
川黔地区濒危植物红豆树种群结构与数量动态特征
2024-03-10 19:10王明彬韦小丽韦忆王嫚余大龙袁刚毅
广西植物 2024年1期
王明彬 韦小丽 韦忆 王嫚 余大龙 袁刚毅

摘 要: 红豆树(Ormosia hosiei)是我国南方特有珍稀濒危植物,具有极高的经济价值和人文价值。为了解红豆树种群在川黔山区的生存现状及制约种群发展的原因,该文以川黔4个区域红豆树自然种群为调查对象,采用典型样地调查的方法,根据不同种群的分布特征,每個种群分别设置2个面积为20 m × 20 m的具有代表性的样地进行调查,通过编制种群静态生命表、绘制存活曲线、计算数量动态指数、利用生存函数及时间序列模型分析了川黔山区不同红豆树种群的结构与动态特征。结果表明:(1)河株村和干耳盘村种群龄级结构呈倒“J”型结构,陈家湾村和绒春村种群呈不规则“哑铃”型结构,红豆树种群自然更新能力强,但中龄级个体容易缺失。(2)4个种群均为增长型,存活曲线接近于Deevey- Ⅱ型,其中河株村、陈家湾村和绒春村种群消失率和死亡率呈“多峰型”,种群在生长中期容易受到外界干扰。(3)红豆树种群数量动态变化表现为幼龄级个体死亡率高,成熟龄级个体数量动态稳定的特点,其形成原因与山区地形特点和红豆树种子传播特性有关。(4)RDA排序结果显示,海拔和坡度是影响幼龄个体数量结构的主导因子,干扰强度是影响种群中高龄个体数量结构的主要因素。综上表明,川黔山区红豆树天然种群结构与数量特征的形成受制于山区地形特点、红豆树自身繁殖特性以及环境压力;通过对种群幼龄个体密度控制和中龄个体生长空间进行管理,可以提高种群幼苗存活率,促进中龄个体健壮生长,维持红豆树天然种群稳定性。
关键词: 红豆树, 濒危植物, 种群结构, 静态生命表, 生存函数
中图分类号: Q948.15
文献标识码: A
文章编号: 1000-3142(2024)01-0179-14
Population structure and number dynamic characteristics
of endangered plant Ormosia hosiei in
Sichuan and Guizhou provinces, China
WANG Mingbin, WEI Xiaoli*, WEI Yi, WANG Man, YU Dalong, YUAN Gangyi
( College of Forestry, Guizhou University, Guiyang 550025, China )
Abstract: Ormosia hosiei is a rare and endangered plant that is narrowly distributed in South China, with important economic and humanistic values. In order to find out its survival status and the reasons that restrict its development, we investigated the natural populations of O. hosiei in four regions of Guizhou and Sichuan provinces. According to the distributive characteristics of different populations, we set up two 20 m × 20 m for each population by using the method of plot investigation and conducted a community survey in each plot. The structural characteristics of O. hosiei populations were analyzed by establishing population static life tables and drawing populations survival curves. Four survival functions, number dynamic index, and time series model analysis were used to describe survival status. The results were as follows: (1)The age-class structure of O. hosiei populations were in the shape of “J”at Hezhu Village and Ganerpan Village, in the shape of “Dumbbell”at Chenjiawan Village and Rongchun Village, the populations had strong natural renewal ability, but middle-aged individuals were easily missing. (2)The four populations belonged to the increasing type, the survival curves of populations are close to Type Deevey-Ⅱ, the disappearance rate and mortality rate of O. hosiei populations showed a “Multimodal” pattern in