的材料。不育期内花粉败育彻底,转育温度22~23℃,开花习性优良,有较好的配合力,其中4 个品种:云223s(鉴定编号:滇鉴稻(2022)06 号)、云225s(鉴定编号:滇鉴稻(2022)07 号)、YUN992s(鉴定报告)、YUN1025s(鉴定报告)已通过云南省农作物品种审定委员会组织的鉴定。

表1 试验材料信息
1.2 试验方法SNP 分型和染色体作图 每个材料剪取5 株苗期叶片混样,提取DNA,并对DNA样品进行纯度和完整度检测后,送华智水稻生物技术有限公司采用1K mGPS 芯片对参试材料进行基因型检测。SNP 位点变异数、杂合率、等位基因频率、多态性信息含量(PIC,Polymorphism information content)采用PowerMaker 3.25 软件计算。染色体作图采用MG2C version 2.1 软件进行。遗传多样性及聚类分析 根据SNP 分型结果,样本的遗传多样性利用PowerMarker 软件采用Nei′s 遗传距离构建UPGMA 聚类图。不育系抗逆、品质特性基因位点的分析 参考供体材料功能基因信息,通过Excel完成数据整理与位点信息比对,分析15 份不育系材料抗性基因及优良品质基因分布。
2 结果与分析
2.1 SNP 位点多态性分析基因分型后共获得5293 个高质量SNP 位点分型数据,其中3302 个具有多态性。这些多态性SNP 位点在染色体上的分布如图1,其中位于水稻第1 染色体上的SNP 位点最多,有391 个,第8 染色体上最少,有173 个,平均每1Mb 的区间内含有7.6 个多态性SNP 标记。这些多态性SNP 标记能够满足后续分析的需要。
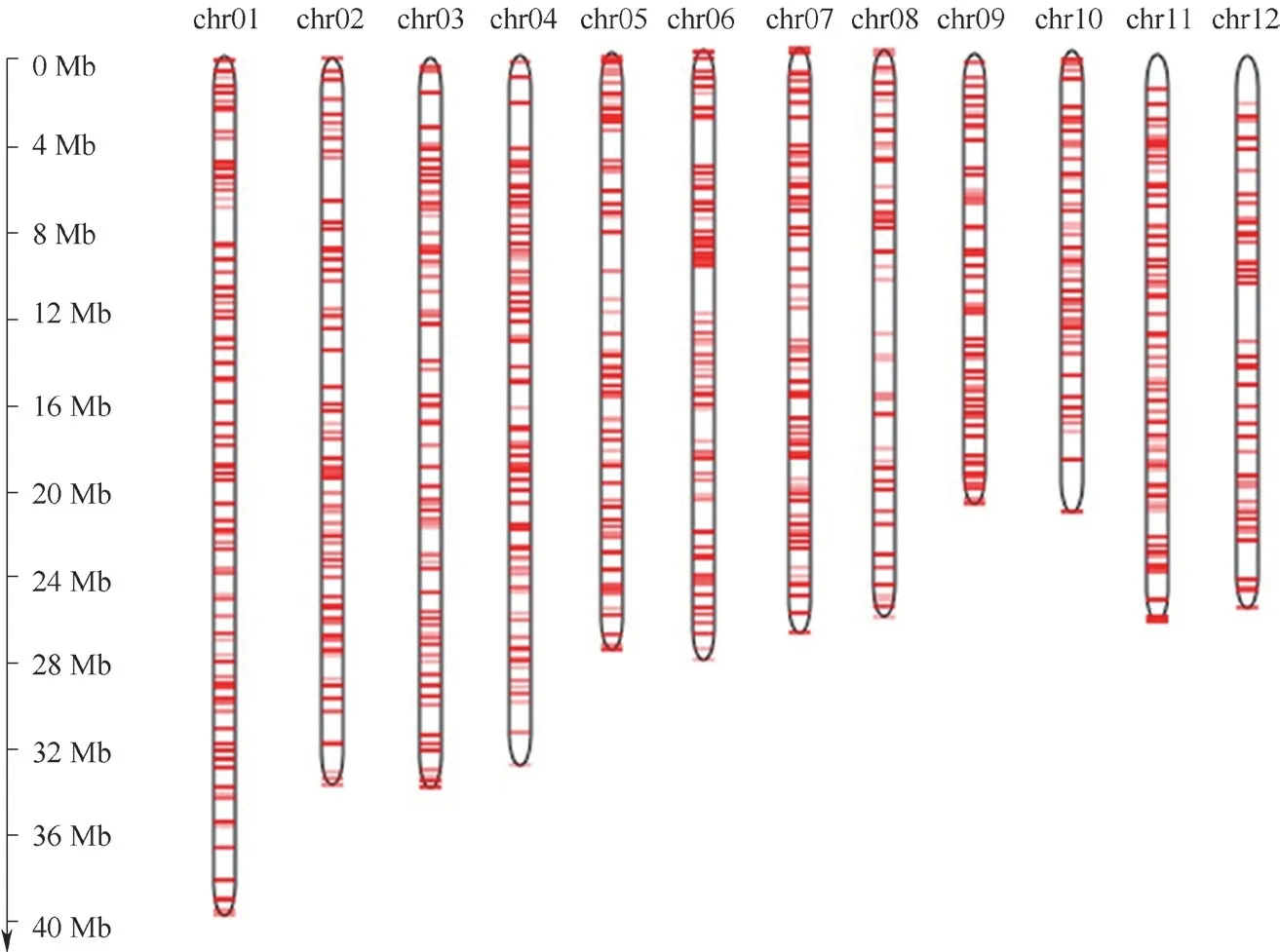
图1 多态性SNP 位点在水稻染色体上的分布
