
FMD73D基因真核表达质粒的构建、表达及对I型IFN信号通路的作用
2024-04-27 22:21苗勤吴香菊齐静丛晓燕李均同王林杜以军
山东农业科学 2024年2期
苗勤 吴香菊 齐静 丛晓燕 李均同 王林 杜以军


关键词:口蹄疫病毒:3D蛋白;真核表达:I型IFN信号通路
口蹄疫(foot-and -mouthdisease,FMD)是由口蹄疫病毒(Foot-and-mouth disease VLrLLS,FM-DV)引起的一种急性、高度接触性人畜共患传染病,主要感染牛、羊、猪等偶蹄动物,给畜牧业造成了严重的经济损失,被世界卫生组织列为A类动物疫病之首,我国也将其列为一类传染病。口蹄疫病毒属小RNA病毒科,是单股正链RNA病毒,呈二十面体对称结构。FMDV基因组全长约8500bp,仅有一个开放阅读框(open readingframe,ORF),约6.5 kb,编码一个大的多聚蛋白,在L、2A和3C的作用下裂解产生了病毒的4种结构蛋白[VP4(1A)、VP2(1B)、VP3(1C)和VPl(1D)]和8个非结构蛋白(L、2A、2B、2C、3A、3B、3C、3D)。FMDV有7种血清型,分别为A型、O型、C型、亚洲I型、SAT1型、SAT2型和SAT3型,且各血清型之间无交叉保护。
I型IFN是机体抵抗病毒天然免疫的核心,FMDV感染细胞后,病毒复制产生的RNA被宿主细胞的模式识别受体所识别,激活下游信号通路,刺激产生I型IFN和促炎细胞因子,从而启动抗病毒反应。已有研究表明,FMDV多种蛋白,如前导蛋白L、蛋白水解酶3C、非结构蛋白2B、2C等,参与调节先天性免疫信号通路,拮抗宿主天然免疫,促进病毒增殖。
FMDV 3D蛋白是病毒的RNA聚合酶,介导FMDV基因组的合成,且核苷酸和氨基酸序列高度保守。此外,3D蛋白具有T细胞表位,是一种潜在的免疫增强剂,还可以作为免疫佐剂使用。但3D蛋白在天然免疫中的作用尚不明确,本研究旨在构建FMDV 3D基因的真核表达质粒,并探索3D蛋白对I型IFN信号通路的影响,为FMD的防控提供新的靶向分子。
1材料与方法
1.1试验材料
1.1.1细胞与质粒HEK-293T细胞、PK-15细胞、pXJ41真核表达载体、荧光素酶报告基因质粒pIFN-Luc和内参报告质粒pRL-TK均由山东省畜禽疫病防治与繁育重点实验室保存。
1.1.2主要试剂PrimeSTAR⑩HS DNA Polymer-ase、DL5000 DNA Marker、DL2000 DNA Marker,pMD18-T载体购自TaKaRa宝生物工程(大连)有限公司;Lipofectamine3000 Reagent、Alexa Flu-or 488-羊抗鼠IgG(H+L)购自Invitrogen公司;Western一抗稀释液购自碧云天生物技术有限公司;NC膜購自BIO-RAD公司;辣根过氧化物(HRP)标记的羊抗兔抗体购自武汉博士德生物工程有限公司;Sparkjade ECL super(极超敏化学发光试剂盒)购自山东思科捷生物技术有限公司;感受态细胞DH5a、双荧光素酶报告基因试剂盒、RNA-easy Isolation Reagent试剂盒均购自南京诺唯赞生物科技有限公司:限制性内切酶HindⅢ、Kpn I及T4 DNA Ligase购自Thermo FisherScientific公司;质粒小提试剂盒、通用型DNA纯化回收试剂盒购自北京天根生物工程有限公司:DMEM细胞培养基、Opti-MEM培养基、胰蛋白酶一EDTA均购自Gibco公司:抗荧光衰减封闭剂购自Solarbio公司;鼠源、兔源Myc抗体购自Sigma -Aldrich公司;actin抗体购自上海泊湾生物科技有限公司。
1.2试验设计与方法
1.2.1FMDV 3D基因真核表达质粒的构建与鉴定
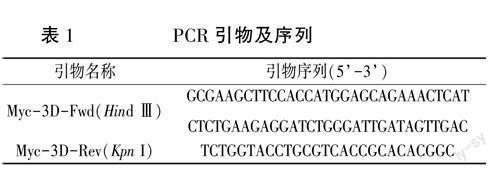
