
转录组和蛋白组联合揭示甜瓜响应涝害胁迫的分子机制
2024-04-17 01:01张焕欣闫承璞李国权张新龙乐美旺朱方红刘文革
核农学报 2024年4期
张焕欣 闫承璞 李国权 张新龙 曹 娜 乐美旺朱方红 刘文革
(1江西省农业科学院园艺研究所,江西 南昌 330200;2中国农业科学院郑州果树研究所,河南 郑州 450009)
涝害是普遍存在且频繁发生的严重自然灾害,影响全球约10%的耕地,导致作物减产约15%~80%[1]。我国是受涝害影响较为严重的国家,尤其在长江中下游地区,春季经常阴雨连绵,夏季多暴雨天气,导致土壤含水量过高,因此春夏季节土壤经常处于涝渍状态[2-3]。甜瓜(CucumismeloL.)是具有重要经济价值的葫芦科作物,根系呼吸能力强,需氧量高,不耐湿涝,对涝害胁迫非常敏感,其产量和品质都会因涝害胁迫而降低,甚至导致植株死亡[3]。涝害胁迫已成为我国长江中下游地区甜瓜产业发展的制约因素之一,因此,探明甜瓜对涝害胁迫的响应机制,对生产优质高产甜瓜具有重要理论意义和应用价值。
涝害胁迫对植物的影响具有多面性,最严重的是导致低氧次生胁迫。氧气在水中的低扩散率与低溶解率导致其较难从气-水表面进入水中,难度约为气相中的320 000倍[4]。当土壤遭遇涝害胁迫时,土壤孔隙被水分填充,进而阻碍空气进入,致使供给植物根系的氧气大幅减少[5-6]。在低氧情况下,植物根系无法进行有氧呼吸作用。作为临时补救途径,植物通过无氧呼吸作用来维持细胞活性,但是长时间的无氧呼吸会影响植物生长发育必须的能量供给、中间代谢物、养分吸收等,而且无氧呼吸产生的乙醇、乙醛等物质也会对植物自身产生毒害作用[2]。另外,涝害胁迫致使植物体内活性氧产生和清除的平衡遭到破坏,从而加速H2O2、超氧阴离子等活性氧的累积,影响植物的正常生长[7]。涝害胁迫还会严重影响植物的光合作用,使叶片气孔关闭且气孔扩散阻力增加、气孔导度下降,进一步导致CO2与O2扩散交换受阻,光合作用下降;此外,涝害胁迫会破坏叶绿体结构,抑制光合色素合成,加速光合色素降解,降低光合产物的运输速率,从而影响植物能量的供应。
鉴于此,本研究通过对甜瓜耐涝型材料L45 和涝敏感型材料L39 叶片进行转录组和蛋白组测序,分析差异表达的基因和蛋白,以期挖掘甜瓜涝害胁迫响应中的关键基因及通路,从而为耐涝甜瓜分子育种实践提供理论依据。
1 材料与方法
1.1 试验材料与处理
2021年9—10月在江西省农业科学院园艺研究所的实验室和玻璃温室进行试验。供试材料为甜瓜(CucumismeloL.)品系L39 和L45,由江西省农业科学院园艺研究所西甜瓜研究室提供,其中L39 为涝敏感型材料,L45 为耐涝型材料[8]。选取籽粒饱满的甜瓜种子放入55 ℃温水中,迅速搅拌,水温降至30 ℃左右时停止搅拌,浸泡4 h。洗净种子表面黏液,沥干水分,用湿毛巾包裹后,放入30 ℃恒温设备中催芽,待80%以上种子露白即可播种。将萌发整齐一致的种子播于填装有混合基质(草炭∶蛭石∶珍珠岩=3∶1∶1,V/V)的塑料钵(7 cm×7 cm×7 cm)中,置玻璃温室培养,常规栽培管理。选取三叶一心时期长势一致的健壮植株,移入塑料盒(71 cm×45.5 cm×18 cm)中,进行涝害胁迫处理,水位保持在高出基质表面3 cm;对照不进行涝害胁迫处理,即正常管理。涝害胁迫后5 d分别取两个材料的处理组和对照组叶片样品,2 次生物学重复,每重复30株,样品收集后迅速置于液氮中,存储于-80 ℃冰箱备用。正常管理的对照样品分别命名为L39-C1、L39-C2、L45-C1和L45-C2,涝害胁迫处理的样品分别命名为L39-W1、L39-W2、L45-W1和L45-W2。
1.2 RNA提取与转录组测序
