
玉米逆境胁迫响应基因ZmbZIP84的克隆与功能验证
2024-04-17 00:57曹丽茹庞芸芸叶飞宇马晨晨新王振华鲁晓民
核农学报 2024年4期
曹丽茹 庞芸芸 叶飞宇 马晨晨 张 新王振华 鲁晓民,*
(1河南省农业科学院粮食作物研究所,河南 郑州 450002;2神农种业实验室,河南 郑州 450002;3郑州大学农学院,河南 郑州 450002)
玉米(ZeamaysL.)在我国广泛种植,是重要的粮食作物和饲料作物,同时也是食品、化工业中不可缺少的原材料。在玉米生长周期内,所需降水量至少为250 cm[1],但我国有50%以上的玉米种植在干旱、半干旱地区,尤其在北方和西南地区经常受到干旱和高温天气的危害[2]。在植物生长发育过程中遭遇高温、干旱等非生物胁迫时,体内一系列复杂的抗逆机制会被激活,其中,转录调控在植物抵御外界非生物胁迫、影响作物正常生长发育及产量方面发挥重要作用[3-4]。
碱性亮氨酸拉链(Basic leucine zipper,bZIP)转录因子是高等植物中家族成员数量较多且功能较为复杂的转录因子家族之一[5]。据报道,bZIP 转录因子在植物抵抗生物和非生物胁迫的应激反应中发挥重要作用[6]。Li 等[7]发现玉米转录因子bZIP68 通过抑制DREB1 转录因子的冷诱导表达从而提高玉米的耐寒性。苹果(Maluspumila)转录因子MdbZIP44 通过增强MdMYB1与下游靶基因启动子的结合,促进花青素积累响应脱落酸(abscisic acid,ABA)[8]。水稻(Oryza sativa)OsbZIP52 在寒冷和干旱胁迫中起负调节作用,OsbZIP52 过表达后提高了植株对寒冷和干旱胁迫的敏感性[9];Kang 等[10]发现NaCl、聚乙二醇(polyethylene glycol,PEG)-6000 和ABA 等能够强烈诱导甘薯(Dioscoreaesculenta)转录因子IbbZIP1表达,IbbZIP1过表达后能显著提高拟南芥(ArabidopsisthalianaL.)的耐盐性和耐旱性;Ma等[11]发现玉米转录因子ZmbZIP4参与玉米根系发育,增加侧根数量、促进初生根生长,ZmbZIP4 过表达后提高植物抵抗非生物胁迫的能力;玉米ZmbZIP72 过表达后能够提高转基因拟南芥的抗旱和耐盐性,同时增强ABA诱导基因如RD29B、RAB18和HIS1-3的表达[12]。大量研究表明,bZIP转录因子广泛参与植物的生长发育并提高植物在生物及非生物胁迫下的耐受力,因此,挖掘bZIP转录因子家族中能够响应相关胁迫的基因对提高植物在逆境中的耐受性具有重要的意义。
鉴于此,本研究对三叶期玉米幼苗进行正常灌溉、干旱胁迫及复水处理,分别取处理下的叶片和根系送转录组测序;分析转录组数据,筛选核心节点基因,克隆ZmbZIP84并进行生物信息学分析,探究其不同组织及逆境胁迫下的表达模式,利用CRISPR/Cas9 技术获得拟南芥纯合突变体植株bzip84-1、bzip84-2、bzip84-3,分析在高温、干旱胁迫下野生型(Columbia,Col)和bzip84-1、bzip84-2、bzip84-3植株的表型,初步探究该基因的功能。本研究旨在探究bZIP转录因子在玉米逆境胁迫和生长发育中的作用,为培育抗逆玉米品种提供基因资源。
1 材料与方法
1.1 试验材料处理及试验设计
选取大小一致、颗粒饱满的玉米自交系郑8713 为试验材料,人工气候长日照培养箱(28 ℃、16 h光照/8 h黑暗)进行培养。待幼苗生长两周后,选择生长整齐一致的三叶期幼苗进行干旱-复水胁迫处理。利用SYSWSD 土壤温湿度测定仪(辽宁赛亚斯科技有限公司)测量土壤水分含量,干旱处理时土壤含水量为45%~50%,复水处理时土壤含水量>95%。取玉米胁迫处理前、干旱5 d、复水3 d 的叶片和根,分别命名为T0Y、T5dY、TR3dY 和T0G、T5dG、TR3dG,并提取RNA,于-80 ℃保存。5 株混为一个样品,每个样品各3 个生物学重复。