Hezhu Village, Chenjiawan Village and Rongchun Village, indicating that the populations were susceptible to external disturbances in the middle period. (3)All O. hosiei populations in different regions appeared to decrease in the early stage and keep dynamically stable in the medium-late stage, and the formation reasons were related to the topographic characteristics of mountainous areas and the seed dispersal characteristics of O. hosiei. (4)RDA analysis showed that altitude and slope were the dominant factors affecting the populations structure of young individuals, and interference intensity was the main factor affecting the number structure of older individuals. All the above results indicate that the structural and number characteristics of O. hosiei populations in different regions are restricted by the topographic characteristics of mountainous areas, the reproductive characteristics of O. hosiei and the environmental pressure; By controlling the density of young individuals and managing the growth space of middle-aged individuals, we can increase the survival rate of the seedlings, promote the healthy growth of middle-aged individuals, and maintain the stability of the natural populations of O. hosiei.
Key words: Ormosia hosiei, endangered plant, population structure, static life table, survival function
珍稀瀕危植物指受自身原因或外界干扰的影响,其野生种群面临很大灭绝风险的一类特殊群体。环境条件的不断变化和人为活动的过度干扰,导致这类物种的生存受到严重威胁(马晨晨等,2017; 许恒和刘艳红,2018)。濒危植物保护生物学以濒危物种的生物生态学特性、生活史及生存环境为切入点,通过分析濒危植物的生存潜力、发展规律及其濒危机制,为濒危植物的保护和管理提供科学的指导依据(臧润国等,2016; 高岚等,2019)。其中,研究种群结构与动态是了解濒危植物生态特性的重要方法,也是探讨物种濒危原因的重要途径(王进等,2020)。通过静态生命表和存活曲线可以展现种群结构特征,了解种群不同年龄个体之间的动态差异,从中找出种群发展与变化规律(金慧等,2019; 唐凤等,2022)。通过生存函数及时间序列预测模型等统计学方法,可以分析种群生存现状及演替方向,评价种群与环境之间的相互关系,找到制约种群发展的关键因素(李娟等,2019)。种群结构与数量动态特征是种群生态学研究的核心内容,也是濒危植物保护生物学的重要方法,以种群结构与数量特征来探讨物种濒危机制,对濒危物种的保护和利用具有重要意义(邹天才等,2021)。
红豆树(Ormosia hosiei)为蝶形花科(Papilionoideae)红豆属(Ormosia)的常绿或落叶乔木,为国家二级重点保护野生植物。红豆树是我国特有种,主要分布于我国贵州、四川、福建、浙江等省份,是我国南方地区珍贵用材树种之一,也是南方地区重要的风水树种,具有极高的经济价值和人文价值(李峰卿等,2018a)。由于红豆树木材珍贵,树皮与种子被老百姓视为重要的药材(张琳婧等,2021),因而其遭到过度的利用与破坏,加之红豆树自然扩散能力差,其更新幼苗幼树均分布在母株周围导致激烈的种内和种间竞争,造成红豆树种群分布范围以及种群数量不断减少,趋于濒危状态(邱浩杰等,2020)。国内外有关红豆树的研究集中在生物学特性、遗传分布、育苗造林、人工林经营等方面(蔡勇,2007; Zhang et al., 2012; 尤根彪等,2017; 李峰卿,2018a)。针对红豆树天然种群特征的研究主要集中于浙江、福建等地,分布于溪流、河谷等流域附近的红豆树种群(谢慕华,2015; 刘鹏等,2017; 李杨等,2021),李峰卿等(2017)对3个小流域红豆树天然居群的研究表明,水流对红豆树种子的传播起重要作用,是影响红豆树种群发展的重要因素。对分布于西南山区,与河流相隔较远的红豆树种群生存现状、种群结构特征、种群发展规律及濒危原因的研究尚未见文献报道。缺乏种群生态学特征的相关研究,不利于该区域红豆树种群的保护和管理,因此,本研究以川黔地区生长于山地上的红豆树天然种群为研究对象,采用样地调查的方法,通过对分布于不同自然条件下的红豆树种群结构特征、发展规律及影响因素进行对比研究,拟探讨以下两个科学问题:(1)川黔山区红豆树天然种群生存现状如何,不同自然条件下种群结构特征与动态发展规律有何不同;(2)立地环境对红豆树种群结构特征有何影响,种群如何适应外界干扰。以期通过对川黔地区红豆树种群结构与动态的研究,为该区域红豆树天然种群的保护和管理提供科学的理论依据。