根据GenBank公布的FMDV O/BY/CHA/2010(GenBank accession no. JN998085) 3D基因序列,由北京擎科生物技术有限公司合成3D基因并连接到pMD18-T载体上。结合真核表达载体pXJ41上的酶切位点序列,加入保护性碱基及Myc标签序列,利用分子生物学软件Primer Premier5.0设计了一对扩增3D基因的特异性引物:Myc-3D-Fwd(HindⅢ)和Myc-3D-Rev,引物削匕京擎科生物技术有限公司合成,序列见表1。
以pMD18-T-3D克隆质粒为模板进行PCR扩增,PCR反应体系(总体系为25uL)如下:上、下游引物(10pmol/uL)各1uL,模板(200ng/uL)0.5uL,dNTP 2uL,5xPrime STAR buffer 5uL,PrimeSTARHS DNA Polyerase(2.5 U/卜LL) 0.25uL,ddH,0补足至25uL。反应条件如下:98℃预变性2min;98℃变性10s,58℃退火30s,72℃延伸2min,30个循环;72℃延伸7min;4℃保存。PCR产物进行琼脂糖凝胶电泳,用DNA纯化回收试剂盒进行回收。
将回收纯化的3D基因和pXJ41载体用Hindm/Kpn I双酶切,酶切产物胶回收纯化后用T4DNA Ligase进行16℃过夜连接,将连接产物转化至DH5a感受态细胞,37℃过夜培养16h,挑取单菌落接种于含有氨苄西林抗性的LB培养基中,37℃、200r/min过夜培养后提取质粒,进行HindⅢlKpn I双酶切鉴定,鉴定为阳性的质粒送北京擎科生物科技有限公司测序,命名为pXJ41 -Myc-3D。
1.2.2Western blotting检测FMDV 3D蛋白的表达
将HEK-293T细胞接种于6孔板中进行培养,待细胞密度达到70%~90%时,用Lipofectamine3000转染试剂按说明书进行操作,转染质粒pXJ41-Myc-3D和pXJ41空载体各2ug。培养24h后,弃去上清,用等体积预冷的PBS洗涤细胞2次,将6孑L板置于冰上,加入细胞裂解液裂解细胞20min后,4℃12000r/min离心10min。取上清液与上样缓冲液混合后煮沸10min。将处理好的样品进行12% SDS-PAGE凝胶电泳后,采用湿法(100V,70min)将蛋白转印至NC膜。将NC膜转移至5%脱脂奶粉封闭液中,室温轻摇封闭1h,用一抗稀释液稀释的兔抗Myc和兔抗actin于4℃过夜孵育,二抗为用5%脱脂奶粉稀释的辣根过氧化物(HRP)标记的羊抗兔抗体,室温孵育1h,洗膜后用Sparkjade ECL super(极超敏化学发光试剂盒)显色,BIO-RAD凝胶成像仪进行曝光。
1.2.3IFA检测FMDV 3D蛋白的细胞定位用无菌镊子夹取细胞爬片置于24孔板内,接种PK-15细胞,待细胞密度达到40%~50%时,用Lipo-fectamine3000转染试剂按说明书进行操作,各孔分别转染质粒pXJ41-Myc-3D和pXJ41空载体各0.5ug。培养24h后弃上清,用4%多聚甲醛固定45min;PBS洗10minx3次后,加入0.1%TritonX-100室温通透30min;PBS洗10min×3次后,加入1:600稀释的鼠源Myc抗体室温孵育2h;PBS洗10minx3次后,加入1:1000稀释的Alexa Fluor488-羊抗鼠抗体,室温避光孵育1h:PBS洗10minx3次后,加入1:10000稀释的DA-PI室温孵育5min;PBS洗10minx3次后,取出爬片置于滴有抗荧光衰减封闭剂的载玻片上,封片后于荧光显微镜下观察3D蛋白在细胞内的定位情况。
1.2.4Luciferase检测FMDV 3D蛋白对IFN启动子的影响