利用RNAprep pure Plant Kit植物总RNA提取试剂盒(天根,北京)提取RNA,DNase I (Sigma,美国)去除基因组DNA。利用Nanodrop 2000 超微量分光光度计(Thermo Fisher,美国)和Bioanalyzer 2100 生物分析仪(Agilent,德国)测定RNA的浓度和质量,质检合格的样品用于转录组文库构建。利用带有Oligo (dT)的磁珠富集mRNA,然后加入打断试剂使mRNA片段化,以其为模板反转录为第一链cDNA,再加入dNTPs、DNA聚合酶、RNase H 与缓冲液以合成第二链cDNA;经纯化回收、粘性末端修复、3′末端加碱基A、连接接头、片段大小的选择后,使用NEBNEXT Ultra RNA Library Prep Kit文库制备试剂盒(New England Biolabs,美国)进行PCR扩增;利用Agilent Bioanalyzer 2100生物分析仪检测cDNA 文库的质量,质检合格的样品利用Illumina HiSeqTM4000高通量测序平台(Illumina,美国)进行测序。
1.3 转录组数据分析
从下机原始数据中去除含接头的读段、含未知碱基比例超过5%的读段和低质量读段,将获得的有效读段通过HISAT2[9]映射到NCBI 甜瓜基因组(https://www.ncbi.nlm.nih.gov/genome/?term=Cucumis+melo)。利用StringTie 软件[10]通过FPKM(reads per kilo bases per million mapped reads)衡量基因的表达量。使用DESeq2[11]鉴定差异表达基因(differentially expressed genes,DEGs),筛选条件设置为:错误发生率(false discovery rate,FDR)≤0.05 且表达倍数差异在2 倍以上即|log2Ratio|≥1。使用Bioconductor 中的topGO[12]和KOBAS软件[13],以FDR≤0.05为标准,对DEG进行Gene Ontology(GO)富集分析和Kyoto Encyclopedia of Genes and Genomes(KEGG)途径富集分析。
1.4 蛋白质提取与蛋白组测序
利用三氯乙酸(trichloroacetic acid,TCA)/丙酮沉淀结合SDT 裂解法进行蛋白提取,采用二辛可酸(bicinchoninic acid,BCA)法进行蛋白质定量,然后各样品取蛋白质20 μg 分别加入6×上样缓冲液,沸水浴5 min,进行12% 十二烷基硫酸钠-聚丙烯酰胺凝胶电泳(sodium dodecyl sulfate-polyacrylamide gel electrophoresis,SDS-PAGE)电泳,考马斯亮蓝染色。各样品取80 μg蛋白质溶液,分别加入二硫苏糖醇(dithiothreitol,DTT)至终浓度为100 mmol·L-1,沸水浴5 min,冷却至室温。经尿素缓冲液、碘乙酰胺缓冲液、NH4HCO3溶液、胰蛋白酶缓冲液后收集滤液,采用C18 Cartridge 对肽段进行脱盐,肽段冻干后加入甲酸溶液复溶,OD280肽段定量。各样品采用纳升流速Easy nLC 系统(Thermo Fisher,美国)进行分离,缓冲液A 液为0.1%甲酸水溶液,B液为0.1%甲酸乙腈水溶液(乙腈为80%),样品经色谱分离后用 Q Exactive Plus 质谱仪(Thermo Fisher,美国)进行质谱分析,原始数据利用数据依赖性采集(data-dependent acquisition,DDA)方式进行采集。
1.5 蛋白组数据分析