1.2 ZmbZIP84基因的克隆
将玉米自交系B73播种于河南省农业科学院原阳基地,每行种植16株,行长4 m,行距60 cm,株距25 cm,共种植6行,且最大田间持水量一直保持在80%以上。以B73 幼苗的cDNA 为模板,用基因特异性引物(ZmbZIP84-F:5′-ATGGCTTCCTCCAGCGGGA-3′ ;ZmbZIP84-R:5′-CGAGTACCAGCTCTGCTAG-3′)和高保真P505 酶进行基因扩增。反应体系共50 μL:高保真P505 酶1 μL、10 μmol·L-1正反引物各2 μL、dNTP Mix 1 μL、2×Phanta Max Buffer 25 μL、100 ng·μL-1cDNA 1 μL、ddH2O 18 μL。反应程序为:95 ℃预变性5 min;95 ℃变性15 s,56 ℃退火15 s,72 ℃延伸20 s,35个循环;72 ℃终延伸5 min。聚合酶链式反应(polymerase chain reaction,PCR)产物用1%琼脂糖凝胶电泳检测,并利用胶回收试剂盒(北京天根生化科技有限公司)回收目的条带。目的产物连接PMD18-T载体,将阳性菌落进行菌液PCR 检测,把检测到目的片段的菌液送到北京六合华大基因科技有限公司(青岛)测序。
1.3 ZmbZIP84基因的生物信息分析
使用Cytoscape3.8.2 软件构建基因共表达网络图。利用ExPasy ProtParam 网站(https://web.expasy.org/protparam/protparam-doc.html)对ZmbZIP84基因编码的蛋白质的分子质量、疏水性及蛋白稳定性等进行预测;利用MEGA11软件构建ZmbZIP84蛋白与不同物种中的同源蛋白进行进化树分析;利用MEME 在线工具(http://meme-suite.org/tools/meme)对蛋白的保守基序(motif)进行分析,把基序的最大数值设置为6。
1.4 RNA的提取及cDNA的合成
取0.1 g 玉米组织样品,液氮中磨碎,用TRIzol 法提取试剂盒(Invitrogen,北京)提取总RNA。用NANODROP 2000 浓度测定仪(Thermo,北京)检测浓度和纯度,1%琼脂糖凝胶电泳检测总RNA 的质量。取1 μg 的总RNA 用Prime-ScriptTMRT reagent Kit with gDNA Eraser(TaKaRa,北京)反转录成cDNA,稀释至终浓度100 μg·μL-1作为基因扩增和实时荧光定量PCR(quantitative real-time PCR,qRT-PCR)模板。
1.5 qRT-PCR样品及试验处理
以田间正常生长(生长条件同1.2)的玉米自交系B73 为材料,取三叶期的幼根、幼茎、幼叶以及成熟期的根、茎、叶、雄穗、雌穗等不同组织的样品进行表达模式分析。取0.1 g 组织样品于液氮中磨碎,TRIzol 法提取试剂盒提取总RNA。浓度测定仪检测其浓度和纯度,1%琼脂糖凝胶电泳检测总RNA 的质量,取1 μg的总RNA 用Prime-ScriptTMRT reagent Kit with gDNA Eraser反转录成cDNA,稀释至终浓度100 μg·μL-1。根据目的基因的保守性,利用NCBI网站设计特异性引物(ZmbZIP84-F:5′-AGAAAGGGCGGTTCTTGCG-3′;ZmbZIP84-R:5′-AAGCGGATGCTGTCGAACC-3′)。