15 份不育系种质的遗传多样性分析结果表明,3302 个SNP 标记共检测到6640 个等位基因,主要等位基因频率(MAF,Major allele frequency)平均值为0.850,变化范围为0.064~0.970。期望杂合率(Heterozygosity)平均值为0.004。基因多样性在0.064~0.500 之间,平均值为0.204。多态性信息含量(PIC)变幅为0.062~0.375,平均值为0.264,最高值为0.375,其中1584 个SNP 标记的PIC 值在0.300 以上(图2)。多态性分析结果表明这些不育系材料的多样性较低。
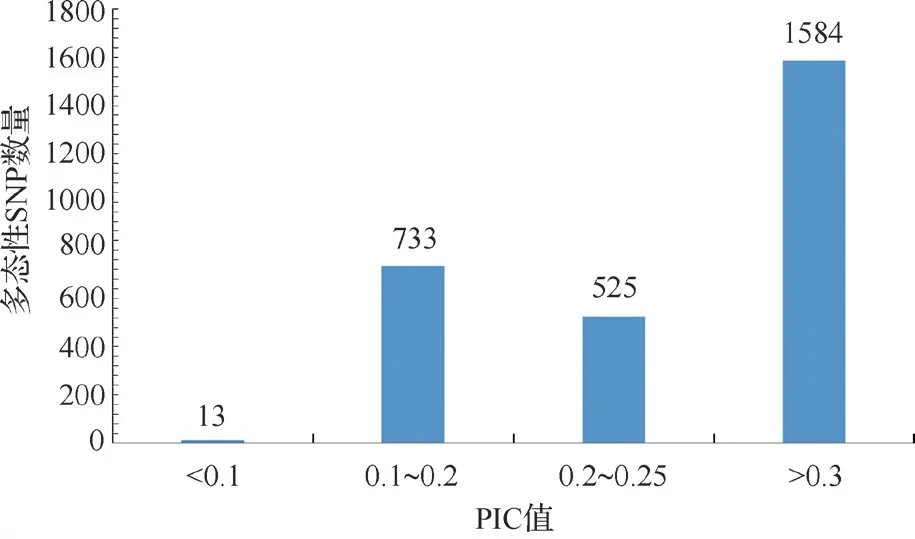
图2 SNP 标记在15 份不育系中多态性信息含量(PIC)的分布
2.2 不育系聚类分析根据Nei′s 遗传距离,对15个不育系进行聚类分析,结果表明,参试材料被划分为两大类群,即粳型不育系亚群(Ⅰ)和籼型不育系亚群(Ⅱ)(图3)。YUN1025s 是作为对照的粳型不育系材料,单独分作一个亚群。籼型不育系亚群中,S9 单独为一组,其他13 个不育系为一组,其中S7 和S8 具有相同的母本,遗传距离最近,仅有51个位点不同。S2 和S10 具有相同的父本,遗传距离为0.019,S6、S11、S1、云223s 4 份材料有一个亲本相同,也聚在了一起。以上聚类结果与亲缘关系一致,该组标记可用于不育系鉴定及亲缘关系溯源。
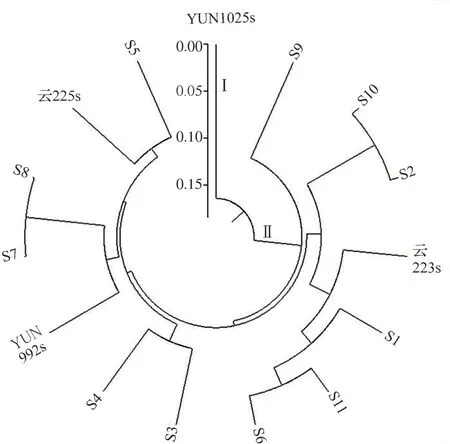
图3 基于Nei′s 遗传距离的聚类树状图
2.3 杂合率统计分析从15 份不育系的SNP 杂合率统计结果(表2)可知,各不育系材料的杂合率均较低,变幅为0.36%~2.54%。S2 的SNP 杂合率最高,为2.54%,YUN992s 的SNP 杂合率最低,为0.36%,说明这些不育系的基因组纯合度较高,具有较高的遗传稳定性。
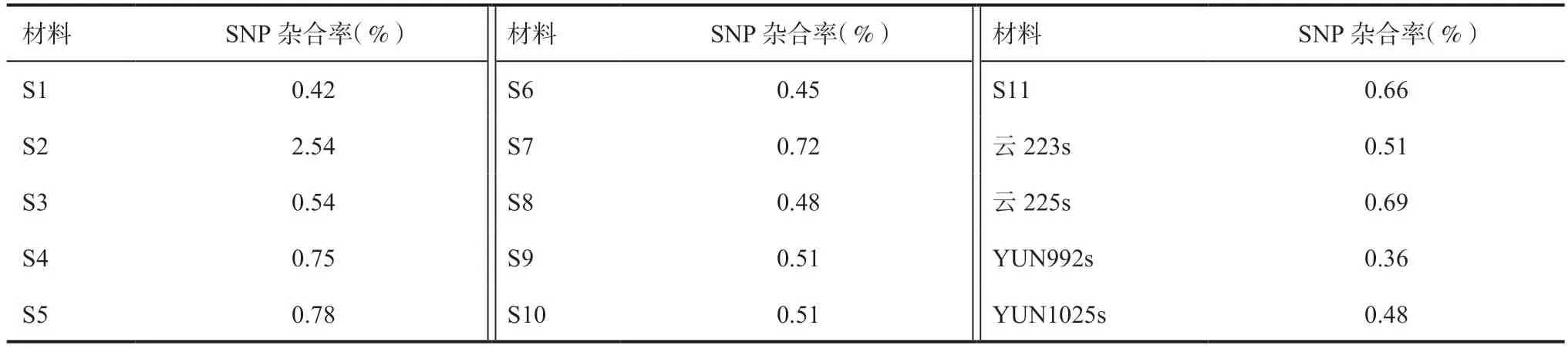
表2 水稻两用核不育系材料的杂合率分析