利用MaxQuant 1.5.5.1 软件进行数据库搜索,使用的数据库为Uniprot-Cucumis_25421_20190812(http://www.uniprot.org),通过肽段水平过滤(FDR≤0.01)获得具有显著性的谱图和肽段列表,为了控制蛋白的假阳性率,在蛋白水平进行过滤(FDR≤0.01),采用无标记定量(label free quantitation,LFQ)算法进行定量分析[14],FDR≤0.05 且表达倍数差异在2 倍以上即|log2Ratio|≥1的为差异表达蛋白(differentially expressed proteins,DEPs)。以FDR≤0.05为标准,对DEPs进行GO富集分析和KEGG途径富集分析。
1.6 生理指标及测定方法
脱落酸、游离水杨酸、茉莉酸和α-亚麻酸含量采用高效液相色谱法测定。脱落酸的提取与测定参考Yang 等[15]的方法。游离水杨酸含量的测定参考Dong等[16]的方法。茉莉酸含量的测定参考Kristl 等[17]的方法。α-亚麻酸含量的测定参考杨会芳等[18]的方法。利用植物类黄酮含量检测试剂盒(索莱宝,北京)提取甜瓜叶片中的类黄酮,测定样本提取液在470 nm 处的吸光值,通过制作芦丁标准曲线计算样本中类黄酮的含量(单位为mg·g-1FW)。
1.7 实时荧光定量PCR(quantitative real-time PCR,qRT-PCR)验证
使用与转录组文库构建相同的RNA 样品,第一链cDNA 用TransScript®All-in-One First-Strand cDNA Synthesis SuperMix(全式金,北京)合成,每个样品3次重复,甜瓜CmACT基因作为内参基因[8]。PCR体系为20 μL:Top Green qPCR SuperMix(全式金,北京)10 μL、cDNA模板0.5 μL、10 μmol·L-1引物1 μL(表1)、ddH2O 8.5 μL。所用仪器为LightCycler®96(Roche,德国),程序设置为:95 ℃预变性 300 s;95 ℃变性 10 s,58 ℃退火 10 s,72 ℃延伸10 s,共45个循环;95 ℃熔解 10 s,65 ℃ 60 s,97 ℃ 1 s[8]。

表1 qRT-PCR所用引物Table 1 The primer sequences for qRT-PCR
1.8 数据分析
利用SAS 9.1.3 软件的ANOVA 程序进行方差分析,采用最小显著差异法进行差异显著性比较分析,显著性水平为P<0.01。
2 结果与分析
2.1 涝害胁迫对内源性物质含量的影响
如图1 所示,涝害胁迫处理后,涝敏感型材料L39中脱落酸含量显著提高,为对照的2.72 倍,而耐涝型材料L45 中脱落酸含量无显著变化;L39 和L45中游离水杨酸含量在涝害胁迫处理后都显著提高,分别为对照的1.63和1.72倍;茉莉酸含量在涝害胁迫处理后也显著提高,L39和L45中分别为对照的1.90和2.12倍。涝害胁迫后类黄酮显著积累,L39和L45中类黄酮含量分别为对照的2.27 和2.47 倍;涝害胁迫后α-亚麻酸含量在耐涝型材料L45中显著提高,为对照的2.24倍,而在L39中无显著变化(图2)。
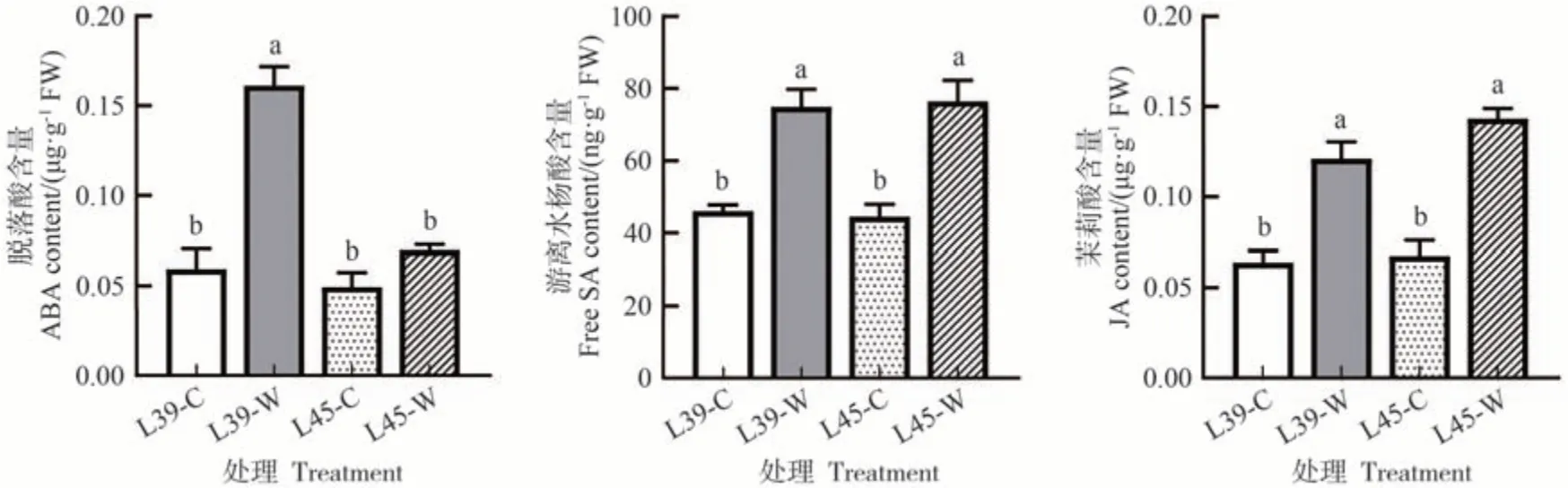
图1 涝害胁迫处理对甜瓜叶片激素含量的影响Fig.1 Effects of waterlogging stress on endogenous hormone contents of melon leaves
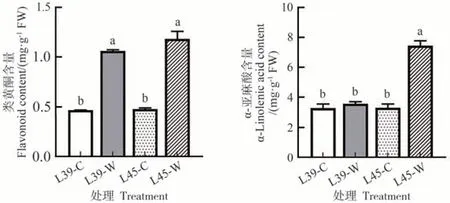
图2 涝害胁迫处理对甜瓜叶片类黄酮和α-亚麻酸含量的影响Fig.2 Effects of waterlogging stress on flavonoid and α-linolenic acid contents of melon leaves