1 材料与方法
1.1 研究区概况
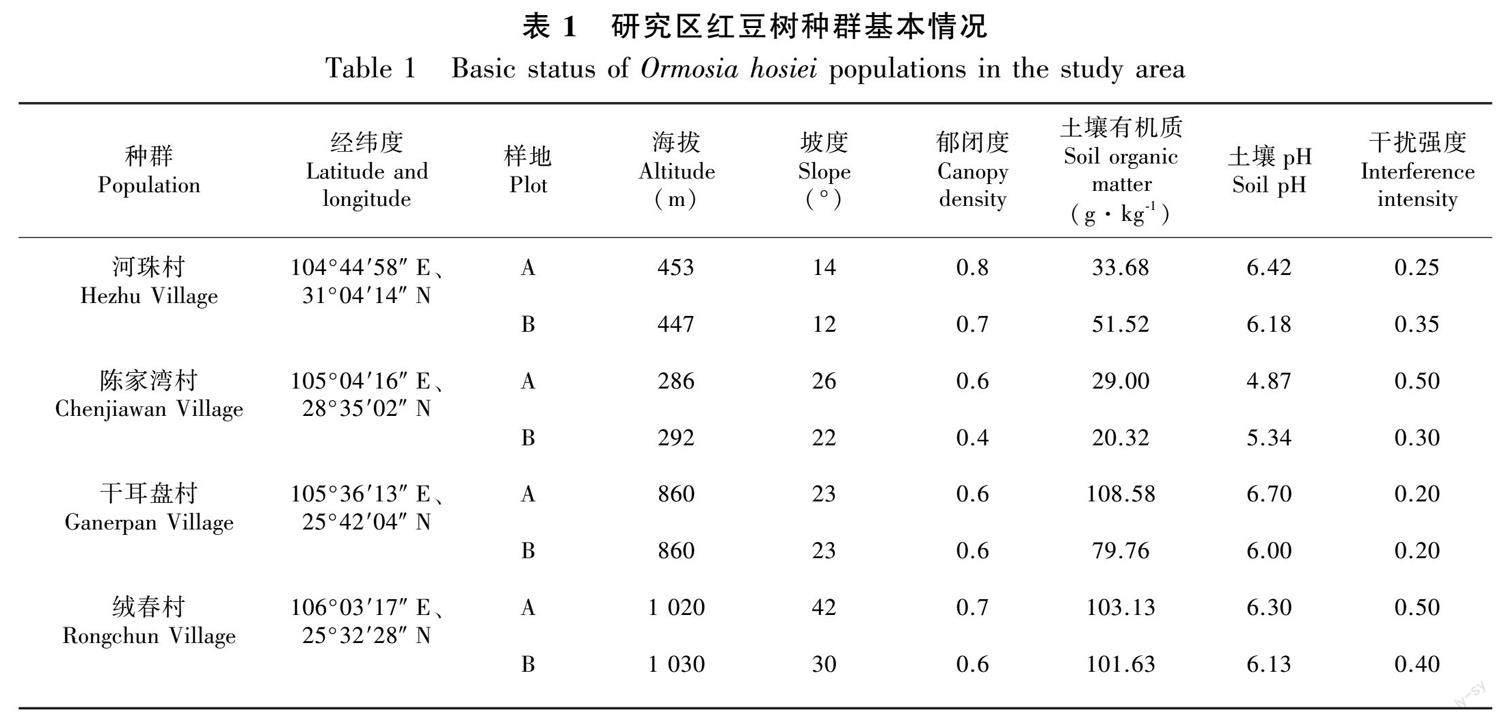
2021年6—11月期间,根据资料记载对贵州及四川两省14个样点进行实地踏查,发现存在4个种群数量较大的红豆树天然种群,即四川省中江县河株村和江安县陈家湾村、贵州省关岭县干耳盘村和望谟县绒春村。研究区地处四川及贵州两省,该区域属于亚热带季风气候,海拔在289~1 020 m,年平均气温16.7~19.5 ℃,年平均降雨量900~1 656 mm。4个红豆树分布区林分类型为常绿阔叶混交林或针阔混交林,除红豆树外,4个区域主要乔木树种有柏木(Cupressus funebris)、油茶(Camellia oleifera)、朴树(Celtis sinensis)、桑树(Morus alba)、枳椇(Hovenia acerba)、山核桃(Carya cathayensis)、枫香树(Liquidambar formosana)等。灌木层主要有油茶、楤木(Aralia chinensis)、杜荆(Vitex negundo)、吴茱萸(Tetradium ruticarpum )等。草本层植物主要有鳞盖蕨(Microlepia hancei)、鸢尾(Iris tectorum)、大叶仙茅(Curculigo capitulata)、香附子(Cyperus rotundus)、紫茎泽兰(Ageratina adenophora)等。研究区红豆树分布地理位置见图1,研究区基本情况见表1。
1.2 样地设置及调查方法
调查样地的大小取决于目标树种分布范围、地形以及种群数量(李伟等, 2014)。珍稀濒危植物受地理分布、地形条件及人类干扰的影响,种群分布区域可能相当有限,调查样地需要根据种群实际分布特征进行设计。Li 和 Zhang(2015)的研究表明,30 m × 30 m的样地可为濒危植物的种群结构研究提供很好的代表性样本。本研究根据红豆树不同种群的分布特征,每个种群分别设置2个面积为20 m × 20 m的具有代表性的样地进行调查。调查内容:(1)详细记录样地环境信息,即经纬度、海拔、坡度、郁闭度等数据;(2)对样地内所有红豆树及其他乔木树种进行每木检尺,分别记录物种名、胸径(diameter at breast height, DBH)、树高(tree height, H)等信息;(3)以5 m × 5 m为调查单元,对红豆树更新幼苗(H<50 cm)编号,记录其地径、高度等数据;(4)各样地内沿对角线各取3个0~20 cm土壤样品,用于测定土壤有机质、pH等;(5)不同样地的人为干扰强度根据调查结果参照袁刚毅等(2021)的方法进行赋值,最大赋值为1,最小为0。
1.3 年龄结构划分
采用得到广泛应用的“空间代替时间”,以树高或胸径代替龄级的方法进行红豆树种群特征分析(孟宪帅和韦小丽, 2011; 杨慧琴等,2020)。结合样地调查数据和红豆树种群的生长状况,参考张群芳等(2015)对红豆树种群龄级结构的划分方式,将树高(H)<50 cm的个体划为第Ⅰ级,H>50 cm且胸径(DBH)<5 cm的个体归为第Ⅱ级,之后DBH以5 cm为一个径阶,DBH≥25 cm则为第Ⅶ级。
1.4 种群动态量化
采用陈晓德(1998)种群结构动态量化分析方法对红豆树种群结构进行定量描述,以种群动态变化指数(Vn)来评价红豆树种群结构动态。
1.5 种群静态生命表和存活曲线
对原始调查数据进行匀滑修正(Bisht et al., 2013; 杨立荣等,2015),以修正后各龄级内个体数进行红豆树种群静态表编制。以标准化存活数为纵坐标,龄级为横坐标绘制红豆树种群存活曲线;参考Helt 和 Loucks(1976)的数学模型,采用指数模型Nx=N0e-bx来检验Deevey- Ⅱ型存活曲线,幂函数方程Nx=N0x-b检验Deevey- Ⅲ存活曲线类型。
1.6 生存分析
采用生存函数S(i)、累计死亡率函数F(i)、死亡密度函数f(ti)及危险率函数λ(ti)进行红豆树种群的生存分析(杨凤翔等,1991),了解种群动态发展规律。
1.7 时间预测模型
采用时间序列预测中的一次移动平均法(拓锋等,2021)对红豆树不同种群动态变化进行分析,预测经过2个、4个、6个龄级时间后不同红豆树种群各龄级个体数量。
1.8 数据统计分析
利用Excel 2010进行数据处理,Origin 2022统计分析及绘图。采用Canoco 5 软件的冗余度分析(RDA)方法检验红豆树种群数量结构与环境因子之间的关联性,以海拔、坡度、郁闭度、土壤有机质、土壤pH及干扰强度作为环境数据,参照国家林业行业标准中主要树种龄级与龄组划分(LY/T 2908—2017)方法及红豆树种群生长特性,将红豆树种群划分为3个龄组:幼龄组Ag1(包含第Ⅰ和第Ⅱ龄级)、中龄组Ag2(包含第Ⅲ、第Ⅳ、第Ⅴ齡级)、高龄组Ag3(包含第Ⅵ和第Ⅶ龄级),并将3个龄组个体数量作为物种数据,同时使用物种数据及环境数据对样地进行约束性排序分析(RDA)。
2 结果与分析
2.1 种群龄级结构特征及动态变化分析