2.4 指纹图谱构建进一步对15 个水稻两用核不育系的有效位点进行比对筛选,筛选PIC 值大于0.37 且无缺失的位点构建15 个两用核不育系的指纹图谱。最终筛选获得384 个核心多态性SNP 位点,分布于12 条染色体上,通过Excel 及AI 绘图获得15 个不育系的特征分子指纹图谱(图4)。在本研究中所用材料在系谱上具有一定的亲缘关系,筛选出的384 个位点仍能完全区分这15 个材料,每个材料都有其独特的指纹图谱,说明这些位点多样性较丰富,具有较高的鉴别力,可考虑推荐这384 对引物作为不育系鉴定的核心引物。

图4 15 份不育系材料的DNA 指纹图谱
2.5 水稻核不育系抗逆及品质特性相关基因差异位点解析以某个基因区域全部检测位点与供体材料功能基因单倍型相似性0.9~1.0 为标准,通过比对参试材料与供体材料某个功能基因信息标记的相似度确定目标基因材料,若达到相似性标准,则参试品种判断为具有该目标基因,其表型与供体材料相似。由表3 可看出,15 个水稻两用核不育系中共有9 个基因,包含52 个位点与抗逆、品质优良特性相关,这15 个不育系大部分在抗褐飞虱、抗稻瘟病、稻米品质上有优异等位基因。S4、S9 有抗白叶枯病基因Xa21的优异等位基因,对水稻白叶枯病具有一定的抗性。S9、YUN1025s 有低镉OsHMA3基因的优异等位基因,可降低稻米镉毒性。S7、S9 检测到了香味基因Badh2。
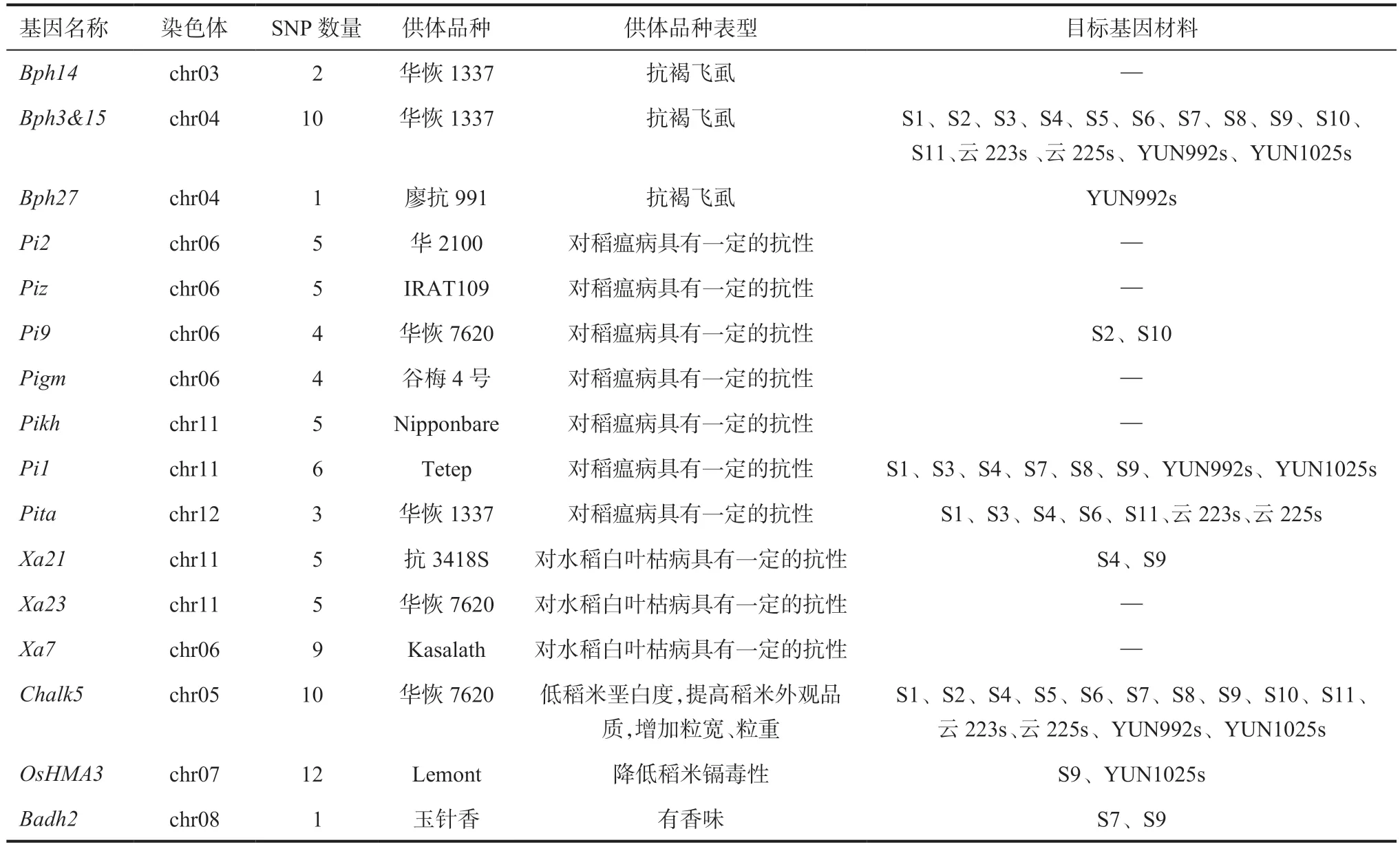
表3 水稻两用核不育系主要抗逆、品质相关基因SNP 位点统计
3 结论与讨论
遗传多样性研究对加快水稻的遗传改良及育种具有重要的指导意义。在育种中,充分利用遗传多样性研究,设置合理的育种目标,指导亲本选择,利用分子标记辅助选择技术(MAS)筛选目标性状,减少有害基因的影响,可以加快育种速度[13-15]。SNP 标记具有数量多、多态性丰富、适于高通量检测等优点,已广泛应用于分子标记定位、突变体基因的挖掘、构建指纹图谱、遗传多样性分析等研究[16-17]。余卫霖等[18]利用水稻56K SNP 芯片,构建水稻雌性核不育系SNP 分子指纹数据库,为雌性核不育系的分子鉴定及其优良抗逆特性的研究奠定了基础。本研究利用1K mGPS 水稻SNP 芯片对15 个水稻两用核不育系种质进行基因分型,分析了15 份水稻材料的遗传多样性。