2.2 转录组测序整体评估
利用高通量测序仪对8个样品测序得到44 275 178~48 310 008 条原始读段,再通过数据过滤处理得到38 287 494~43 584 000条有效读段,有效碱基数为5.74~6.54 Gb,GC含量为43.50%~47.00%;与甜瓜参考基因组进行比对,发现成功比对到25 254 777~33 659 962条有效读段;碱基质量值大于20 的比例为99.98%~99.99%,碱基质量值大于30的比例为97.90%~98.08%,表明测序质量较高(表2)。
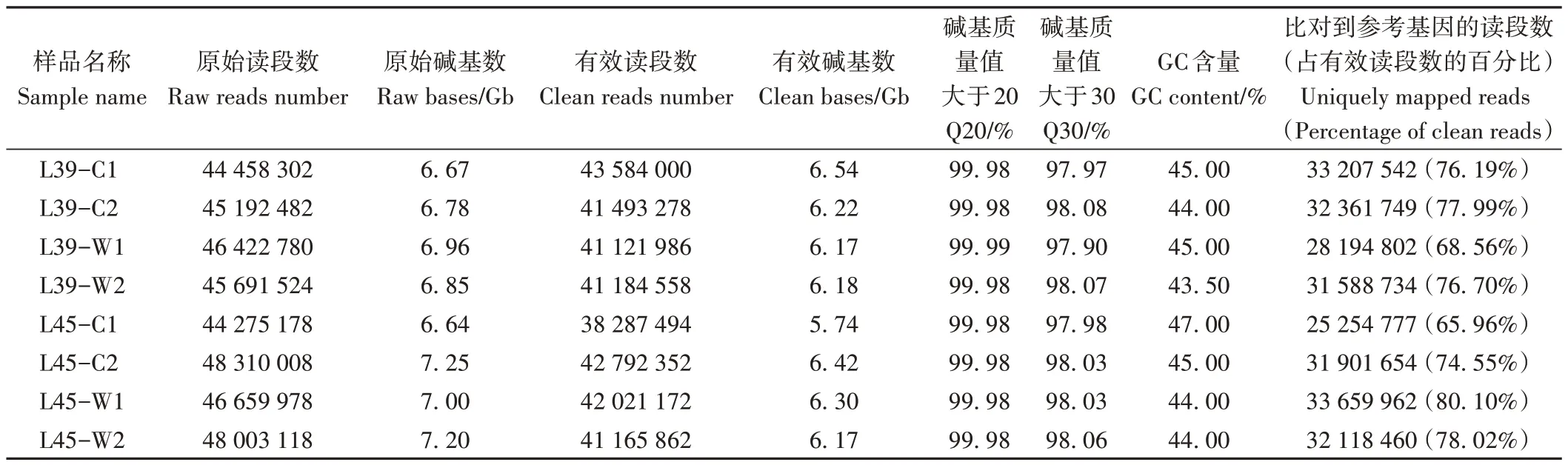
表2 转录组测序数据统计Table 2 Summary of RNA-seq data
2.3 差异表达基因鉴定
在L39-W vs L39-C 和L45-W vs L45-C 两个比较组中分别鉴定差异表达基因(differentially expressed genes,DEGs),发现L39-W vs L39-C的差异表达基因数为3 532个,上调和下调基因数分别为2 266和1 266个;L45-W vs L45-C 的差异表达基因数为1 842 个,上调和下调基因数分别为1 280和562个(图3)。
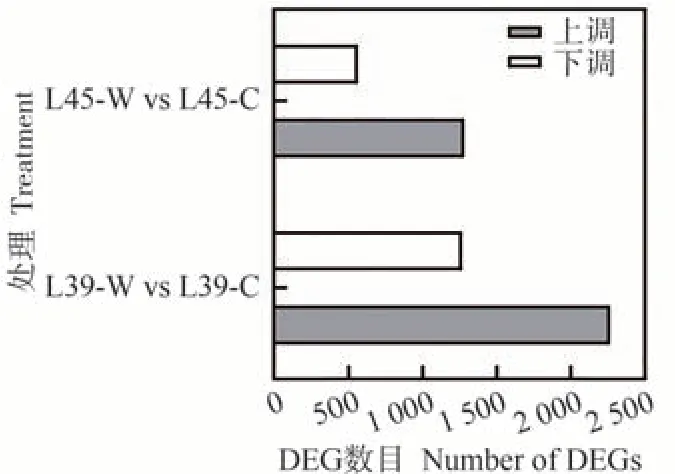
图3 差异表达基因统计Fig.3 Statistical histogram of differentially expressed genes (DEGs)
2.4 差异表达基因功能显著性富集分析
差异表达基因GO功能显著性富集分析发现,L39-W vs L39-C 和L45-W vs L45-C 分别有280 和177 个显著富集的GO 条目,其中生物过程条目分别有139 和104 个。生物过程条目中,光合作用(GO:0015979)、肉桂酸生物合成过程(GO:0009800)、次级代谢产物生物合成过程(GO:0044550)、氧化还原过程(GO:0055114)、脱落酸激活信号通路(GO:0009738)、L-苯丙氨酸分解代谢过程(GO:0006559)、缺水反应(GO:0009414)、吲哚硫甙代谢过程(GO:0042343)、苯丙素生物合成过程(GO:0009699)、脱落酸响应(GO:0009737)等在L39-W vs L39-C 中显著富集;乙烯激活信号通路(GO:0009873)、谷胱甘肽代谢过程(GO:0006749)、毒素分解代谢过程(GO:0009407)、茉莉酸介导的信号通路(GO:0009864)、乙烯响应(GO:0009723)、吲哚硫甙代谢过程(GO:0042343)、水杨酸响应(GO:0009751)、苯丙素生物合成过程(GO:0009699)、叶片衰老负调控(GO:1900056)、水杨酸介导的信号通路(GO:0009862)等在L45-W vs L45-C 中显著富集。
差异表达基因KEGG 功能显著性富集分析发现,L39-W vs L39-C和L45-W vs L45-C分别有35和11个显著富集的KEGG途径,乙醛酸和二羧酸代谢(ko00630)、缬氨酸、亮氨酸和异亮氨酸降解(ko00280)、甘氨酸、丝氨酸和苏氨酸代谢(ko00260)、苯丙素生物合成(ko00940)、酮类的合成与降解(ko00072)、半萜类和三萜类生物合成(ko00909)、脂肪酸降解(ko00071)、丙氨酸、天冬氨酸和谷氨酸代谢(ko00250)等代谢途径在L39-W vs L39-C 中显著富集(图4);植物-病原菌互作(ko04626)、黄酮和黄酮醇生物合成(ko00944)、α-亚麻酸代谢(ko00592)、亚油酸代谢(ko00591)等代谢途径在L45-W vs L45-C中显著富集(图5);此外,光合作用(ko00195)、类胡萝卜素生物合成(ko00906)、植物激素信号转导(ko04075)、丝裂原活化蛋白激酶(mitogen-activated protein kinase,MAPK)信号通路(ko04016)、异黄酮生物合成(ko00943)在L39-W vs L39-C和L45-W vs L45-C中都显著富集。