使用胸径(DBH)及树高(H)分别绘制了4个红豆树种群的龄级结构。由图2可知,不同红豆树种群之间龄级结构存在明显差异。河珠村和干耳盘村总体表现为幼龄级个体数量较多,为中、老龄级个体数量逐级减少的倒“J”型结构,陈家湾村和绒春村表现为幼龄、老龄级个体数量多,为中龄级数量较少的不规则“哑铃型”结构。
从表2可以看出,河珠村种群在第Ⅰ和第Ⅱ龄级为衰退状态,第Ⅱ至第Ⅳ龄级为增长状态;陈家湾村和绒春村种群在第Ⅰ和第Ⅱ龄级为增长状态,第Ⅱ至第Ⅳ龄级为衰退状态;干耳盘村种群在整个龄级都处于增长状态。4个种群数量动态变化指数均为Vpi>V″pi>V′pi>0,表明4个红豆树种群均为增长型。4个种群在无干扰下增长性参数Vpi排序为陈家湾村>干耳盘村>河珠村>绒春村;对随机干扰的敏感性参数P′max排序为陈家湾村>河珠村=绒春村>干耳盘村;在随机干扰条件下的增长性参数V′pi排序为陈家湾村>河珠村>干耳盘村>绒春村;对非随机干扰的敏感性参数P″max排序为陈家湾村>绒春村>河珠村>干耳盘村;非随机干扰条件下种群的增长性参数V″pi排序为陈家湾村>绒春村>河珠村>干耳盘村。总体上,4个种群在面对外界干扰的情况下,都是呈增长状态,但是增长速度缓慢,并且陈家湾村种群在受干扰下敏感度高于其他3个种群。
2.2 红豆树种群静态生命表及存活曲线
2.2.1 静态生命表 由表3可知,红豆树种群不同龄级个体数差别较大,种群期望寿命ex随着龄级增长呈逐渐降低趋势,河珠村种群期望寿命ex表现出两个高峰,分别在第Ⅰ龄级和第Ⅳ龄级。4个种群的死亡率(qx)和消失率(Kx)变化各有不同。河珠村种群的死亡率和消失率在第Ⅰ龄级和第Ⅱ龄级维持在较低水平,在第Ⅲ龄级达到第一个峰值后下降,随后在第Ⅵ龄级达到第二个峰值,说明该种群在生长中期受到一定程度上的人为干扰和环境压力;陈家湾村和绒春村种群qx和Kx在第Ⅰ龄级较高,在第Ⅱ龄级急速降至最低水平后,分别于第Ⅴ龄级和第Ⅵ龄级出现峰值,表明陈家湾村和绒春村2个种群在生长初期受到一定的生存压力,幼龄个体死亡率较高,需要加强对幼龄个体的保护;干耳盘村种群qx和Kx变化呈现逐步攀升的变化趋势,表明该种群处于较为稳定的状态。
2.2.2 存活曲线分析 从图3可以看出,随着龄级的增加,4个红豆树种群标准化存活数对数(lnlx)逐渐减小,陈家湾村、干耳盘村和绒春村存活曲线总体上呈对角线型,但河珠村种群呈阶梯型,在中龄级个体波动较大。分别采用指数模型Nx=N0e-bx和幂函数方程Nx=N0x-b检验存活曲线类型(表4),各种群指数模型的R2值和F值均大于幂函数模型,4个种群存活曲线均符合Deevey- Ⅱ。
2.3 红豆树种群生存分析
种群生存函数如图4所示,随着龄级的增加,红豆树种群生存率S(i)呈逐步降低的趋势,累计死亡率F(i)呈逐步增加的趋势。河珠村和陈家湾村种群在第Ⅱ至第Ⅲ龄级之间出现两者的平衡点,表明两地红豆树种群在Ⅲ龄级之后就开始进入衰退期。而在干耳盘村和绒春村,两者平衡点出现于更早的龄级,分别在第Ⅱ龄级和第Ⅰ至第Ⅱ龄级之间,说明干耳盘村和绒春村红豆树种群相比河珠村和陈家湾村种群更早进入种群衰退期。
红豆树种群危险率函数λ(ti)呈递增的趋势,在河珠村,种群于第Ⅱ至第Ⅲ龄级之间出现危险率骤增的情况。红豆树种群死亡密度函数f(ti)走势较稳,基本保持在0.2以下,在第Ⅰ至第Ⅱ龄级种群个体数骤减。通过4种生存函数的分析发现,4个红豆树种群数量动态变化表现为幼龄级个体死亡率高,成熟龄级个体数量稳定的特点。
2.4 红豆树不同种群数量动态的时间序列预测分析
时间序列预测结果如图5所示,河珠村种群第Ⅱ龄级个体数在经历2个龄级时间后减少了15.8%,经历4个、6个龄级时间后各龄级个体数均有不同程度的增加。陈家湾村种群在经历2个龄级时间后,第Ⅱ龄级个体数有明显增长,经历4个、6个龄级后,第Ⅲ至第Ⅵ龄级个体数量有不同程度的减少,第Ⅶ龄级数量先增加后减少。干耳盘村种群在经历2个、4个、6个龄级时间后,各龄级个体数均有不同程度的增加,其中第Ⅵ龄级数量变化稍有波动。绒春村种群在经过2个、4个、6个龄级后,第Ⅱ龄级个体数量有明显的增加,第Ⅲ至第Ⅵ龄级个体数无明显变化,第Ⅶ龄级个体数逐步减少。
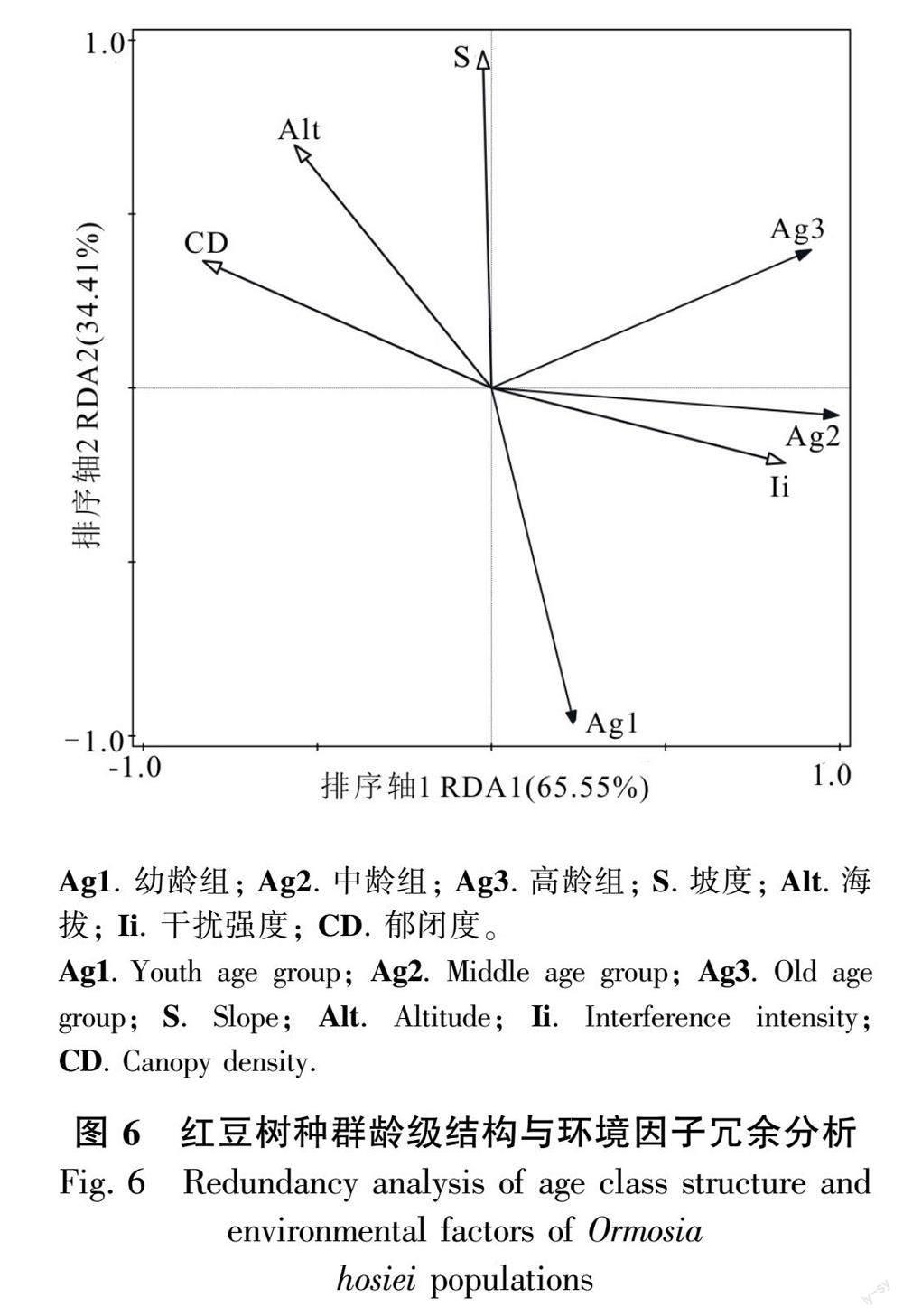
2.5 环境因子对红豆树种群结构的影响
RDA排序结果(图6)显示,排序轴RDA1与排序轴RDA2两轴对不同龄组个体数量变异的解释率分别为65.55%与34.41%,前两个排序轴对个体数量变异的解释率达99.96%,解释效果显著(F=5.2,P<0.05)。其中,干扰强度(R2=0.826,P=0.012)与第一典范轴相关性最高,其次是郁闭度(R2=-0.800,P=0.028), 两者对种群数量结构变异的解释率分别为48.2%与11.3%;坡度(R2=0.042,P=0.008)与第二典范轴相关性最高,对种群数量结构变异解释率为33%。从环境因子对不同龄组个体数量变异解释上来看:坡度和海拔是限制幼龄个体发展的主导因子;郁闭度和海拔对红豆树种群个体数量起负向影响,其中郁闭度对中龄个体数量的影响较大;适度的人为干扰对红豆树中、高龄个体起正向影响,其中对中龄个体的影响较为明显。
3 讨论与结论
3.1 红豆树种群龄级结构及数量动态变化趋势