以稀释后的cDNA为模板,利用Hieff qPCR SYBR Green Master Mix(上海翊圣生物科技有限公司)进行qRTPCR 扩增,反应体系共20 μL: Hieff qPCR SYBR Green Master Mix10 μL、正反引物各引物0.4 μL、100 ng·μL-1cDNA 1 μL、ddH2O 8.2 μL。以玉米18S(18S-F:5′-C CTGCGGCTTAATT GACTC-3′;18S-R:5′-CCAGACCT CAGCCTGCTAAC-3′)为内参,反应程序为:95 ℃预变性5 min;95 ℃变性10 s,60 ℃退火30 s,40 个循环。采用2-△△CT法计算基因的相对表达量[13],每个处理3 个重复。
1.6 非生物胁迫试验处理
以饱满的玉米自交系B73 为材料,于人工气候长日照培养箱(28 ℃,16 h 光照/8 h 黑暗)进行培养。待幼苗生长2 周后,选择长势一致的玉米三叶期幼苗进行胁迫处理。干旱胁迫:利用20% PEG-6000(Holland营养液+20% PEG)进行处理[14];高温胁迫:将温度升高至42 ℃进行处理[15];盐胁迫:利用200 mmol·L-1浓度的NaCl 进行处理[14];氮胁迫:利用缺乏铵态氮的Holland营养液进行处理[14]。
1.7 CRISPR/Cas9 构建拟南芥敲除载体、阳性植株的获得及抗旱性检测
查找ZmbZIP84在拟南芥中的同源基因(AT4G345 90),参照Tsutsui 等[16]的方法构建CRISPR/Cas9载体。首先登录CRISPRdirect网站(http://crispr.dbcls.jp/)设计靶点,分析gRNA结构,在Cas-OFFinder网站(http://www.rgenome.net/cas-offinder/)评估脱靶情况,筛选靶点,设计引物(F: 5′-ATTGGATTCAAACGTCGTCAGGAT-3′,R:5′- GATT CAAACGTCGTCAGGATGTTT-3′)。将引物浓度稀释至100 μmol·L-1,取正反引物各10 μL于200 μL PCR 管中混合均匀,100 ℃加热5 min,自然冷却至室温,使引物聚合为二聚体形式。利用限制性内切酶AarI(Thermo,北京)酶切PKI1.1R 载体,体系为20 μL:10×Buffer AarI 2 μL、DNA 1.5 μL、50×oligonucleotide 0.4 μL、AarI 0.6 μL、Nuclease-free water 15.5 μL。37 ℃孵育8 h,65 ℃孵育20 min,用DNA回收试剂盒(北京天根生化科技有限公司)回收线性化载体。利用T4 ligase(TaKaRa,北京)连接靶点和PKI1.1R 载体,体系为15 μL:靶点引物二聚体5.8 μL、T4 连接Buffer 1.5 μL、线性化载体1 μL、T4连接酶0.7 μL、Nucleasefree water 6 μL,22 ℃孵育30 min。将上一步连接产物转化DH5α 感受态细胞,用壮观霉素(spectinomycin,Spec)平板进行阳性克隆的筛选。用引物U6.26:5′-TGTCCCAGGATTAGAATGATTAGGC-3′和靶点反向引物进行PCR鉴定,目标条带大小约260 bp,用引物U6.26对阳性单克隆菌液测序验证。将测序验证后的阳性质粒转化农杆菌。阳性菌落经PCR 验证后利用花序浸染法浸染野生型(Col)拟南芥,成熟后收获的种子为T0代。T0代种子消毒后,涂在含有潮霉素的1/2 MS 培养基(Murashige and Skoog medium)上进行阳性苗的筛选,PCR验证为阳性的幼苗进行种植,成熟后单株收获,即T1代种子。T1代种子消毒后,涂在不含抗生素的1/2 MS 培养基上,筛选分离比符合3∶1 的分离单株,PCR验证为阳性的幼苗进行种植并收获,即T2代种子。相同的培养基再种植一代,选择不发生分离的T2代单株进行种植并收获,为纯合的T3代种子。