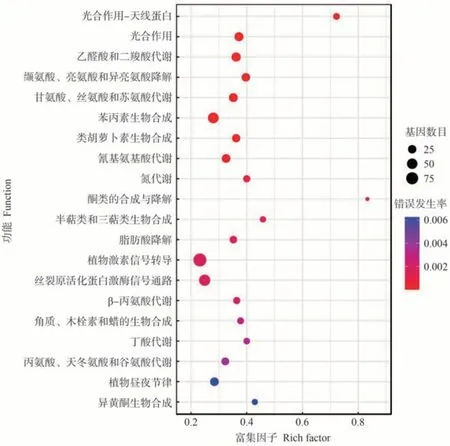
图4 L39-W vs L39-C差异表达基因的KEGG富集分析Fig.4 Significant enrichment analysis of KEGG pathway for DEGs of L39-W vs L39-C
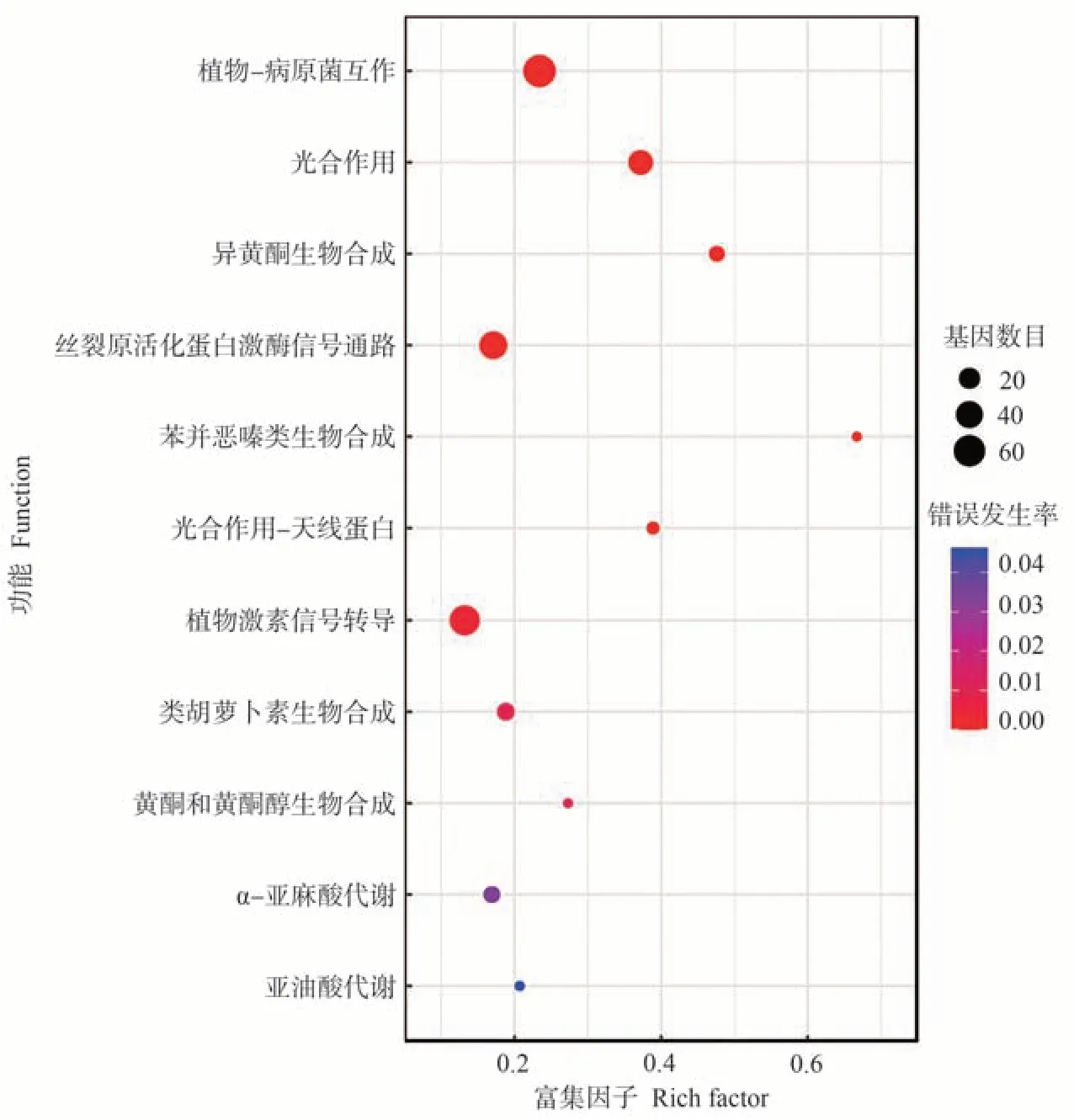
图5 L45-W vs L45-C差异表达基因的KEGG富集分析Fig.5 Significant enrichment analysis of KEGG pathway for DEGs of L45-W vs L45-C
2.5 蛋白组测序整体评估
本研究共鉴定得到22 765条肽段,3 259个蛋白。对鉴定到的肽段序列长度进行统计分析,发现多数肽段分布在5~20 aa,占所有肽段的86.94%(图6-A)。对所有鉴定到的蛋白质进行统计,发现蛋白相对分子质量主要为10~60 kDa,占总蛋白的75.97%。分子质量为0~10 kDa 和60~100 kDa的蛋白分别占总蛋白的1.38%和16.48%,大于100 kDa的蛋白约占总蛋白6.17%(图6-B)。
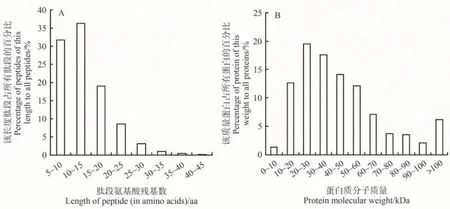
图6 肽段与蛋白质统计Fig.6 Statistical histogram of peptide and protein
2.6 差异表达蛋白鉴定
在L39-W vs L39-C 和L45-W vs L45-C 两个比较组中分别鉴定差异表达蛋白(differentially expressed proteins,DEPs),发现L39-W vs L39-C 的差异表达蛋白数为105 个,上调和下调蛋白数分别为73 和32 个;L45-W vs L45-C的差异表达蛋白数为38个,上调和下调蛋白数分别为21和17个(图7)。
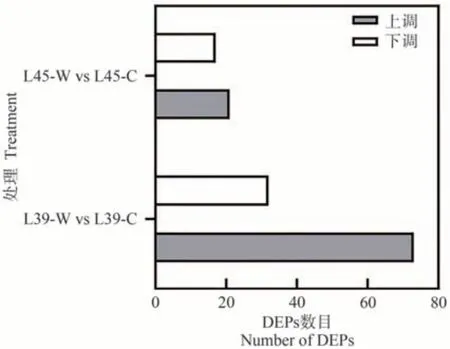
图7 差异表达蛋白统计Fig.7 Statistical histogram of differentially expressed proteins (DEPs)
2.7 差异表达蛋白功能显著性富集分析
差异表达蛋白GO功能显著性富集分析发现,L39-W vs L39-C 和L45-W vs L45-C 分别有82和53个显著富集的GO条目,其中生物过程条目分别有39和25个。生物过程条目中,氧化还原过程(GO:0055114)、过氧化氢分解代谢过程(GO:0042744)、氧化应激响应(GO:0006979)、甘氨酸代谢过程(GO:0006544)、L-丝氨酸代谢过程(GO:0006563)、苏氨酸代谢过程(GO:0006566)、类固醇生物合成过程(GO:0006694)、渗透胁迫响应(GO:0006970)、香豆素生物合成过程(GO:0009805)、脂肪酸β 氧化(GO:0006635)等在L39-W vs L39-C 中显著富集;腈代谢过程(GO:0050898)、苯丙素生物合成过程(GO:0009699)、花青素3-O-葡萄糖苷生物合成过程(GO:0033485)、飞燕草色素3-O-葡萄糖苷生物合成过程(GO:0033486)、色素沉积(GO:0043473)、蛋白谷胱甘肽化(GO:0010731)、韧皮部蔗糖装载(GO:0009915)、D-丝氨酸代谢过程(GO:0070178)、类黄酮生物合成过程(GO:0009813)、胁迫应答(GO:0006950)等在L45-W vs L45-C中显著富集。