将HEK-293T细胞接种于12孔板中进行培养,待细胞密度达到70%~90%时,用Lipofectamine3000转染试剂按说明书进行操作,分别转染pXJ41-Myc-3D和空载体pXJ41各0.4ug,同时分别转染IFN启动子报告基因质粒和内参质粒pRL-TK 0.04ug。24 h后感染水疱性口炎病毒(Versicular sto-matLtLs VLrUS,VSV)(MOI=0.1),10h后,按双荧光素酶报告基因试剂盒说明书方法收取细胞进行检测。
1.2.5Real-time PCR检测FMDV 3D蛋白对IFN-mRNA的影响将HEK-293T细胞接种于24孔板中进行培养,待细胞密度达到70%~90%时,用Lipofectamine⑩3000转染试剂按说明书进行操作,分别转染质粒pXJ41-Myc-3D和空载体pXJ41各0.5ug,24h后感染VSV(MOI=0.1),10h后,收集细胞使用RNA-easy Isolation Reagent试剂盒提取总RNA,以其为模板反转录得到cD-NA,以GAPDH作为内参基因检测IFN mRNA水平,Real-time PCR引物序列详见表2。
1.2.6Real-time PCR检测病毒拷贝数将HEK-293T细胞接种于24孔板中进行培养,待细胞密度达到70%~90%时,用Lipofectamine3000转染试剂按说明书进行操作,分别转染质粒pXJ41-Myc-3D和空载体pXJ41各0.5ug,24h后感染VSV(MOI=0.1),10h后,细胞及其上清反复冻融后,一部分用于病毒滴度检测;一部分使用RNA-easyIsolation Reagent试剂盒提取总RNA,以其为模板反转录得到cDNA,绝对荧光定量检测VSV拷贝数。
1.2.7TCID50测定病毒滴度将HEK-293T细胞接种于96孔板中进行培养,待细胞密度达到90%时,将1.2.6中的样品12000r/min离心10min后取上清,用2%细胞维持液按10倍梯度稀释,共10个梯度,每个梯度4个重复,将稀释好的样品加入96孔板中,每孔100uL,并设置8个孔加入等体积细胞维持液做阴性对照,观察并记录细胞病变情况,按照Reed-Muench法计算TCID50。
1.3数据统计与分析
使用GraphPad Prism 5软件对试验数据进行统计学分析。
2结果与分析
2.1pXJ41-Myc-3D真核表达质粒的构建及鉴定
以pMD18-T-3D克隆质粒为模板,PCR扩增获得3D基因。扩增产物经1%琼脂糖凝胶电泳可见大小约1500bp的特异性目的片段,与预期大小相符(图1)。重组质粒pXJ41-Myc-3D经HindⅢ/Kpn I双酶切后得到预期大小的片段(图2),测序结果表明pXJ41-Myc-3D真核表达质粒构建成功。
2.2Western blotting检测3D蛋白的表达
经Western blotting检测到pXJ41-Myc-3D在HEK-293T细胞中成功表达,大小约55kDa左右(图3)。
2.3IFA检测FMDV 3D蛋白的细胞定位
IFA结果显示,FMDV 3D蛋白在PK-15细胞中的表達主要定位在细胞核内,细胞胞浆中也有少量表达(图4)。
2.4双荧光素酶报告基因检测FMDV 3D蛋白对IFN启动子的影响
双荧光素酶报告基因检测结果显示,VSV感染后,pXJ41-Myc-3D转染组的IFN启动子活性极显著低于空载体pXJ41转染组,表明FMDV3D蛋白抑制了VSV诱导的IFN启动子活性(图5)。
2.5Real-time PCR检测FMDV 3D蛋白对IFNmRNA的影响
Real-time PCR检测结果显示,VSV刺激后,pXJ41-Myc-3D转染组的IFN mRNA表达水平极显著低于空载体pXJ41转染组,表明FMDV 3D蛋白抑制了VSV诱导的IFN mRNA表达水平(图6)。
2.6FMDV 3D蛋白对VSV增殖的影响
Real-time PCR检测VSV拷贝数结果显示,pXJ41-Myc-3D转染组极显著高于空载体组(图7)。TCID50结果显示,pXJ41-Myc-3D转染组VSV TCID50明显高于空载体转染组(图8)。表明FMDV 3D蛋白通过抑制I型IFN的产生促进了VSV的复制。