利用野生型拟南芥Col 和稳定纯合的T3代种子,经消毒后分别涂在无抗1/2 MS 培养基上。生长5 d后,选择长势一致的幼苗移至营养土中培养,待生长到5 片叶,进行干旱和高温胁迫(土壤含水量45%)处理一周,观察表型差异并测定生理生化指标。
1.8 ZmbZIP84基因亚细胞定位
根据玉米MaizeGDB(https://maizegdb.org)公布的ZmbZIP84基因序列,利用NCBI 设计目的基因引物(ZmbZIP84-pMDC83-GFP-SpeI-F:5′-GGACTAGTAT GGCTTCCTCCAGCGGGA-3′;ZmbZIP84-pMDC83-GFPBamHI-R:5′-CGGGATCC CAGATCAGACGTGGCGG TGA-3′)。以测序正确的ZmbZIP84-PMD18 为模板,克隆蛋白编码区(coding sequence,CDS)序列。原始载体pMDC83-GFP 经SpeI 和BamHI 双酶切将载体线性化后,利用T4连接酶将目的条带连接到带有绿色荧光蛋白(green fluorescent protein,GFP)标签的融合表达载体pMDC83上。将菌液PCR 目的片段正确的阳性菌落送到北京六合华大基因科技有限公司(青岛)测序。测序正确的菌液抽取质粒后,使用热激法将该质粒转到农杆菌感受态GV3101 中。使用注射法将菌液注射到烟草背面的叶肉细胞中,48~96 h在激光共聚焦显微镜下观察GFP绿色荧光信号。
1.9 生理指标的测定
叶绿素含量采用95%乙醇法测定[17],脯氨酸含量采用酸性茚三酮法测定[18],超氧化物歧化酶(superoxide dismutase,SOD)活性采用NBT 法测定[18],过氧化物酶(peroxidase,POD)活性采用愈创木酚法测定[18]。
1.10 统计分析
每组试验进行3 个生物学重复,每组数据3 个重复。用Excel 2010 进行数据处理并制图,用SPSS 22 对数据进行分析。
2 结果与分析
2.1 ZmbZIP84的鉴定
对转录组数据中基因每千个碱基的转录每百万映射读取的片段(fragments per kilobase of exon model per million mapped fragments,FPKM)和功能注释进行分析,发现有27 个基因参与干旱-复水胁迫响应。选取相关系数大于0.5 的基因构建共表达网络图。结果显示,ZmbZIP84基因与其他基因之间的关联度较高,表明ZmbZIP84基因是核心节点的关键基因(图1),可知ZmbZIP84基因在干旱-复水处理过程中起着重要的作用。
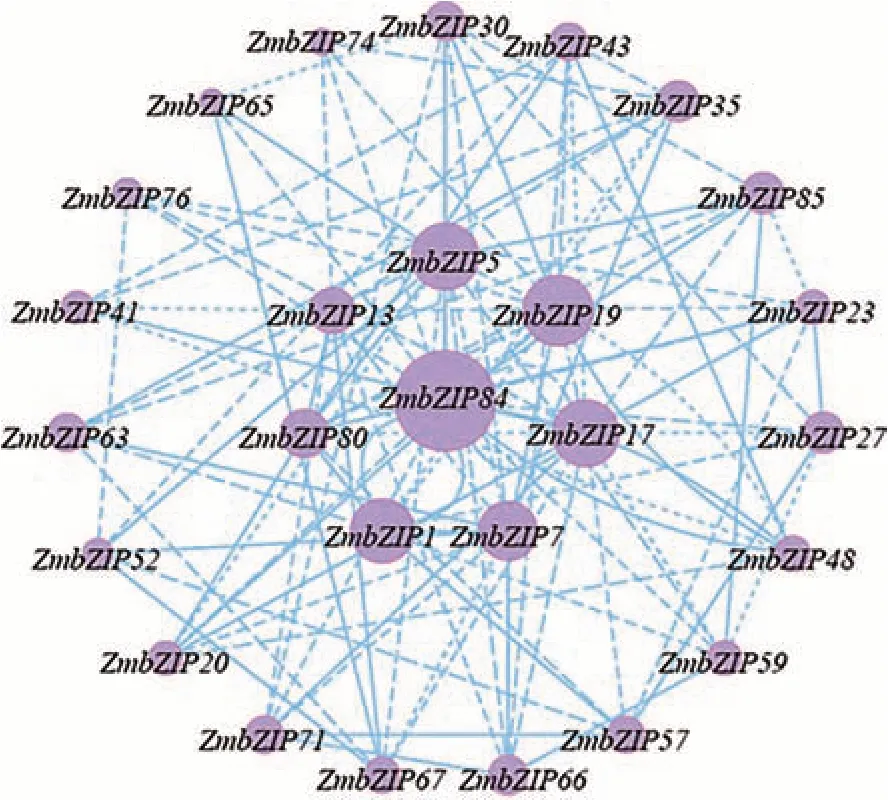
图1 差异表达基因的共表达网络Fig.1 Co-expression network of differentially expressed genes
2.2 ZmbZIP84基因的克隆及生物信息学分析
利用网站MaizeGDB 下载ZmbZIP84基因的cDNA序列,以玉米自交系B73幼苗的cDNA为模板,利用PCR技术对ZmbZIP84基因进行扩增。1.2%的琼脂糖凝胶电泳进行检测,结果得到约489 bp的条带(图2-A),回收目的条带。将胶回收产物连接PMD18-T载体,阳性菌落送到北京六合华大基因科技公司(青岛)进行测序,通过比对测序结果,发现克隆到的基因与参考序列一致。
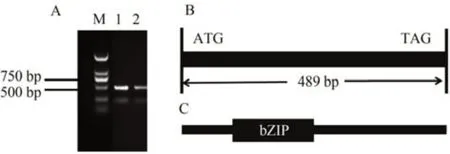