差异表达蛋白KEGG 功能显著性富集分析发现,L39-W vs L39-C和L45-W vs L45-C分别有16和13个显著富集的KEGG途径。苯丙素类生物合成(ko00940)、托烷、哌啶和吡啶生物碱生物合成(ko00960)、内质网中蛋白质加工(ko04141)、脂肪酸降解(ko00071)、卟啉与叶绿素代谢(ko00860)、酪氨酸代谢(ko00350)等在L39-W vs L39-C 中显著富集(图8);α-亚麻酸代谢(ko00592)、黄曲霉毒素生物合成(ko00254)、氰基氨基酸代谢(ko00460)、细胞色素P450 对有害异物的代谢(ko00980)、铂类耐药性(ko01524)等在L45-W vs L45-C中显著富集(图9);此外,次生代谢物生物合成(ko01110)和类黄酮生物合成(ko00941)在L39-W vs L39-C 和L45-W vs L45-C中都显著富集。
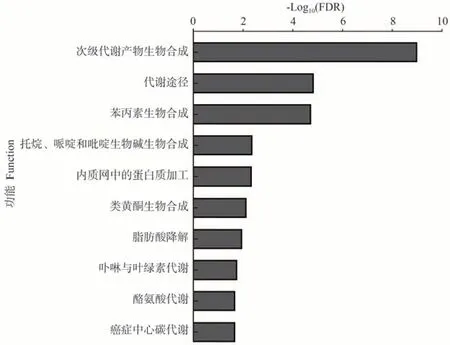
图8 L39-W vs L39-C差异表达蛋白的KEGG富集分析Fig.8 Significant enrichment analysis of KEGG pathway for DEPs of L39-W vs L39-C
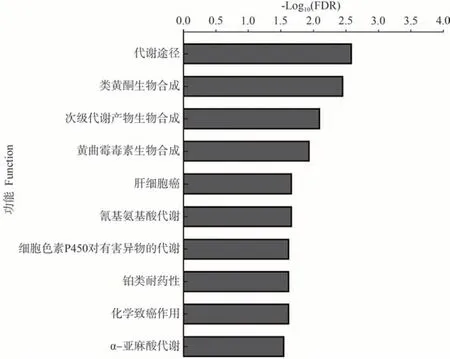
图9 L45-W vs L45-C差异表达蛋白的KEGG富集分析Fig.9 Significant enrichment analysis of KEGG pathway for DEPs of L45-W vs L45-C
2.8 转录组与蛋白组联合分析
将转录组与蛋白组数据联合分析发现,L39-W vs L39-C比较组中有38个基因在mRNA和蛋白质水平上联合上调表达,12 个基因联合下调表达(电子附表1);L45-W vs L45-C 比较组中有7个基因在mRNA 和蛋白质水平上联合上调表达,3个基因联合下调表达(电子附表2)。L39-W vs L39-C 比较组中联合上调表达的基因显著富集在脂肪酸降解(ko00071)、类黄酮生物合成(ko00941)、苯丙烷生物合成(ko00940)、缬氨酸、亮氨酸和异亮氨酸生物合成(ko00290)、二苯乙烯类、二芳庚烷类和姜酚生物合成(ko00945)、内质网中的蛋白质加工(ko04141)、苯丙氨酸代谢(ko00360)、精氨酸和脯氨酸代谢(ko00330)KEGG 途径中,而L45-W vs L45-C比较组中联合上调表达的基因显著富集在α-亚麻酸代谢(ko00592)、苯丙烷生物合成(ko00940)、二苯乙烯类、二芳庚烷类和姜酚生物合成(ko00945)、亚油酸代谢(ko00591)和类黄酮生物合成(ko00941)KEGG 途径中;对联合下调表达的基因进行分析发现,卟啉与叶绿素代谢(ko00860)和叶酸合成(ko00790)在L39-W vs L39-C 中显著富集,MAPK 信号通路(ko04016)在L45-W vs L45-C中显著富集(表3)。
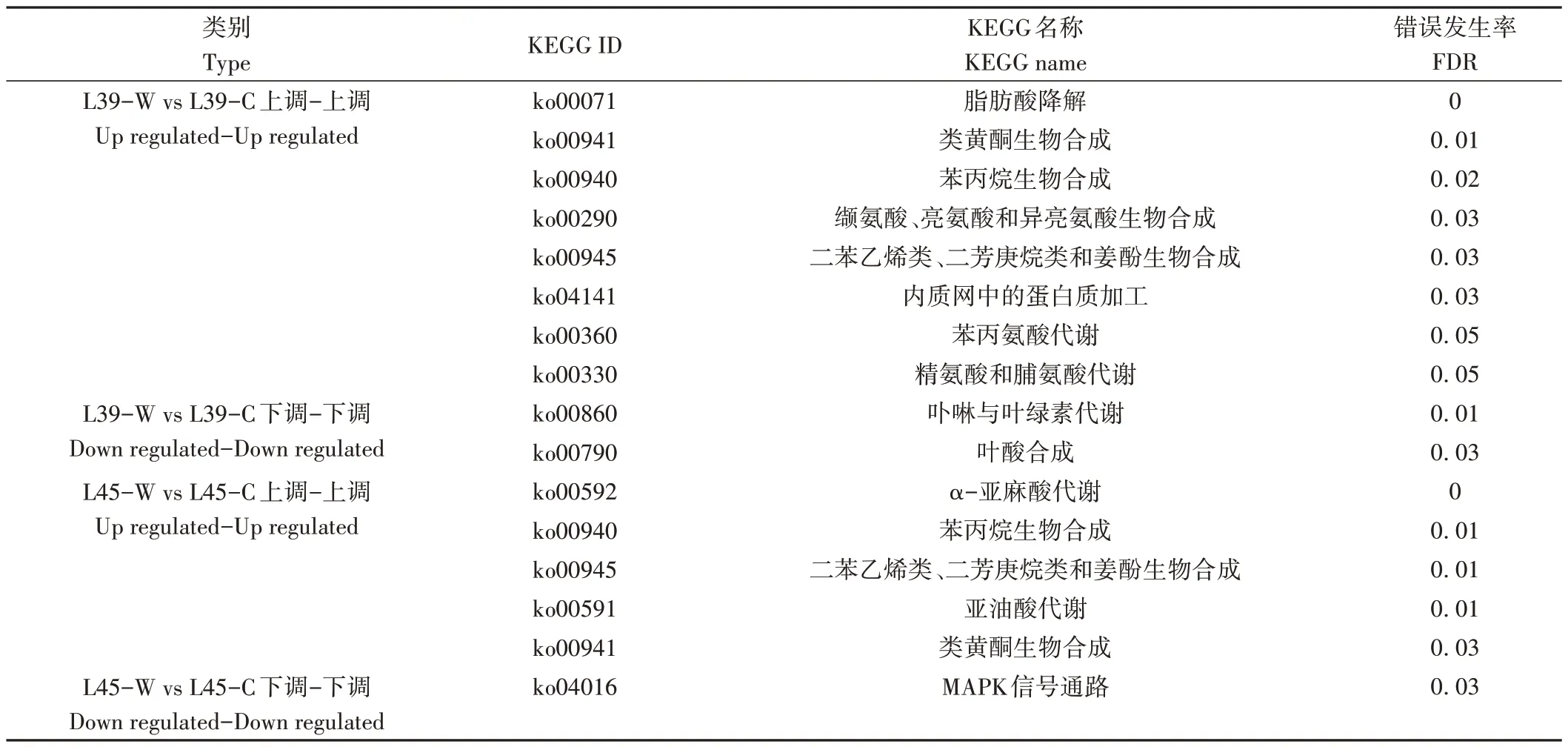
表3 转录组与蛋白组联合表达基因KEGG富集分析Table 3 Significant enrichment analysis of KEGG pathway for jointly expressed genes between transcriptome and proteome