物种自身的生物学、生态学特性及周围环境因子对种群特征有着重要的影响作用(张文辉等,2005)。红豆树种群龄级结构显示,4个种群均呈现幼龄个体数量比例较高的特点,表明种群自我繁殖能力较强,具有较高的增长潜能, 这与红豆树结实间隔期长,但母树单次结实量大的繁殖特性有关(刘鹏等,2017)。从不同种群龄级结构差异来看,河珠村和干耳盤村种群龄级结构呈现倒“J”型,陈家湾村和绒春村种群龄级结构均有不同程度缺失,呈不规则“哑铃”型结构。其中,河珠村种群第Ⅲ龄级个体死亡率和消失率最高, 而陈家湾村和绒春村种群第Ⅰ龄级和第Ⅱ幼龄级个体死亡率和消失率最高。可以看出,幼龄个体数量波动及中龄个体数量缺失是导致不同种群龄级结构差异的主要原因,其影响因素有以下三点。(1)红豆树种子传播特性:由于红豆树种子重量大,千粒重达1 310 g(罗旋等,2021),空间扩散能力差,分布于山区以及隔河流较远的红豆树种群缺乏流水这一重要的传播媒介,更新幼苗集中分布于母树周围,幼苗存在激烈的种内竞争,一定程度上限制了幼龄个体向中龄个体发展,是导致陈家湾村和绒春村种群第Ⅰ龄级和第Ⅱ龄级个体死亡率和消失率都高的重要因素。(2)红豆树的生长习性:红豆树幼苗的生长需要适当的荫蔽环境(潘惠均,2014),但当个体发展到中龄阶段以后,对光照的需求逐渐增强,较高的郁闭度使得红豆树中龄个体取光不足,是导致河珠村种群中龄个体发展受限的重要原因。(3)外界干扰:中龄级个体数量缺失的原因一方面可能与随着个体长大导致的种内和种间激烈竞争有关,另一方面可能与遭受破坏有关(黄冬柳等,2019)。虽然红豆树被列为国家二级保护树种,但调查中发现当地政府和居民只注重红豆树古大树的保护,而忽略了对其种群的保护。在红豆树种群中,幼龄个体一般会被老百姓忽略,古大树老百姓不敢砍,中龄个体则成为樵采、盗伐的对象,这也是导致红豆树种群生境破碎化,种群数量不断减少的重要原因。此外,调查中发现红豆树有着较强的萌蘖能力(李峰卿等,2017),陈家湾村、绒春村2个种群分别有27.7%、42.8%的幼苗个体来自于萌蘖更新,显然是由于外界破坏导致部分中龄级个体死亡后由树桩萌发产生的幼苗,中龄个体的缺失一定程度上激发了红豆树种群的自我调节能力。种群动态变化分析证实了这一观点,种群在受外界随机干扰的情况下增长性参数排序V′pi为陈家湾村>河珠村>干耳盘村>绒春村,说明在受到外界轻微干扰的情况下,红豆树种群通过负反馈调节激发更高的增长潜能,而过度的破坏则会使红豆树种群丧失这种自我恢复能力,这与张群芳等(2015)对四川省万源市红豆树种群结构的研究结果类似。生存分析与时间序列分析能直观地体现红豆树种群结构特征与发展动态(张晓鹏等,2022; 李敏敏等,2022),川黔地区红豆树种群数量动态变化整体表现为幼龄级个体死亡率高,成熟龄级个体数量稳定的特点,中龄级个体受幼龄个体数量变化的影响,在不同种群中出现较大的结构差异。从时间序列预测分析也可以看出,红豆树种群在未来2个、4个、6个龄级后,幼龄龄级个体数变化较为明显,其他龄级个体数变化相对平稳。幼龄个体数量结构的变化是种群演替发展的核心,因此,如何针对红豆树种群生长特性和生境特点进行幼龄级个体的保护和管理是促进红豆树种群自然恢复和更新的关键。
3.2 制约红豆树种群不同龄级个体发展的关键因素
种内、种间竞争及周围环境对种群个体的发展均有至关重要的影响(吴邦利等,2018)。本研究中RDA排序结果表明,坡度、海拔是影响幼龄级个体发展的关键因素,与幼龄个体呈负相关。红豆树幼龄个体不仅受到个体间竞争的影响,外部环境也制约着幼龄个体的发展。坡度较大的山区,红豆树散落的种子和萌发的幼苗易因坡度过大而向下坡迁移,尤其是受雨季滑坡的影响,导致种子和幼苗失去原有的生长环境而不能幸存(黄萍和刘艳红,2018);随着海拔的升高,温度降低,不利于红豆树种子萌发和幼苗生长,特别是西南山区每年冬天都会出现不同程度的凝冻灾害,导致一些幼苗不能越冬。此外,适度的人为干扰对红豆树种群中龄组和高龄组个体有着正向影响,适度的人为干扰可以调节种群密度,能为种群保留个体留出足够的生存空间(张群芳等,2015)。可见,在保证不破坏红豆树自然种群生境的前提下,适当进行种群密度调整,是稳定红豆树中高龄级个体发展的有效措施。
3.3 红豆树种群保护和恢复策略
研究濒危物种的种群特征是了解植物濒危机制的重要基础,是濒危植物保护和恢复的理论依据(吴其超等,2021)。种群结构特征受多方面因素的影响,其中坡度、海拔、人为干扰是影响红豆树种群结构特征的主导因子,其自身的生物学特性也是种群结构特征的重要影响因素(田慧霞等,2017)。在本研究中,川黔山区红豆树种群因远离流域,种子缺乏传播媒介,导致种群发展受到限制,4个区域红豆树种群均显示出幼龄个体密度较大的特点。由于红豆树自身的生物学特性,幼龄个体容易聚集分布于母树周围,密度过大和死亡率较高,建议采取人工移栽以减少幼苗密度,提高幼龄个体存活率。同时,进行人工采种培育,开展野外回归以促进红豆树种群的恢复和自然更新(任海等,2014; 丁剑敏等,2018)。陈家湾村和绒春村种群呈现出中龄个体“缺失”的种群结构,针对种群中龄个体容易受到外界破坏的特点,采取相应的保护措施,如建立自然保护区,加强对红豆树保护的宣传力度,增强老百姓的保护意识,避免对其生境造成再破坏(李丹等,2016)。红豆树种群具有一定的自我调节能力,当种群密度较大的时候,对中龄个体进行适度的人为干预,调节生长空间能稳定种群数量并保持其旺盛的生长。此外,调查中发现绒春村种群还存在红豆树古大树被过度保护的现象,即对大树周围建立树池、硬化路面,严重限制其根系呼吸和养分吸收,导致其生长衰退。针对此类现象应积极和当地政府沟通,及时拆除以保护为名义建立的树池及硬化铺装,并实施复壮等措施。
参考文献:
BI SHT VK, KUNIYAL CP, NAUTIYAL BP, et al., 2013. Spatial distribution and regeneration of Quercus semecarpifolia and Quercus floribunda in a subalpine forest of western Himalaya, India [J]. Physiol Mol Biol Plants, 19(3): 443-448.