图2 ZmbZIP84 基因(A)、编码蛋白结构(B)、结构域(C)Fig.2 Gene structure of ZmbZIP84(A),coding protein structure (B) and domain (C)
对ZmbZIP84的基因序列进行分析,发现该基因位于玉米的第4 号染色体,编码区全长489 bp,编码162 个氨基酸(图2-B)。对该基因的蛋白保守结构域进行预测,显示ZmbZIP84具有典型的bZIP 转录因子结构域(图2-C)。
利用Expasy Protpara 网站对ZmbZIP84 的氨基酸序列进行理化性质的预测和分析。结果表明,分子式为C718H1182N226O235S;相对分子量为17.13 kDa,理论等电点(pI)为6.29;带负电荷的残基总数(Asp+Glu)为18,带正电残基总数(Arg+Lys)为17,为酸性蛋白;亲水性系数为-0.107,说明该蛋白是亲水性蛋白;不稳定系数为84.75,属于不稳定蛋白。对其氨基酸组成成分进行分析,其中Ala(A)含量最多,占21.6%;其次为Arg(R),占9.3%;含量最少的是Ile(I),占0.6%。
2.3 ZmbZIP84蛋白进化树分析及保守基序分析
为了研究ZmbZIP84与其他物种的亲缘关系,利用邻接法(neighbor-joining,NJ)构建系统进化树。由图3可知,bZIP 转录因子具有较高的保守性(图3-A),与ZmbZIP84 亲缘关系最近的物种为柳枝稷和高粱(图3-B)。对同源物种的6 个保守基序进行分析,结果发现这7 个物种含有相似的保守基序(如:motif 1、motif 2、motif 3、motif 4),说明高粱、哈利谷草、柳枝稷、水稻、玉米、光稃稻、粳稻物种间可能存在某种相似的生物学功能(图3-C)。
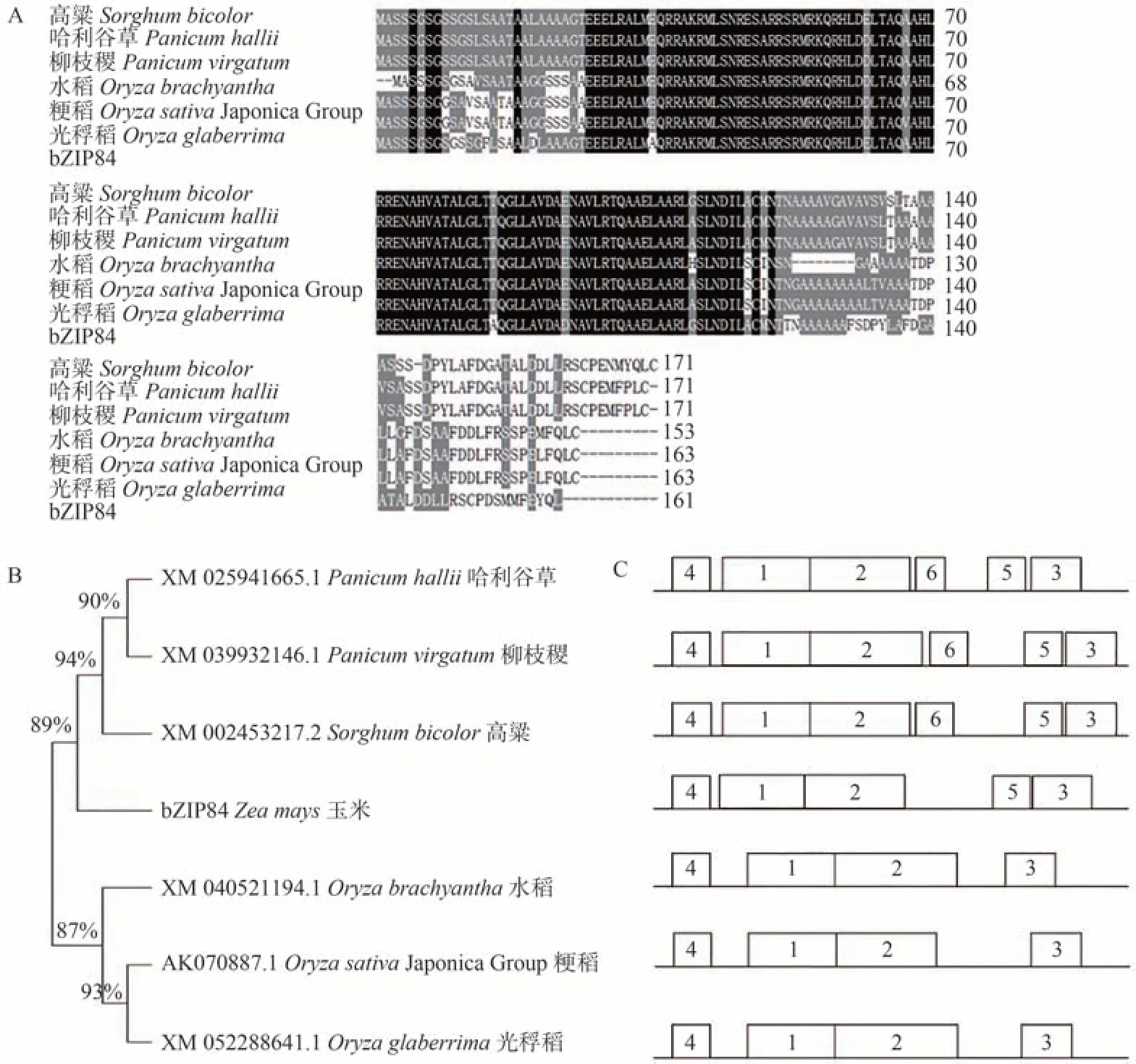
图3 ZmbZIP84与其他物种的序列(A)分析、蛋白进化树(B)及保守基序(C)分析Fig.3 Sequence analysis (A) of ZmbZIP84 and other species,protein evolutionary tree (B) and conserved motif analysis(C)
2.4 ZmbZIP84的启动子顺式元件分析
启动子顺式作用元件在基因表达的调控过程中起着重要作用。为了进一步探究ZmbZIP84的功能,利用PlantCARE在线软件对ZmbZIP84基因ATG上游2 kb启动子的顺式元件进行预测(表1)。发现ZmbZIP84基因启动子区含有多种功能元件,除了在启动子中最常见的TATA-box、CAAT-box 元件外,还包括ABRE、G-Box、Sp1、GT1-motif、CGTCA-motif、RY-element、Abox和TCCC-motif 等其他元件。其中,ABRE 参与响应ABA 的信号转导过程,能够响应干旱胁迫;TGACGmotif 和CGTCA-motif 是茉莉酸甲酯(metyl jasmonate,MeJA)的结合元件,通过参与茉莉酸甲酯的传导从而来调节茉莉酸相关基因的表达;O2-site 是参与调控玉米蛋白代谢的顺式作用元件;TGA-element 是生长素响应元件,能够提高生长素相关基因的表达。此外,还发现Box Ⅱ、Sp1、TCT-motif 等一些光刺激应答元件。结果表明,ZmbZIP84基因的表达可能被识别这些元件的转录因子所调控,从而在植物生长发育、响应激素信号刺激及逆境胁迫过程中发挥重要作用。

表1 ZmbZIP84基因启动子顺式元件分析Table 1 Cis-element analysis of ZmbZIP84 gene promoter