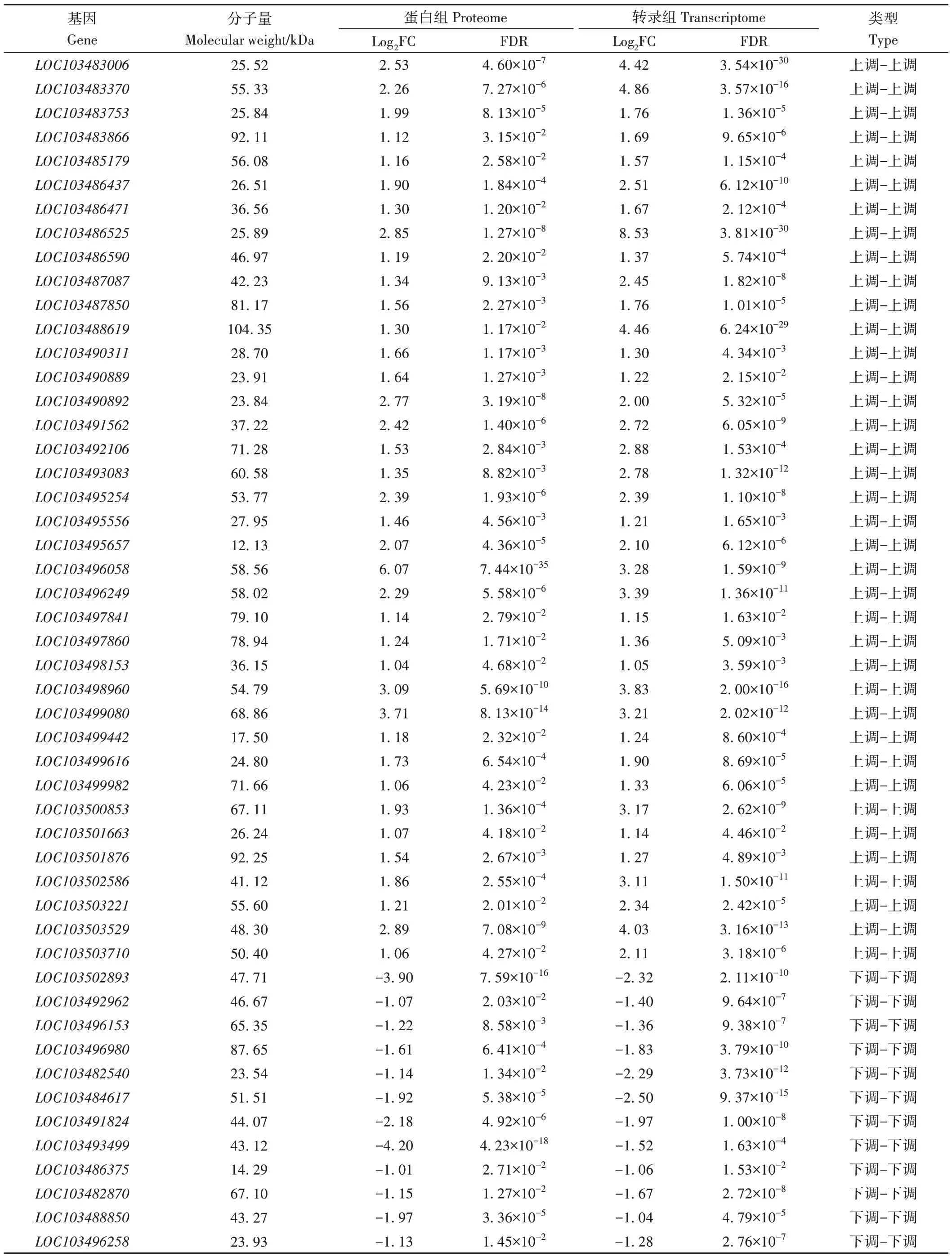
电子附表1 L39-W vs L39-C联合表达基因Electronic Table S1 Jointly expressed genes between transcriptome and proteome of L39-W vs L39-C
2.9 差异表达基因的qRT-PCR验证
利用qRT-PCR 方法检测了参与类黄酮生物合成、苯丙烷生物合成、α-亚麻酸代谢、脱落酸响应、水杨酸生物合成等15 个差异表达基因(表1)的相对表达水平,以验证转录组测序结果的可靠性。如图10 所示,qRT-PCR 与转录组测序的结果基本一致,相关性分析结果显示二者的相关系数达0.983 2,表明本研究中转录组测序结果的准确性和可重复性较高。
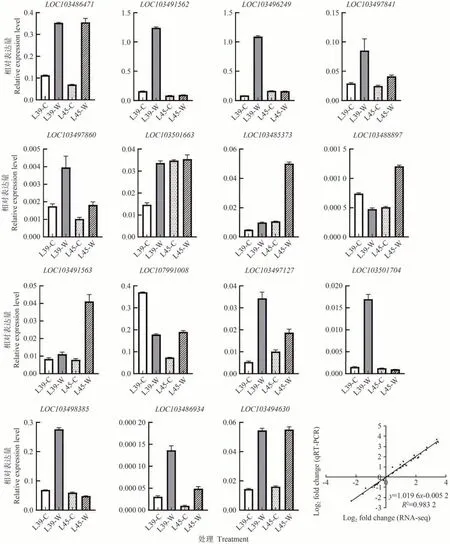
图10 差异表达基因的qRT-PCR验证Fig.10 qRT-PCR confirmation of differentially expressed genes
3 讨论
涝害胁迫是世界范围内农业遭遇最为频繁的非生物胁迫之一,严重影响作物的产量和品质。为了探明甜瓜响应涝害胁迫的分子机制,本研究利用转录组和蛋白组测序对甜瓜耐涝型材料L45 和涝敏感型材料L39在正常培养和涝害胁迫处理条件下进行了比较分析,发现L39-W vs L39-C的差异表达基因数为3 532个(图3),差异表达蛋白数为105 个(图7),而L45-W vs L45-C的差异表达基因数为1 842 个(图3),差异表达蛋白数为38 个(图7)。与耐涝型材料L45 相比,涝敏感型材料L39在涝害胁迫处理后的差异表达基因和差异表达蛋白数目更多,这与黄瓜[19-20]、野豇豆[21]、紫花苜蓿[22]等植物的研究结果相类似,表明涝敏感型材料遭遇涝害胁迫后转录和蛋白水平的变化更大。
植物激素作为信号转导途径的关键因子,在植物生长发育以及对抗各种生物和非生物胁迫中都发挥着重要作用。据报道,脱落酸、水杨酸、茉莉酸、乙烯等激素参与了植物涝害胁迫响应[3,23]。本研究中转录组结果发现植物激素信号转导KEGG 途径在L39-W vs L39-C 和L45-W vs L45-C 中都显著富集(图4、5)。脱落酸不仅可以调节气孔关闭,还可以影响与应激反应和代谢变化有关的基因表达,在植物逆境胁迫响应中发挥重要作用[24]。Wang 等[25]发现在芝麻中,10 个脱落酸途径基因对涝害胁迫有响应。涝害胁迫处理后葡萄叶片中脱落酸生物合成相关基因上调表达[26]。本研究中,差异表达基因GO 功能显著性富集分析结果发现脱落酸激活信号通路、脱落酸响应和缺水反应在L39-W vs L39-C 中显著富集;此外,涝害胁迫处理后,涝敏感型材料L39 中脱落酸含量显著提高,而耐涝型材料L45中无显著变化(图1),推测脱落酸的涝害胁迫响应可能在涝敏感型甜瓜材料中更为突出。水杨酸在提高植物抗逆性方面具有巨大潜力,它可以减轻干旱、低温、高温、盐胁迫等非生物胁迫的不利影响[16,27-29]。Koramutla 等[30]报道水杨酸在涝害胁迫下可以促进小麦形成不定根以增强涝害适应能力。葡萄中参与水杨酸生物合成相关基因在涝害胁迫后诱导表达[26]。本研究发现涝害胁迫处理后,涝敏感型材料L39 和耐涝型材料L45中水杨酸含量都显著提高(图1),推测水杨酸在甜瓜响应涝害胁迫中发挥重要作用。茉莉酸在介导或协调植物的应激反应中具有关键功能[31-32]。茉莉酸途径相关基因已被证实参与涝害胁迫响应[25-26]。相类似地,本研究中转录组与蛋白组联合分析发现,L39中参与茉莉酸生物合成的基因LOC103497841和LOC103497860在涝害胁迫后显著上调表达,L45中参与茉莉酸生物合成的基因LOC103485373和LOC107991008在涝害胁迫后显著上调表达(电子附表1、2),此外,涝害胁迫处理后,涝敏感型材料L39和耐涝型材料L45中茉莉酸含量都显著提高(图1),表明茉莉酸在甜瓜涝害胁迫响应中发挥重要作用。以上响应涝害胁迫的激素可能有助于提高甜瓜的耐涝性,需要进一步深入研究。