CAI Y, 2007. Preliminary study on seedling technology of Ormosia hosiei in western Hubei [J]. For Prospect Des, (2): 97-100. [蔡勇, 2007. 鄂西红豆树育苗技术初步研究 [J]. 林业勘察设计, (2): 97-100.]
CHEN XD, 1998. A study on the method of quantitative analysis for plant population and community structural dynamic [J]. Acta Ecol Sin, 18(2): 104-107. [陈晓德, 1998. 植物种群与群落结构动态量化分析方法研究 [J]. 生态学报, 18(2): 104-107.]
DING JM, ZHANG XD, LI GL, et al., 2018. Genetic considerations in recovery of endangered plants [J]. Plant Sci J, 36(3): 452-458. [丁剑敏, 张向东, 李国梁, 等, 2018. 濒危植物居群恢复的遗传学考量 [J]. 植物科学学报, 36(3): 452-458.]
GAO L, L ZP, LE JX, et al., 2019. Research progress of conservation biology on Torreya fargesii, a rare and endangered plant endemic to China [J]. J Chongqing Norm Univ(Nat Sci Ed), 36(2): 130-135. [高岚, 吕治平, 乐佳兴, 等, 2019. 中国特有裸子植物巴山榧树的保护生物学研究进展 [J]. 重庆师范大学学报(自然科学版), 36(2): 130-135.]
HETT JM, LOUCKS OL, 1976. Age structure models of balsam fir and eastern hemlock [J]. J Ecol, 64(3): 1029-1044.
HUANG DL, LIANG SC, JIANG Y, 2019. Quantitative dynamics of Cyclobalanopsis glauca population in Guilin karst mountainous areas [J]. Chin J Ecol, 38(3): 663-671. [黃冬柳, 梁士楚, 姜勇, 2019. 桂林岩溶石山青冈种群数量动态 [J]. 生态学杂志, 38(3): 663-671.]
HUANG P, LIU YH, 2018. Effects of stand structure and terrain factors on seedling regeneration of Pinus tabuliformis forest in the Songshan National Nature Reserve, Beijing [J]. Chin J Ecol, 37(4):1003-1009. [黄萍,刘艳红, 2018. 北京松山油松林林分结构和地形对幼苗更新的影响 [J]. 生态学杂志, 37(4): 1003-1009.]
JIN H, ZHAO Y, LIU LJ, et al., 2019. Quantitative characteristics and population dynamics of the endangered plant Thuja koraiensis in Changbai Mountain, China [J]. Chin J Appl Ecol, 30(5): 1563-1570. [金慧, 赵莹, 刘丽杰, 等, 2019. 长白山区濒危植物朝鲜崖柏种群数量特征及动态 [J]. 应用生态学报, 30(5): 1563-1570.]
LI D, ZHANG XR, YANG XB, et al., 2016. Exploration of the protective effect on endangered plant populations inside the nature reserve — a case study of Vatica mangachapoi in Changjiang,Hainan [J].For Resour Manag, (1):118-125. [李丹, 张萱蓉, 杨小波, 等, 2016. 自然保护区对濒危植物种群的保护效果探索——以海南昌江县青梅种群为例 [J]. 林业资源管理, (1): 118-125.]
LI FQ, CHEN HW, ZHOU ZC, et al., 2018a. Analysis on variation of seed and seedling traits of superior trees of Ormosia hosiei and primary screening of optimum family [J]. J Plant Resour Environ, 27(2): 57-65. [李峰卿, 陈焕伟, 周志春, 等, 2018a. 红豆树优树种子和幼苗性状的变异分析及优良家系的初选 [J]. 植物资源与环境学报, 27(2): 57-65.]
LI FQ, ZHOU ZC, CHU XL, et al., 2018b. Analysis of genetic diversity among progenies from plus trees of Ormosia hosiei and its correlation with growth [J]. For Res, 31(1): 42-50. [李峰卿, 周志春, 楚秀丽, 等, 2018b. 红豆树优树子代遗传多样性及与生长相关性分析 [J]. 林业科学研究, 31(1): 42-50.]
LI FQ, ZHOU ZC, XIE YJ, 2017. Genetic diversity and genetic differentiation of different populations of Ormosia hosiei in three watersheds [J]. Mol Plant Breed, 15(10): 4263-4274. [李峰卿, 周志春, 谢耀坚, 2017. 3个小流域红豆树天然居群的遗传多样性和遗传分化 [J]. 分子植物育种, 15(10): 4263-4274.]
LI J, ZOU L, L YM, et al., 2019. Population structure and dynamic characteristics of narrowly distributed endemic species Jasminum yuanjiangense [J]. Guihaia, 39(9): 1271-1280. [李娟, 邹琳, 吕亚媚, 等, 2019. 狭域特有植物元江素馨的种群结构及动态特征 [J]. 广西植物, 39(9): 1271-1280.]