2.5 ZmbZIP84基因组织表达模式分析
分析ZmbZIP84基因在玉米不同时期不同组织中的表达情况。结果显示,该基因在所有检测的组织中均表达,但表达模式存在明显的差异。其中,成熟根中的表达量最高,幼根、雌穗及幼叶中的表达量次之,幼茎中表达量最低(图4)。
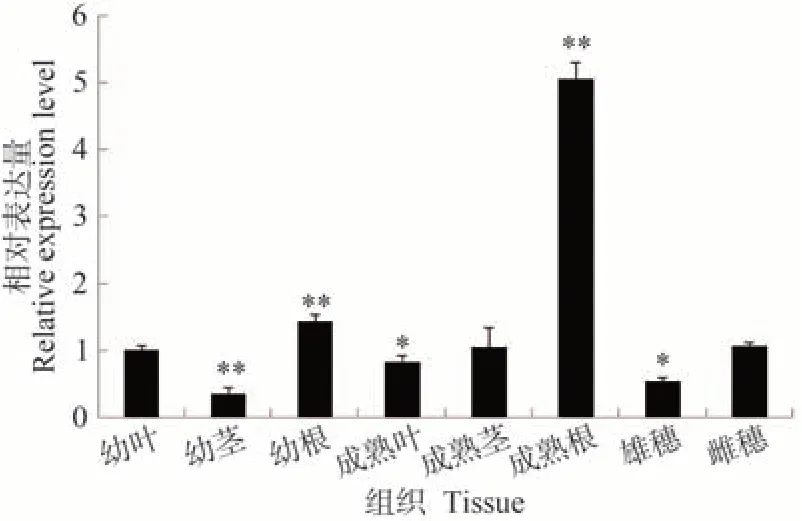
图4 ZmbZIP84基因在玉米不同组织中的表达Fig.4 The expression of ZmbZIP84 gene in different tissues of maize
2.6 ZmbZIP84 基因在非生物胁迫诱导表达模式分析
通过对ZmbZIP84基因在干旱、NaCl、高温及缺氮处理下的表达量进行分析,发现该基因在逆境胁迫下能够参与响应。结果表明,在高温、干旱及氮处理下ZmbZIP84基因的表达量显著或极显著上调,而在盐胁迫处理下该基因的表达量显著或极显著下调(图5)。在高温处理条件下(图5-A),ZmbZIP84基因的表达量显著或极显著上调,且上升趋势一直持续到48 h,此时表达量达到处理前水平的12.5 倍;之后表达量缓慢下调;恢复正常培养36 h 时,表达量降至最低,但表达量仍比胁迫前高。在干旱处理条件下(图5-B),ZmbZIP84基因的表达量变化趋势与高温处理相似;干旱处理8 h时,表达量已上调至13.7 倍;随着干旱胁迫时间的增加,ZmbZIP84基因的表达量出现缓慢下降的趋势,在胁迫52 h时,表达量又出现缓慢上调趋势;之后表达量缓慢下调至84 h;在复水处理后,随着时间的增加,表达量变化趋于稳定。在盐处理条件下(图5-C),ZmbZIP84基因的表达量呈下调趋势;盐处理36 h时,表达量为处理前水平的0.3 倍,为最低水平;解除胁迫12~36 h,表达量逐渐下调。在缺氮条件下(图5-D),ZmbZIP84基因的表达量也呈显著或极显著上调趋势;缺氮处理48 h时,表达量上调至4.52 倍;而解除胁迫后,表达量恢复至处理前水平。以上结果表明,干旱、高温、盐、缺氮处理均可诱导ZmbZIP84基因的表达。
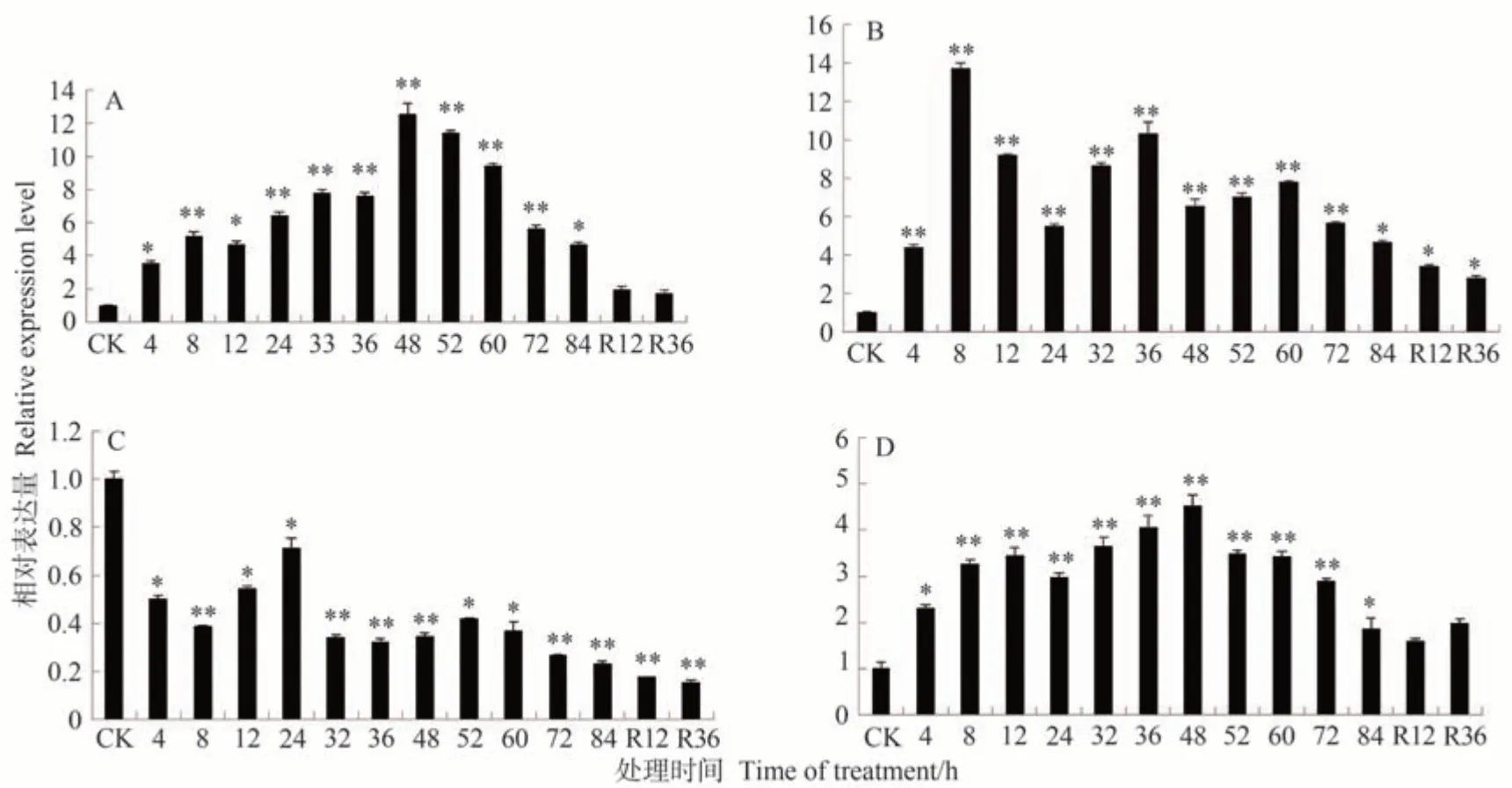
图5 ZmbZIP84基因在高温(A)、干旱(B)、盐(C)、缺氮(D)胁迫下的表达Fig.5 Expression of ZmbZIP84 gene under drought (A),high temperature (B),NaCl (C) and nitrogen deficiency(D) stress
2.7 干旱、高温胁迫下ZmbZIP84 敲除拟南芥植株的表型分析
ZmbZIP84基因与拟南芥AT4G34590的同源性最高,序列相似性达61.54%。将玉米与拟南芥同源度最高的片段利用CRISPR/Cas9 技术获得bZIP84缺失型拟南芥突变体(图6-A)。将野生型Col 和突变体T3 代的纯合株系bZIP84-1、bZIP84-2和bZIP84-3进行干旱和高温胁迫处理。在正常处理下,Col和突变体植株的长势一致且良好(图6-B、C)。干旱胁迫处理5 d后,发现突变体植株生长受到严重抑制,叶片萎蔫,甚至干死;而Col 植株影响较小,但弱于同时期正常处理的Col 植株。高温处理5 d 时,各株系均出现了较为严重的萎蔫、干枯现象;但相对于突变体植株,Col植株的萎蔫程度相对较轻。以上结果说明,在干旱、高温胁迫处理下,缺失bZIP84基因导致拟南芥抗旱性和耐高温能力明显下降。