来源于苯丙烷途径的类黄酮物质是一类重要的防御性抗氧化剂,具有自由基清除活性和抗氧化活性[33]。Wang等[34]发现涝害胁迫可以显著提高菊花查尔酮异构酶(chalcone isomerase,CHI)、黄烷酮3-羟化酶(flavanone 3-hydroxylase,F3H)、黄烷酮4-还原酶(flavanone 4-reductase,DFR)和花色素合成酶(anthocyanidin synthase,ANS)编码基因的转录水平,类黄酮含量显著提高。Li等[35]报道涝害胁迫使类黄酮生物合成相关基因在枫杨叶片中显著上调。本研究发现涝害胁迫处理后,涝敏感型材料L39 和耐涝型材料L45 中类黄酮含量都显著提高(图2),此外,转录组与蛋白组联合分析中,类黄酮生物合成途径在L39-W vs L39-C 和L45-W vs L45-C 比较组的联合上调表达的基因中都显著富集(表3),暗示类黄酮在响应涝害胁迫中的重要作用,可能作为抗氧化剂来减轻涝害胁迫造成的ROS 损伤。Zeng 等[22]在紫花苜蓿中发现涝害胁迫处理使苯丙烷生物合成途径在耐涝品种和涝敏感品种中都显著富集。本研究也有类似结果,转录组与蛋白组联合分析中发现苯丙烷生物合成途径在耐涝型材料L45和涝敏感型材料L39比较组的联合上调表达的基因中都显著富集(表3),推测苯丙烷途径在植物响应涝害胁迫中可能发挥重要作用。从膜脂质中释放α-亚麻酸对植物响应非生物胁迫非常重要[36]。涝害胁迫后,枫杨叶片中α-亚麻酸代谢途径显著上调[35]。Qiao 等[37]发现鸭茅遭遇涝害胁迫后叶片中参与α-亚麻酸代谢途径的基因表达上调。本研究发现涝害胁迫处理后,耐涝型材料L45 中α-亚麻酸含量显著提高,而涝敏感型材料L39 无显著变化(图2),此外,转录组与蛋白组联合分析中,α-亚麻酸代谢途径仅在L45-W vs L45-C比较组的联合上调表达的基因中显著富集(表3),推测甜瓜可能通过调控α-亚麻酸代谢相关基因的表达来增强膜脂质的不饱和性,从而提高对涝害胁迫的耐受性。下一步将结合基因功能研究方法对相关基因进行功能验证,以期深入解析甜瓜响应涝害胁迫的分子机制。
4 结论
本研究利用转录组和蛋白组学方法探讨了甜瓜耐涝型材料L45 和涝敏感型材料L39 在正常培养和涝害胁迫处理条件下的差异表达基因和蛋白,发现L45-W vs L45-C 中鉴定到1 842 个差异表达基因和38 个差异表达蛋白,而L39-W vs L39-C 中鉴定到3 532 个差异表达基因和105 个差异表达蛋白。差异表达基因富集分析与内源激素含量表明脱落酸、水杨酸和茉莉酸参与甜瓜涝害胁迫响应。转录组和蛋白组联合分析发现,类黄酮和苯丙烷生物合成途径在甜瓜响应涝害胁迫中发挥重要作用。此外,α-亚麻酸代谢相关基因的表达可能有助于提高甜瓜耐涝性。
猜你喜欢
今日农业(2021年15期)2021-10-14
保健与生活(2021年13期)2021-07-14
安徽农业科学(2021年4期)2021-03-18
中国果树(2020年2期)2020-07-25
天然产物研究与开发(2018年11期)2018-11-30
湖北农业科学(2018年8期)2018-06-08
现代农业科技(2018年4期)2018-03-23
临床医药文献杂志(电子版)(2017年77期)2017-08-15
食品界(2016年4期)2016-02-27
军事文摘·科学少年(2015年6期)2015-05-30