多态性位点PIC 平均值为0.264,介于0.25~0.50 之间,按照孙志广等[13]、Botstein 等[19]的理论,参试材料为中度多态性,但是其最高值仅为0.375,表明这些不育系材料的多样性较低,遗传基础较为狭窄,同质化严重。这是由于在不育系选育过程中,严重依赖于几个甚至一个骨干不育系进行选育,如农垦58s、矮培64s。此外遗传多样性分析能够准确分析材料间亲缘关系,通过聚类分析发现粳型不育系YUN1025s 单独归为一类,与其他的籼型不育系明显区别,具有共同亲本的材料首先聚到了一起,说明分子标记可以在分子层面验证不育系的系谱关系,可用于不育系的亲本追溯及指导新的不育系选育。
SNP 标记构建DNA 指纹图谱,已广泛应用于品种真实性和纯度鉴定。李梓榕等[17]通过精简水稻全基因组SNP 标记并构建指纹图谱,最少用12 个SNP标记就可以完全鉴定117 个水稻种质;余卫霖等[18]采用12475 个多态性SNP 位点构建的分子指纹图谱能够清晰地将10 个水稻雌性核不育系区分开,同时也为SNP 标记的精简留下了很大空间;田红丽等[20]利用筛选得到的玉米384 个位点的SNP 组合构建出的玉米杂交种指纹图谱,选取其中200 个位点能够区分目前95%及以上玉米已知品种。本研究中,通过筛选,采用分布于水稻12 条染色体上的384 个核心多态性SNP 位点构建了15 个不育系的指纹图谱,可为这些材料的真实性鉴定提供依据。
随着人民生活水平的不断提高,优质和绿色健康成为稻米生产的新要求,采用高产绿色优质新品种是实际生产中的首选。优良的不育系是选育优良水稻新品种的基础,MAS 技术、基因组编辑技术等现代生物育种技术则是未来育种的首要选择。褐飞虱在全国稻区均有发生,为害较重,Bph3是一个对褐飞虱具有广谱、持久抗性的主效基因[21]。本研究中,15 个不育系主要携带的抗褐飞虱基因主要是Bph3&15基因,有1 个材料同时携带了Bph3&15和Bph27基因。因此,这15 个不育系对褐飞虱有一定的抗性。稻瘟病是水稻的三大病害之一,15 个不育系携带的抗稻瘟病基因主要是Pi1、Pita基因,对稻瘟病具有一定的抗性[22-23]。