LIU P, QUE SQ, LIU LT, et al., 2017. Research status and endangered conservation strategy of Ormosia hosiei [J]. Subtrop Plant Sci, 46(1): 96-100. [劉鹏, 阙生全, 刘丽婷, 等, 2017. 红豆树研究现状及濒危保护建议 [J]. 亚热带植物科学, 46(1): 96-100.]
LI MM, LIU PC, KONG WM, et al., 2022. Population structure and dynamic characteristics of the endangered Pseudotsuga forrestii Craib [J]. Acta Ecol Sin, 42(13): 1-12. [李敏敏, 刘鹏程, 孔维民, 等, 2022. 濒危植物澜沧黄杉种群结构及动态特征 [J]. 生态学报, 42(13): 1-12.]
LI W, ZHANG GF, 2015. Population structure and spatial pattern of the endemic and endangered subtropical tree Parrotia subaequalis (Hamamelidaceae) [J]. Flora, 212: 10-18.
LI W, WANG RX, ZHANG GF, 2014. Point pattern analysis of ex situ population of Taxus wallichiana var. mairei [J]. Chin J Ecol, 33(1): 16-22. [李伟, 王瑞雪, 张光富, 等, 2014. 南方红豆杉迁地保护种群的点格局分析 [J]. 生态学杂志, 33(1): 16-22.]
LI Y, DUAN X, YANG SY, et al., 2021. Preliminary study on population ecological characteristics of national key protected wild plant Ormosia hosiei in Zigui County [J]. Hubei For Sci Technol, 50(1): 6-9. [李杨, 段翔, 杨诗雨, 等, 2021. 秭归县国家重点保护野生植物红豆树种群生态特征初步研究 [J]. 湖北林业科技, 50(1): 6-9.]
LUO X, WEI XL, LUO M, et al., 2021. Effects of light substrates formula on growth and physiology of Ormosia hosiei container seedlings [J]. J Cent S Univ For Technol, 41(2): 72-79. [罗旋, 韦小丽, 骆漫, 等, 2021. 轻基质配方对红豆树容器苗生长和生理的影响 [J]. 中南林业科技大学学报, 41(2): 72-79.]
MA CC, DAI J, XIAO ZQ, et al., 2017. Community structure and distribution of minimum population species of Myristica yunnanensis [J]. Guihaia, 37(6): 783-790. [马晨晨, 代俊, 肖之强, 等, 2017. 极小种群物种云南肉豆蔻的群落结构及其种群现状 [J]. 广西植物, 37(6): 783-790.]
MENG XS, WEI XL, 2011. Life table and survival analysis on Ormosia wild endangered populations [J]. Seed, 30(7): 66-68. [孟宪帅, 韦小丽, 2011. 濒危植物花榈木野生种群生命表及生存分析 [J]. 种子, 30(7): 66-68.]
PAN HJ, 2014. Effects of stand closure on seedling growth of Phoebe chekiangensis and Ormosia hosiei under fir forests [J]. J Agric Sci, (8): 1206-1208. [潘惠均, 2014. 林分郁闭度对杉木林下浙江楠和红豆树幼苗生长的影响 [J]. 浙江农业科学, (8): 1206-1208.]
QIU HJ, SUN JJ, XU D, et al., 2020. The distribution dynamics of Ormosia hosiei under different climate change scenarios since the Last Glacial Maximum [J]. Acta Ecol Sin, 40(9): 3016-3026. [邱浩杰, 孙杰杰, 徐达, 等, 2020. 末次盛冰期以来红豆树在不同气候变化情景下的分布动态 [J]. 生态学报, 40(9): 3016-3026.]
REN H, JIAN SG, LIU HX, et al., 2014. Advances in field return research on rare and endangered plants [J]. Sci China Life Sci, 44(3): 230-237. [任海, 简曙光, 刘红晓, 等, 2014. 珍稀濒危植物的野外回归研究进展 [J]. 中国科学: 生命科学, 44(3): 230-237.]
SU XQ, 2000. Size class and distribution pattern of Castanopsis fissa population in different successional stages [J]. Chin J Appl Environ Biol, 6(6): 499-504. [蘇小青, 2000. 不同演替阶段中黧蒴栲种群的大小结构与分布格局 [J]. 应用与环境生物学报, 6(6): 499-504.]
TANG F, ZOU TC, YANG NK, et al., 2022. Population structure and dynamics analysis of rare and endangered plant Camellia kweichowensis [J]. Guihaia, 42(3): 520-529. [唐凤, 邹天才, 杨乃坤, 等, 2022. 稀有濒危植物贵州红山茶种群结构及数量动态变化的研究 [J]. 广西植物, 42(3): 520-529.]
TIAN HX, LI JM, BI RC, et al., 2017.Betula platyphylla population structure and its spatial distribution pattern in Taiyue Mountain of Shanxi, China [J]. Chin J Ecol, 36(1): 1-10. [田慧霞, 李钧敏, 毕润成, 等, 2017. 山西太岳山白桦种群结构和空间分布格局 [J]. 生态学杂志, 36(1): 1-10.]
TUO F, LIU XD, HUANG DL, et al., 2021. Quantitative dynamics of Picea crassifolia population in Dayekou basin of Qilian Mountains [J]. Acta Ecol Sin, 41(17): 6871-6882. [拓锋, 刘贤德, 黄冬柳, 等, 2021. 祁连山大野口流域青海云杉种群数量动态 [J]. 生态学报, 41(17): 6871-6882.]
WANG MS, YUAN CJ, AN MT, et al., 2016.Community characteristic and population structure of Ormosia saxatilis in Guizhou Province [J]. J W Chin For Sci, 45(1): 81-87. [王茂师, 袁丛军, 安明态, 等, 2016. 贵州濒危树种岩生红豆森林群落特征及种群结构 [J]. 西部林业科学, 45(1): 81-87.]
WANG YT, HUANG ZH, WANG J, et al., 2021. The population structure and dynamic characteristics of Phellodendron amurense in Yanshan Mountains [J]. Acta Ecol Sin, 41(7): 2826-2834. [王泳腾, 黄治昊, 王俊, 等, 2021. 燕山山脉黄檗种群结构与动态特征 [J]. 生态学报, 41(7): 2826-2834.]
WANG J, YAO L, AI XR, et al., 2020. Structure and dynamic characteristics of Betula luminifera populations in different regions of Southwest Hubei Province, China [J]. Chin J Appl Ecol, 31(2): 357-365. [王进, 姚兰, 艾训儒, 等, 2020. 鄂西南不同区域亮叶桦种群结构与动态特征 [J]. 应用生态学报, 31(2): 357-365.]