3讨论与结论
口蹄疫是一种急性、高度接触性人畜共患传染病,我国主要以0型口蹄疫为主,A型口蹄疫呈点状散发,加之境外疫情传人的影响,给我国的畜牧业带来严峻的挑战。FMDV 3D蛋白作为RNA病毒依赖的RNA聚合酶,催化病毒RNA合成,在不同血清型中高度保守,且有研究表明3D在病毒复制早期产生,感染后1.5h首先检测到3D瞬时表达,在核仁中均匀分布。本试验成功构建了真核表达质粒pXJ41-Myc-3D,并在HEK-293T细胞中进行了表达验证。通过IFA检测到3D蛋白在PK-15胞核和细胞胞浆中都有表达,且胞核内表达明显,显示3D主要定位于细胞核,与毕研丽等的研究结果一致。
天然免疫又称先天性免疫或固有免疫,是机体对抗病毒感染的第一道防线,当病毒感染机体后模式识别受体(pattern recognition receptors,PRRs)识别病原相关模式分子(pathogen associat-ed molecule patterns,PAMPs),激活下游信号通路,诱导产生I型IFN等细胞因子,进而启动免疫应答。PRRs家族包括Toll样受体(TLRs)、RIG-I样受体(RLRs)、Nod样受体(NLRs).C型凝集素受体(CLRs),其中TLRs和RLRs主要识别病毒RNA。FMDV在长期进化中形成了一系列策略逃逸宿主的天然免疫,从而促进自身复制,维持对宿主的持续感染。已有研究表明,FMDV编码的多种蛋白可参与调控先天性免疫信号通路,例如Lb蛋白可以通过去除ISG15的泛素化阻断天然免疫的启动,裂解LGP2抑制IFN的表达。3C蛋白通过溶酶体路径降解蛋白激酶PKR,也能通过抑制NF-KB信号通路促进病毒复制。3A蛋白与RIG-I、MDA5和MAVS互作并抑制蛋白表达,抑制RLR介导的IFN产生。2B通过降解RIG-I和MDA5调节IFN信号通路。2B和2C蛋白协同作用,影响MHC-I复合物的形成,逃避机体的天然免疫。3B与RIG-I互作,抑制RIG-I介导的信号通路。本实验室前期发现FMDV3C通过阻断STATl/STAT2核易位来拮抗干扰素信号通路。FMDV L通过与sRNaseL的N末端结构域相互作用,抑制细胞凋亡,拮抗OAS/RNase L通路,抑制抗病毒天然免疫。有研究表明FMDV非结构蛋白3D作为一种反式作用元件调控TLR3信号通路的激活,促进TLR3通路介导的I型干扰素的产生。但天然免疫反应交错复杂,不同的PAMPs可能激活多条信号通路,通过不同分子間相互作用共同完成天然免疫应答。本试验选用HEK-293T作为研究细胞,由于该模式细胞中不含TLR3受体,可排除TLR3信号通路的影响,探究3D蛋白对I型IFN信号通路的作用。
水疱性口炎病毒是一种负链RNA病毒,属弹状病毒科水疱病毒属,是研究I型IFN信号通路常用的模式病毒,细胞感染VSV后可激活I型IFN信号通路。本研究采用VSV模拟病毒感染,通过双荧光素酶报告基因、Real-time
PCR及TCID50试验,发现3D蛋白能够通过抑制I型IFN信号通路的活化抑制先天性免疫,进而促进VSV的复制。
综上,FMDV 3D蛋白抑制了VSV诱导的IFN启动子的活性和IFN mRNA水平,促进了VSV的复制,提示FMDV 3D蛋白是病毒抵抗先天性免疫的重要因子,为其在I型IFN信号通路的作用及机制研究奠定了基础。
猜你喜欢
现代畜牧科技(2021年8期)2021-10-13
昆明医科大学学报(2021年5期)2021-07-22
生命科学研究(2018年1期)2018-05-29
食品科学(2018年10期)2018-05-23
法医学杂志(2015年4期)2016-01-06
法医学杂志(2015年4期)2016-01-06
西南医科大学学报(2015年1期)2015-08-22
中国当代医药(2015年9期)2015-03-01
西南军医(2015年6期)2015-01-23
当代畜禽养殖业(2014年6期)2014-02-27