白叶枯病是对水稻最具破坏性的病害之一,Xa21是白叶枯病显性抗性基因,对大多数白叶枯病病菌表现高抗[5,24],在本研究中,仅有2 份材料携带该基因。其他的抗性基因由于无对应的供体材料进行验证,并未对其进行分析,这15 个不育系材料在抗病虫害方面稍显不足,此后的品种改良中还需聚合更多的抗病虫害基因,增加不育系本身的抗性。在品质方面,大部分材料都携带低垩白度,增加粒宽、粒重的Chalk5基因,具良好的外观品质,有2 个材料携带OsHMA3基因,能有效降低稻米镉毒性,2 个材料携带香味基因Badh2。对这些不育系的遗传特征和育种潜力进行评价,有利于挖掘品质和抗性良好的优势组合,选育优质绿色品种,满足市场需求。
本研究利用SNP 标记对15 份水稻两用核不育系材料基因型检测,并进行遗传多样性分析。结果显示各不育系材料的杂合率均较低,为0.36%~2.54%,多态性信息含量(PIC)平均值为0.264,最高值仅为0.375,表明这些不育系材料的纯合度较高,多样性较低,遗传基础较为狭窄,同质化严重。通过分析水稻褐飞虱、稻瘟病和白叶枯病抗性位点在15 份材料中的分布,发现这批不育系材料携带的抗性基因较单一,在后续的品种改良中还需聚合更多的抗病虫害基因,增加不育系本身的抗性。对品质优异基因的分析发现,这些不育系具有较好的外观品质,部分品种具有低镉和香味基因,为优异不育系挖掘和利用提供了参考。
猜你喜欢
中国水稻科学(2023年3期)2023-05-18
世界科学技术-中医药现代化(2022年3期)2022-08-22
上海农业科技(2022年4期)2022-08-15
昆虫学报(2021年8期)2021-09-06
空间科学学报(2021年1期)2021-05-22
昆虫学报(2020年8期)2020-10-15
环境保护与循环经济(2017年5期)2018-01-22
西南农业学报(2016年6期)2016-04-16
中国果菜(2016年9期)2016-03-01
法医学杂志(2015年4期)2016-01-06