图6 玉米ZmbZIP84基因与拟南芥AT4G34590序列分析(A)、干旱及高温胁迫下bZIP84突变体和野生型的幼苗表型(B、C)Fig.6 Sequence analysis of maize ZmbZIP84 gene and Arabidopsis AT4G34590 gene (A),seedling phenotypes of bZIP84 mutants and wild types under high temperature and drought stress (B,C)
2.8 ZmbZIP84 敲除会降低拟南芥植株对干旱和高温的耐受性
测定干旱和高温胁迫下拟南芥突变体植株bzip84-1、bzip84-2、bzip84-3中叶绿素含量、脯氨酸含量、SOD 和POD 活性均发生变化(图7),发现胁迫处理前,拟南芥植株正常生长,各指标在野生型Col和突变体植株中无显著差异。在干旱和高温胁迫后,叶绿素含量均明显下降,且突变体拟南芥植株的叶绿素含量显著低于野生型(图7-A)。
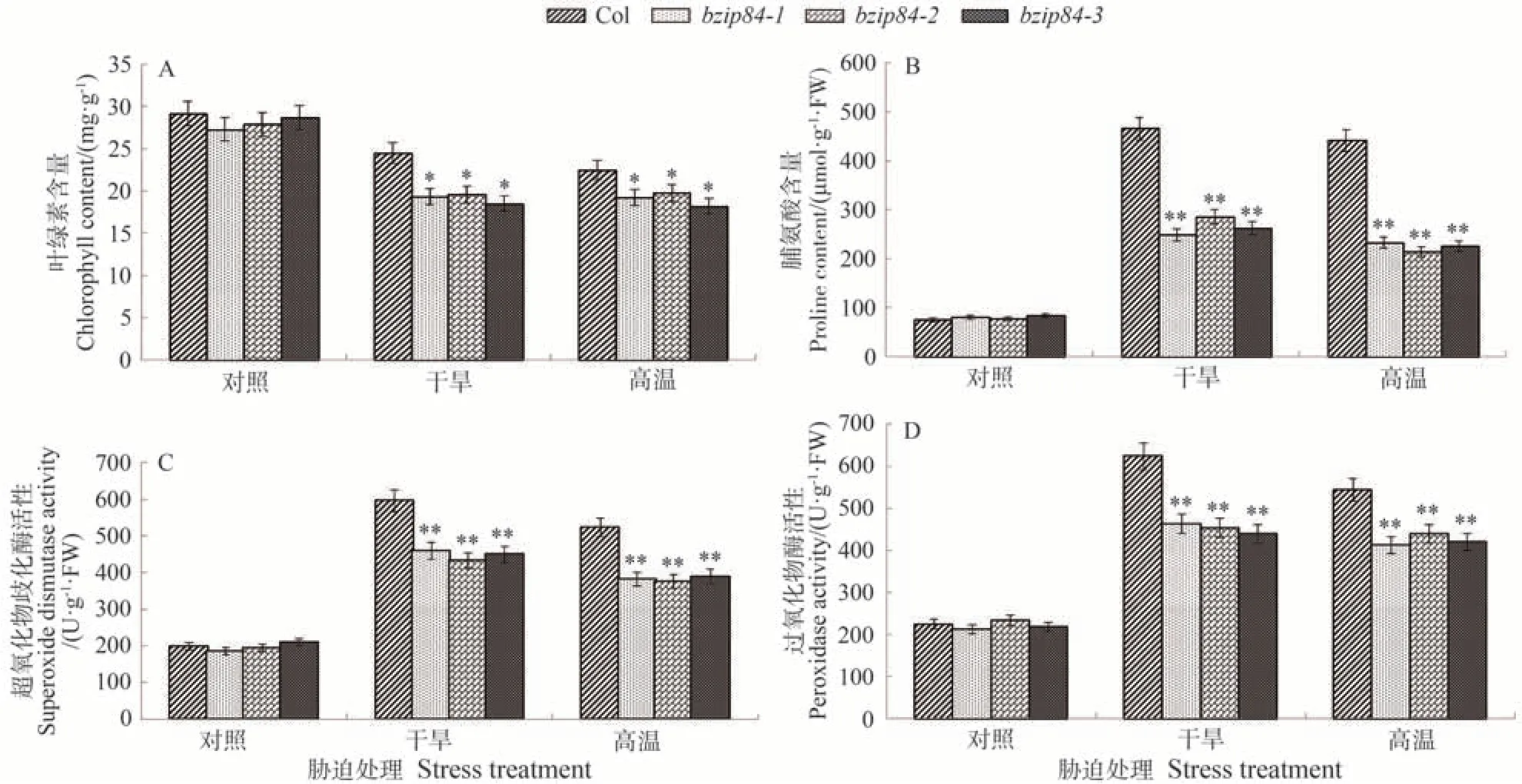
图7 干旱、高温胁迫下bZIP84突变体和野生型拟南芥生理指标测定Fig.7 Physiological indices of bZIP84 mutant and wild Arabidopsis thaliana under drought and high temperature stress
植物体内脯氨酸的含量在一定程度上能够反映植物的抗逆能力。胁迫前各株系间脯氨酸含量无明显差异;在干旱胁迫处理后,Col 株系中的脯氨酸含量是突变体株系中脯氨酸含量的1.63~1.87 倍;在高温胁迫处理后,Col株系中的脯氨酸含量是突变体株系中脯氨酸含量的1.89~2.06倍(图7-B)。
当植株遭受到胁迫时,体内的SOD 含量出现明显增加的趋势,干旱胁迫后,Col 株系中的SOD 含量比突变体株系中SOD含量分别高了29.8%、33.7%、32.5%;高温胁迫后,Col 株系中的SOD 含量比突变体株系中SOD含量分别高36.8%、38.9%、34.1%(图7-C)。在胁迫处理前,Col株系和突变体株系中的POD含量无明显差异,在胁迫后植株体内的POD 含量明显升高。干旱胁迫后,Col 株系中POD 含量比突变体株系中分别高34.7%、37.3%、41.6%;高温胁迫后,Col 株系中POD含量比突变体株系中分别高了31.7%、23.8%、29.3%(图7-D)。
通过测定生理指标,可以发现在高温和干旱胁迫后,拟南芥体内叶绿素含量下降,而脯氨酸、SOD和POD的含量上升,且Col 株系中脯氨酸、SOD 和POD 的含量高于突变体株系,说明拟南芥中缺失bZIP84基因后显著降低了拟南芥对干旱胁迫和高温胁迫的耐受性。
2.9 ZmbZIP84基因编码蛋白的亚细胞定位
为了进一步验证ZmbZIP84蛋白在细胞中的位置,将其进行亚细胞定位。结果显示,对照组在细胞核和质膜均能发出绿色荧光信号,而试验组只能在细胞核中观察到绿色荧光信号,说明该基因编码的蛋白定位于细胞核中,属于核蛋白基因(图8)。
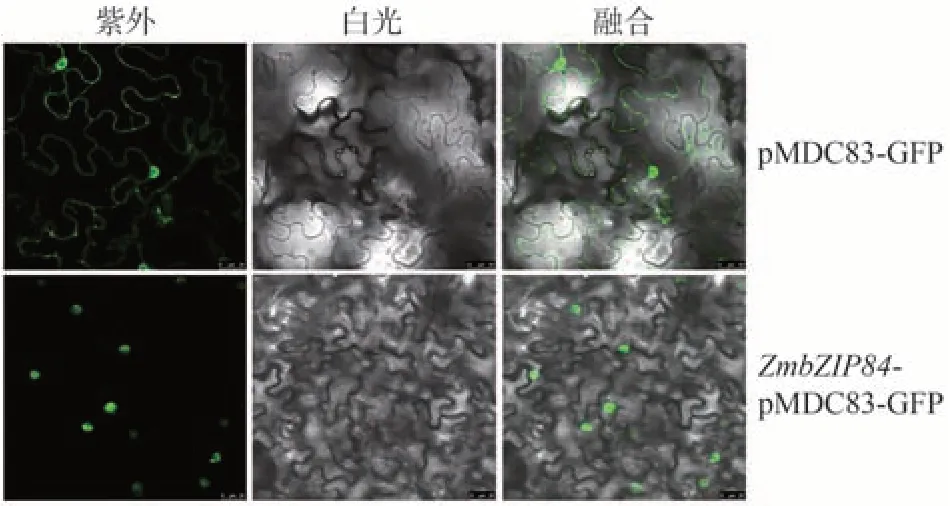
图8 ZmbZIP84的亚细胞定位Fig.8 Subcellular localization of ZmbZIP84
3 讨论
bZIP 转录因子是植物器官发育及生理过程的主要调节因子[19],在植物抗逆中发挥着重要的作用,对选育优质、绿色、高产、抗逆作物品种具有重要意义[14,20]。本研究通过分析转录组数据,在玉米中筛选到一个响应干旱胁迫及复水的转录因子ZmbZIP84,对该基因进行生物信息学分析并克隆;通过qRT-PCR 系统分析该基因在高温、干旱、缺氮及NaCl 处理下的表达模式;利用CRISPR/Cas9 技术获得拟南芥阳性纯合突变体植株,验证该基因在高温及干旱胁迫下的耐受性;通过亚细胞定位确定该基因在细胞中的位置,这些研究内容为进一步深入探究ZmbZIP84的功能及其调控的网络关系奠定了基础。