WU BL, LONG CL, QIN ST, 2018. Population structure and its quantity dynamics of Acer catalpifolium of karst forest in Maolan National Natural Reserve [J]. Acta Bot Boreal-Occident Sin, 38(10): 1918-1926. [吴邦利, 龙翠玲, 秦随涛, 2018. 茂兰喀斯特森林梓叶槭种群结构与数量动态 [J]. 西北植物学报, 38(10): 1918-1926.]
WU QC, ZANG FQ, LI CC, et al., 2021. Population structure and dynamics of endangered Populus wulianensis [J]. Acta Ecol Sin, 41(12): 5016-5025. [吴其超, 臧凤岐, 李呈呈, 等, 2021. 濒危树种五莲杨种群结构与动态特征 [J]. 生态学报, 41(12): 5016-5025.]
XU H, LIU YH, 2018. Relationship between diameter class structure and intraspecific and interspecific competitions of precious and endangering plant Acer catalpifolium [J]. Acta Bot Boreal-Occident Sin, 38(6): 1160-1170. [许恒, 刘艳红, 2018. 珍稀濒危植物梓叶槭种群径级结构与种内种间竞争关系 [J]. 西北植物学报, 38(6): 1160-1170.]
XIE MH, 2015. Structure characteristics of palaeocommunities for the natural forest of Ormosia hosiei [J]. Prot For Sci Technol, 142(7): 33-35. [谢慕华, 2015. 红豆树天然古群落结构特征的研究 [J]. 防护林科技, 142(7): 33-35.]
YOU GB, YE HJ, JIAO JJ, et al., 2017. Size class structure and spatial distribution pattern of Ormosia hosiei plantation in Baiyun Mountain, Lishui [J]. Guihaia, 37(6): 799-805. [尤根彪, 叶和军, 焦洁洁, 等, 2017. 浙江丽水白云山红豆树人工林的径级结构和空间分布格局 [J]. 广西植物, 37(6): 799-805.]
YANG FX, WANG SQ, XU HG, et al., 1991. The theory of survival analysis and its application to life table [J]. Acta Ecol Sin, 11(2): 153-158. [杨凤翔, 王顺庆, 徐海根, 等, 1991. 生存分析理论及其在研究生命表中的应用 [J]. 生态学报, 11(2): 153-158.]
YANG HQ, LIU YH, LIU FL, et al., 2020.Population structure and dynamic characteristics of an endangered and endemic species Rhododendron rex subsp. rex in southwest, China [J]. Acta Bot Boreal-Occident Sin, 40(12): 2148-2156. [杨慧琴, 刘圆缓, 刘芳黎, 等, 2020. 西南特有濒危植物大王杜鹃种群结构及动态特征 [J]. 西北植物学报, 40(12): 2148-2156.]
YANG LR, ZHANG ZL, YUN Y, et al., 2018. The population structure and dynamics of Dracaena cambodiana, an endangered tree on Hainan Island [J]. Acta Ecol Sin, 38(8): 2802-2815. [楊立荣, 张治礼, 云勇, 等, 2018. 濒危植物海南龙血树的种群结构与动态 [J]. 生态学报, 38(8): 2802-2815.]
YUAN GY, GUO QQ, YANG R, 2021.Study on the quantitative dynamics and species diversity of dominant species in forest communities in Caohai National Nature Reserve [J]. Acta Bot Boreal-Occident Sin, 41(7): 1229-1239. [袁刚毅, 郭其强, 杨瑞, 2021. 草海国家级自然保护区森林群落优势种群数量动态及物种多样性研究 [J]. 西北植物学报, 41(7): 1229-1239.]
ZANG RG, DONG M, LI JQ, et al., 2016. Conservation and restoration for typical critically endangered wild plants with extremely small population [J]. Acta Ecol Sin, 36(22): 7130-7135. [臧润国, 董鸣, 李俊清, 等, 2016. 典型极小种群野生植物保护与恢复技术研究 [J]. 生态学报, 36(22): 7130-7135. ]
ZHANG WH, GUO LJ, LIU GB, 2005.Quantity dynamics of Hippophae rhamnoides population in different habitats standing in hilly loess regions [J]. Acta Bot Boreal-Occident Sin, 25(4): 641-647. [张文辉, 郭连金, 刘国彬, 2005. 黄土丘陵区不同生境沙棘种群数量动态分析 [J]. 西北植物学报, 25(4): 641-647.]
ZHANG LJ, ZHOU WJ,NI L, et al., 2021. A review on chemical constituents and pharmacological activities of Ormosia [J]. Chin Herb, 52(14): 4433-4442. [张琳婧, 周文娟, 倪林, 等, 2021. 红豆属植物化学成分及其药理活性研究进展 [J]. 中草药, 52(14): 4433-4442.]
ZHANG QF, PENG PH, WANG J, et al., 2015. Quantitative characteristics of Ormosia hosiei under different disturbance levels [J]. Bull Bot Res, 35(5): 735-740. [张群芳, 彭培好, 王娟, 等, 2015. 不同干扰条件下红豆树种群数量特征的比较 [J]. 植物研究, 35(5): 735-740.]
ZHANG R, ZHOU Z, DU K, 2012. Genetic diversity of natural populations of endangered Ormosia hosiei, endemic to China [J]. Biochem Syst Ecol, 40: 13-18.
ZHANG XP, YU LZ, YANG XY, et al., 2022. Population structure and dynamics of Pinus koraiensis seedlings regenerated from seeds in a montane region of eastern Liaoning Province, China [J]. Chin J Appl Ecol, 33(2): 289-296. [张晓鹏, 于立忠, 杨晓燕, 等, 2022. 辽东山区天然更新红松幼苗种群结构与动态 [J]. 应用生态学报, 33(2): 289-296.]
ZOU TC, LI YY, HONG J, et al., 2021.Species diversity conservation and utilization of Guizhou rare and endangered spermatophyta [J]. Guihaia, 41(10): 1699-1717. [鄒天才, 李媛媛, 洪江, 等, 2021. 贵州稀有濒危种子植物物种多样性保护与利用的研究 [J]. 广西植物, 41(10): 1699-1717.]
( 责任编辑 周翠鸣 )
收稿日期: 2023-03-04
基金项目: 贵州省科技计划项目( [2021]一般501); 贵州省高层次创新型人才培养计划“百”层次人才项目([2016]5661)。
第一作者: 王明彬(1998-),硕士研究生,研究方向为森林培育,(E-mail)244355582@qq.com。
* 通信作者: 韦小丽,博士,教授,研究方向为森林培育,(E-mail)1229304247@qq.com。