研究表明,植物bZIP 转录因子在提高作物的质量、产量及抗逆方面具有积极的作用[21]。有研究报道,ZmbZIP72基因过表达后转拟南芥,能够增加过表达植株对ABA 和渗透胁迫的敏感性,同时提高转基因株系对干旱和盐胁迫的耐受性[12];ZmbZIP81过表达后能够提高转基因拟南芥对NaCl 的耐受性[22]。本研究发现ZmbZIP84基因在玉米植株各器官中差异表达,且在高温胁迫、干旱胁迫及缺氮处理下表达量上调,而在盐胁迫下表达量下调,说明ZmbZIP84基因在玉米的逆境胁迫应答中起重要的作用,这与前人研究结果一致。为进一步研究ZmbZIP84基因的功能,以模式植物拟南芥为受体,利用CRISPR/Cas9 技术敲除ZmbZIP84基因与拟南芥同源基因中高度保守的序列获得阳性纯合突变体植株,对突变体植株分别进行高温、干旱胁迫后发现Col 株系的长势比突变体株系好,说明Col 株系比突变体株系在高温、干旱胁迫下具有较高的耐受性。进一步说明bZIP84基因缺失会降低拟南芥的抗旱、耐高温能力。
在植物抗逆性研究中,叶绿素含量、脯氨酸含量、SOD 及POD 活性等常被用作重要的检测指标,来反应植物的抗逆性能。逆境胁迫下植物体内叶绿体的结构发生改变,导致叶绿素含量及光合作用过程相关酶的活性降低[23-24];同时体内渗透调节物质如脯氨酸等含量增加,以调节细胞水势使植物适应不利环境[25-26]。本研究测定在高温、干旱胁迫下Col 和突变体植株中的叶绿素含量,发现受胁迫后植株体内的叶绿素含量显著下降,但突变体植株的叶绿素含量显著低于Col;而在高温、干旱胁迫下Col 和突变体株系中的脯氨酸含量都显著升高,且Col 株系中的脯氨酸含量是突变体株系中1~2 倍。逆境胁迫会损伤细胞膜,导致细胞内超氧阴离子及过氧化氢(H2O2)等活性氧的累积[27-28],SOD 和POD 作为植物细胞内重要的保护酶,在逆境胁迫下其活性显著增加以清除活性氧,从而维持细胞稳态[29-30]。本研究发现在高温、干旱胁迫下Col和突变体株系中SOD 和POD 酶活性显著增加,且胁迫后Col株系相比于突变体株系中的SOD 和POD 酶活性较高。结果表明,拟南芥突变体植株在高温、干旱胁迫下体内的叶绿素、脯氨酸、SOD 及POD 含量较Col 较低,从而判断ZmbZIP84基因敲除后会降低植物的抗旱、耐高温能力。
4 结论
本研究鉴定并克隆了响应干旱胁迫的基因ZmbZIP84,该基因属于bZIP 转录家族成员,是组成型表达基因,且被高温、干旱、NaCl 和氮处理诱导表达。利用CRISPR/Cas9 技术获得ZmbZIP84基因的拟南芥同源基因缺失型阳性纯合突变体植株,发现突变体植株在干旱和高温胁迫下生长状况远不如野生型。亚细胞定位显示,ZmbZIP84基因编码的蛋白属于核蛋白。本研究初步证明了bZIP84基因缺失会导致拟南芥抗旱性和耐高温能力显著下降,证明了ZmbZIP84转录因子在高温、干旱胁迫中起正向调控作用。
猜你喜欢
中老年保健(2022年3期)2022-11-21
亚热带植物科学(2022年1期)2022-05-17
科学与财富(2019年15期)2019-10-21
四川农业科技(2019年5期)2019-07-01
发酵科技通讯(2018年2期)2018-07-06
湖南林业科技(2017年1期)2017-02-06
安徽医科大学学报(2016年12期)2017-01-15
山东农业工程学院学报(2016年6期)2016-12-01
浙江柑橘(2016年1期)2016-03-11
天津医科大学学报(2015年2期)2015-12-22
